Топологические солитоны в двухцепочечной модели ДНК

Автор: Закирьянов Фарит Кабирович, Хамзин Салават Рифовим, Юлмухаметов Константин Раисович
Рубрика: Математика
Статья в выпуске: 10 (143), 2009 года.
Бесплатный доступ
Рассмотрена математическая модель нелинейной динамики ДНК. Получены солитонные решения, описывающие конформационные изменения в процессе функционирования ДНК. Проведено сравнение аналитических решений с результатами численного эксперимента.
Днк, нелинейная динамика, топологические солитоны
Короткий адрес: https://sciup.org/147158616
IDR: 147158616 | УДК: 577.3.01
Topological solitons in a DNA double strand model
A mathematical model of nonlinear DNA dynamics is presented. Solitonic solutions describing conformational transitions during DNA functioning are obtained. Analytical solutions are compared with numerical simulations results.
Текст научной статьи Топологические солитоны в двухцепочечной модели ДНК
Интенсивные исследования последних десятилетий показали, что в нелинейных системах различной физической природы возможны коллективные возбуждения в виде нелинейных уединенных волн - солитонов. Уединенные волны обладают рядом замечательных свойств, сближающих их с частицами - сохранение формы и скорости движения после взаимодействия. Они активно применяются для описания различных кооперативных эффектов в нелинейных динамических системах, в том числе молекулярных цепочках, например, таких как ДНК [1, 2]. Особый интерес представляют квазилинейные молекулярные цепочки, характеризуемые как геометрическим ангармонизмом, так и ангармонизмом потенциала междоузельного взаимодействия. Учет этих факторов приводит к моделям, описываемым системами нелинейных дифференциальных уравнений, решение которых крайне сложно получить аналитическими методами. Это приводит к необходимости разработки методов компьютерного эксперимента для исследования динамики подобного рода систем.
В работах [3, 4] изучена динамика топологических солитонов, описывающих образование открытого состояния двойной спирали ДНК. Показано, что в двойной спирали ДНК могут появляться четыре типа топологических солитонов (см. рис. 4 из [3]). Изучены взаимодействия между солитонами, их взаимодействие с неоднородностями цепи и устойчивость солитонов по отношению к тепловым колебаниям. Основным методом работ [3, 4] был вычислительный эксперимент с использованием дискретной модели ДНК, основанный на разработанной авторами вариационной технике для нахождения солитонов. Вместе с тем такие же решения можно получить и аналитически для модели цепочек в континуальном приближении.
В данной работе использовали дискретную модель ДНК [3, 4], основанную на модели Инг-лэндера [2]. На рис. 1 схематически представлена часть данной модели. Белые и черные прямоугольники соответствуют парам оснований АТ или GC, а линии - скелету двойной спирали. Гамильтониан для этой модели имеет следующий вид:
----2----+ e2sm -----2-----+ ^^Фп,1’Фп,2) > где Г(р„д, фп2 ) = kx (1 - cos фпХ) + кх (1 - cos ри2) +^ (1 - cos(^„ д - фп2)).
Здесь фпХ и фп2 - углы поворота азотистых оснований в п-х узлах первой и второй цепочки соответственно, 1Х и /2 - моменты инерции оснований первой и второй цепочки соответственно, ех и f2 - коэффициенты упругой связи, кх и к2 - жесткость взаимодействия соседних оснований в одной и разных цепочках соответственно.
В основе численного метода поиска солитонного решения лежит нахождение условного минимума для лагранжиана системы с соответствующими граничными условиями [3,4]:
^,1 = ^-оо,Ь ^,2 = Ф-®,2
ФN^ ~ф®,\' Фм,1 ~Ф®,2-
Найденные точки минимума служат начальными точками для решения системы уравнений движения. Решалась система дифференциальных уравнений второго порядка (задача Коши)
d^nj
_
BH^ nX n^
^ФпЯ - дН(фП,\ФП^ dt1 дфп^
(и = 0, ±1, ± 2...).
Интегрирование уравнений движения проводилось с помощью неявного метода Розенброка 2-го порядка точности.
Кроме численного анализа дискретной модели нами был проведен и теоретический анализ возможности получения решений с использованием теории возмущений. В континуальном приближении с учетом дисперсии длинноволновых колебаний динамика модели описывается системой двух нелинейных дифференци-
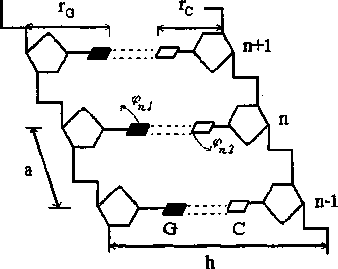
Рис. 1. Фрагмент двойной спирали ДНК, состоящей из трех пар оснований вида GC или (АТ). Расстояние между основаниями вдоль цепи а = 3.4 А, расстояние между цепями h = 16.15 А [3,4]
альных уравнений в частных производных:
1\Ф\тт ~Ч
а4 а4 2 2
^^ж-у^ + й Ф\уу
+ кх sin^ + к2 sin(^ -ф^ = 0,
^гФгтт "ег
( 4 4
a и г 2 ^2^ "у ^2>у + « Ф1уу
+ X] sin фг -к2 sin((9] - ф^) = 0 .
Коэффициенты перед членом с высшей дисперсией ^i^ и членом, содержащим нелинейность по пространственным производным, Ф^уФгуу при малых скоростях являются малыми. С увеличением скорости характерный размер кинков, определяемых уравнением sin-Гордон, уменьшается. В этом случае роль указанных слагаемых будет возрастать и особенно сильно проявляться именно вблизи предельной скорости солитона.
Для упрощения теоретических выкладок нами рассмотрена модель однородной молекулы ДНК, состоящей из двух одинаковых цепочек, где для простоты полагается 1Х = /2 = I, ех=ег=е. Система классических уравнений движения после перехода к новым временным и пространственным переменным путем замены т = t^Ijkx и у = x^sa/kx принимает вид:
Ф\« -Ф\хх-^-Ф\хххх *v^xx + sin Фх ^а^<фх - %) = 0,
125 2S
Фти-Ф1ХХ^ФтхФък + sin^2 -asin(^-ф^ = О, YLe 2s где безразмерный параметр a = k2/kx.
Покажем теперь, что рассматриваемая система уравнений допускает аналитические решения, подобные полученным в работе [3] в ходе численного эксперимента.
Решения будем искать в виде ряда
Ф\ = Фо + Ф\ + ^2 + •••» Фг = Фо + h + Ч> + -, где (Эо - решение системы, не учитывающей взаимодействие цепочек. Без учета взаимодействия цепочек (при а = 0) рассматриваемая система приводит к известному уравнению sin-Гордон, решение которого в виде 2л- -кинка имеет вид
Фо = 4arctgexp(z), где z = (1— s2)-^2^, 5 - скорость солитона, ^ = x-st. Подставляя решение в виде ряда в рассматриваемую систему, получим в первом приближении уравнения для возмущений фх и vx:
Ф\ "Фх +^1008(90 --Г-Фохххх + ^Ф0хФ0хх=Ь’
Y2s 2s
Ч "Ч + Ч COS% ""Фохххх + ^^0хх = °. 12s 2s
Математика
где cos % = 1 - 2/ch2 z. Полученная система уравнений имеет следующее решение:
С, к, +? thz-ik „ 15г, thz
—=---!---- I-----------edkA--!-- chz 24s(l-s2)2^^^^)^^ 12^(1-52)2 chz’
+? th.z-ik „ 15r, thz у, = —1------------г— e dk +---------, chz 24^(1-?)2 _i(i + fc2)ch^ 12s(l — s2)2 ch z где С] и C2 определяются из условия разрешимости уравнений для возмущений второго порядка малости (в нашем случае Q = О и С2 = 0). Эти константы определяют сдвиг кинков друг относительно друга. В нашем приближении этот сдвиг не удается получить аналитически. Таким образом, взаимодействие цепочек изменяет только форму кинка.
Общий вид полученного решения представлен на рис. 2 и качественно соответствует случаю (е) на рис. 4 [3]. Заметим, что при приближении скорости солитона к предельной ( з —> 1) теория возмущений дает большие погрешности.
Солитонное решение задачи можно охарактеризовать топологическим зарядом q = (^ь д2), где q, = (^оо,, " Ф®,, )/2 л- О' = 1,2) является целым числом (^,=0, ±1, ±2,...). Для нахождения солитонного решения с топологическим зарядом q нужно решить задачу на минимум с граничными условиями:
Ф-® д = ^-00,2 = 0’ ^00,1 = 2л^, <рт г = 2лq2-
При компьютерном моделировании стационарных солитонов с топологическим зарядом q = (1, 1) получены результаты, представленные на рис. 3.
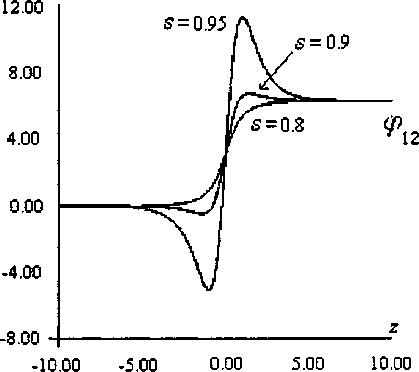
Рис. 2. Вид стационарных солитонов с топологическим зарядом q = (1,1) при различных значениях безразмерной скорости солитона
[-..$i-..$U
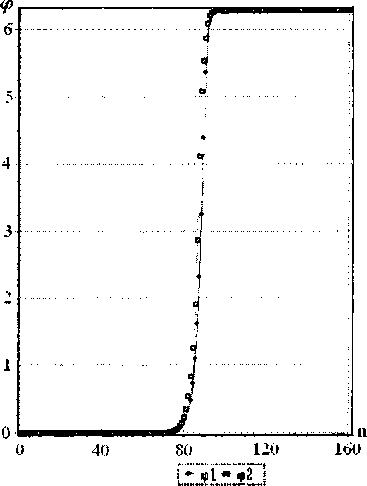
Рис. 4. Вид стационарного солитона с топологическим зарядом q = (1,1) при скорости s = 0,804. К = 0,234 Н/м. Ширина 25 звеньев в каждой цепочке. Тип оснований GC
Рис. 3. Вид стационарного солитона с топологическим зарядом q = (1,1) при скорости s = 0,592. К = 0,234 Н/м. Ширина 27 звеньев в каждой цепочке. Тип оснований GC
Закирьянов Ф.К., Хамзин С.Р., Топологические солитоны Юлмухаметов К.Р. в двухцепочечной модели ДНК
Видно, что происходит сдвиг кинков между двумя цепочками, то есть в одной из цепи кинк уходит немного вперед относительно другого кинка. Анализ показывает, что такой сдвиг происходит не при всех параметрах цепочки и скоростях уединенной волны, в частности, можно задать параметры таким образом и выбрать скорость солитона так, что сдвиг будет отсутствовать на всем протяжении движения солитона по цепи (рис. 4). Данный сдвиг объясняется неоднородностью цепи - у разных пар азотистых оснований различные параметры взаимодействия, масса и инерция, которые влияют на повороты углов в цепи. В случае теоретического исследования сдвиг отсутствует, так как рассматривалась только однородная цепочка (рис. 2). Представленная на рис. 4 динамика кинков без сдвига, имеет хорошее соответствие с аналитическими результатами.
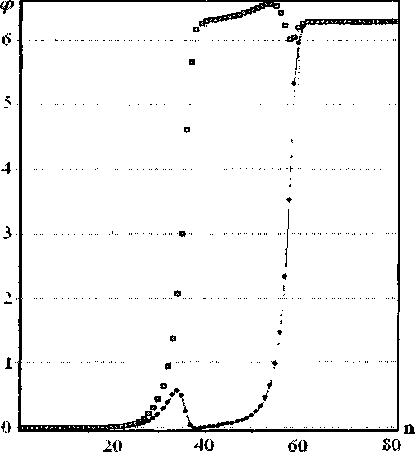
Зависимость энергии топологического солитона q = (1, 1) от его скорости, полученная аналитически, представлена на рис. 5 (.Ei). В ходе численного эксперимента была получена зависимость безразмерной энергии топологического солитона (Е) от безразмерной скорости (s) для всех трех интересующих нас типов топологических зарядов q = (1, 1) , q = (1,0) и q = (О, 1), которая представлена на рис. 6.
800 "1 кДж/'моль
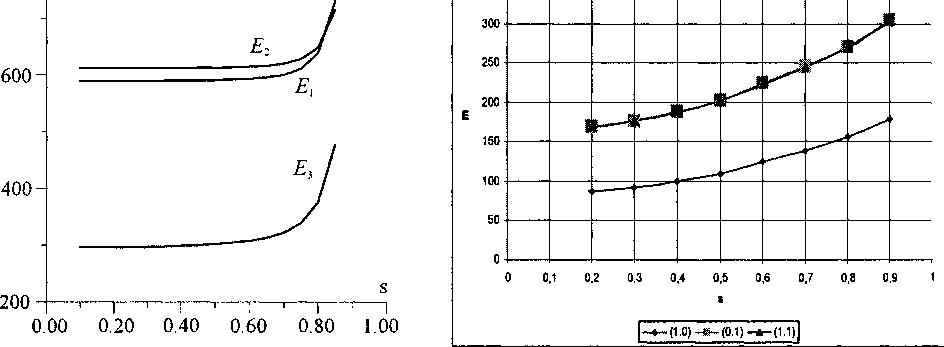
Рис. 5. Зависимости энергий стационарных солитонов с топологическими зарядами q = (1, 1) - Ei, q = (1, -1) - Ег и q = (1, 0) - Е3 от безразмерной скорости s (*i/e = 0,1,о = 0,1)
Рис. 6. Графики зависимости безразмерной энергии топологического солитона (А) от безразмерной скорости (s), для оснований GC
Видно, что графики зависимости безразмерной энергии от безразмерной скорости для топологических солитонов q = (1, 0) и q = (0, 1) практически совпадают, так же как и при аналитическом решении.
Для получения аналитических решений, описывающих солитоны с топологическим зарядом q = (1, -1), используем разложение
Ф\ = Фо + V\ + ^2 + Ф1 = "Фо + И + ^2 + -, при этом в первом приближении получим для возмущений следующую систему уравнений: ^1 -y"*^ cospo --^-Фохххх ^^ФохФохх + «sin(2^0) = О , 12г 2е
Ч - ч"+ч cos % —^-фОхххх ^^ФохФохх -asin(2%) = 0. 12г 2г
Её решение в виде стационарных волн имеет вид
^1 =
15К] 12s(l-s2)2
-2а --+ а- chz
^1
th z-ik
4 =
15^ 12s(l-s2)2
+ 2a--a + Jchz .
24=(l-s2)2}2(1+Ji2)ch^
кх thz-ik
eifadk,
24=(l-s2)2J2(H-li2)ch^
eitedk.
Математика
В итоге фх и р2 имеет солитонные решения с топологическим зарядом q = (1, -1), представленные на рис. 7 и качественно соответствующие случаю (ж) на рис. 4 [3]. Зависимость энергии топологического солитона q = (1, -1) от скорости солитона представлена на рис. 5 (£2)-
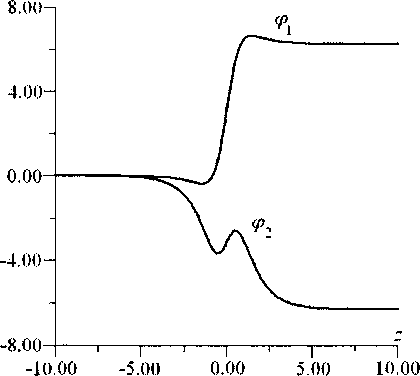
Рис. 7. Вид стационарного солитона с топологическим зарядом q = (1,-1) для значения безразмерной скорости s = 0,9
Решения, полученные численно, и представленные на рис. 8, имеют качественное сходство с аналитическими решениями (рис. 7). Отличие их связано с упомянутым выше влиянием неоднородности цепочек, приводящим к расхождению кинков с течением времени.
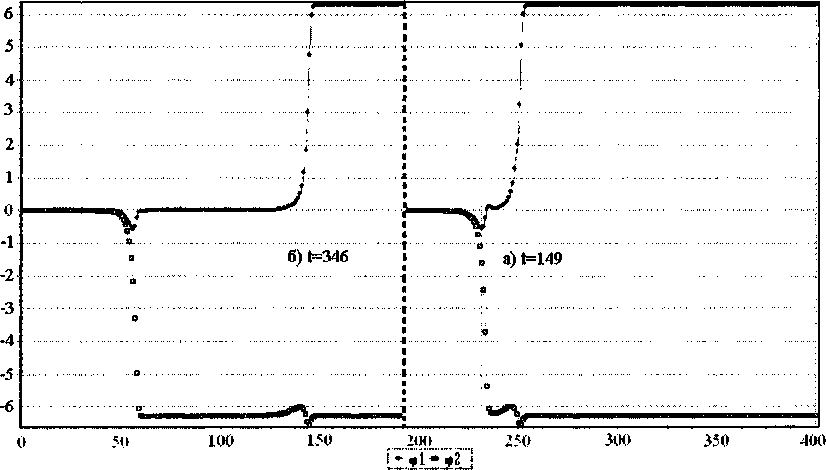
Рис. 8. Вид стационарного солитона с топологическим зарядом q = (1,-1), в разное время численного эксперимента (t- время в программе). Скорости s = 0,898. К = 0,234 Н/м. Ширина 32 звена в каждой цепочке. Тип основания GC. Солитон движется справа налево
Солитоны с топологическими зарядами q = (1, 0) и q = (0, 1) будем считать практически эквивалентными, поскольку они соответствуют одной физической ситуации распространения кинка по одной цепочке, сопровождаемого малым возмущением по другой цепочке. Отличие будет только в значениях энергий рассматриваемых солитонов. Решение рассматриваемой модели в данном случае ищется в виде разложения:
Ф\ = 9»*V\^Vi*-^ Ф2=ух+у2+.„,
В первом приближении для возмущений \ух и vx имеем систему уравнений:
Ух -Ух+Ух cosp0 -5-<р0хххх +^^Олх + asin(p0) = 0, 12s 2s
-
V, -V] + vT --7-фОхххх ^Mta -asm(%) = 0 . 12г 2s
Полученная система уравнений имеет следующее решение: Зк, thz fl thz-ik „
Vx= ---Ц-т----а +--------- r—edk
1 2s<\-sb2^z (2 24г(1-г2)2)_^(1 + ^.2)сЬ^
^1 =
Зкх thz f к\ 6£(1-S2)2chz (65(1-S2)2
-2a (shzln(2chz)-zchz).
Таким образом, взаимодействие цепочек изменяет только форму кинка, вид которого представлен на рис. 9, качественно соответствующем случаям (а-г) на рис. 4 [3]. Численное моделирование (рис. 10) указывает на хорошее качественное соответствие с полученными аналитически результатами. Зависимости энергии топологического солитона q = (1, 0) от его скорости, полученные аналитически и численно представлены на рис. 5 (Дз) и рис. 6.
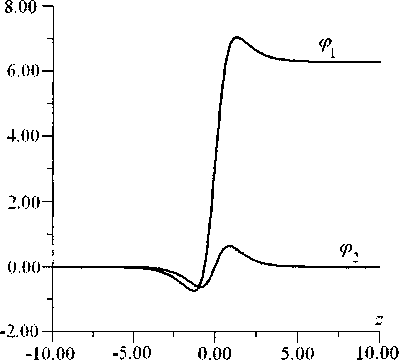
Рис. 9. Вид стационарного солитона с топологическим зарядом q = (1,0) для значения безразмерной скорости s = 0,9
Таким образом, теоретический анализ рассматриваемой модели однородной ДНК показал, что в системе могут быть решения в виде солитонов со следующими топологическими зарядами:
-
1) q = (1, 0) (или q = (0, 1)) (по одной цепочке распространяется возмущенный кинк, а по второй
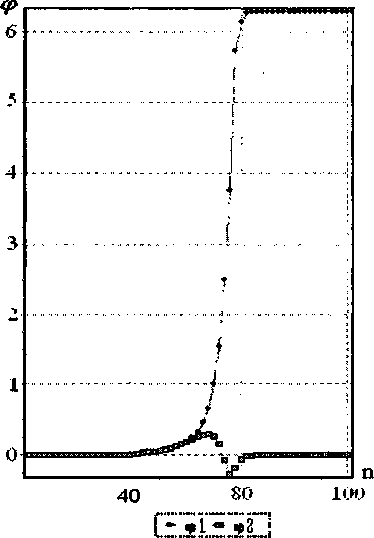
Рис. 10. Вид стационарного солитона с топологическим зарядом q = (1,0) при скорости 5 = 0,901. К = 0,8714 Н/м. Ширина 19 звеньев цепи. Тип основания GC
цепочке - малое возмущение), соответствующие локальным движениям, возникающим при транскрипции ДНК;
-
2) q = (1, 1) (по двум цепочкам распространяются в одном направлении и с одинаковыми скоростями два одинаковых возмущенных кинка), который соответствует переходу ДНК из одного конформационного состояния в другое;
-
3) q = (1, -1) (по двум цепочкам распространяются кинк и антикинк), который описывает процесс репликации ДНК.
Получены зависимости энергии топологических солитонов от безразмерной скорости s. При этом энергетически наиболее выгоден солитон с топологическим зарядом q = (1, 0) (рис. 5, 6). Проведено сравнение результатов численного и аналитического решений. Результаты показывают, что решения, полученные аналитическим методом и численным экспериментом, имеют качественное сходство.
Математика
Авторы выражают искреннюю благодарность профессору М.А. Шамсутдинову за полезные обсуждения в ходе выполнения работы.
Список литературы Топологические солитоны в двухцепочечной модели ДНК
- Yakushevich, L.A. Nonlinear Physics of DNA/L.A. Yakushevich. -Wiley, Chichester, 2004.
- Englander, S.W./S.W. Englander et al. Il Proc. Natl. Acad. Sei. U.S.A. -1980. -V. 77. -P. 7222-7226.
- Ковалева, H.A./H.A. Ковалева, A.B. Савин, Л.И. Маневич и др.//ВМС. -2006. -Т. 48, №3.-С. 454-471.
- Yakushevich, L.V./L.V. Yakushevich, A.V. Savin, L.I. Manevitch//Phys. Rev. E. -2002. -V. 66.-P. 016614.
- Якушевич, Л.В. Методы теоретической физики в исследовании свойств биополимеров/Л.В. Якушевич. -Пущино: ОНТИ НЦБИ АН СССР, 1990.
