Топологические солитоны в однородной асимметричной модели молекулы ДНК

Автор: Закирьянов Фарит Кабирович, Хамзин Салават Рифович
Рубрика: Физика
Статья в выпуске: 30 (206), 2010 года.
Бесплатный доступ
Рассмотрена математическая модель нелинейной динамики однородной, но асимметричной по основаниям искусственной молекулы ДНК. Получены солитонные решения, описывающие конформационные изменения в процессе функционирования ДНК. Исследовано влияние диссипации на динамику солитонов. Рассмотрено прохождение солитонов через границы однородных областей в искусственных последовательностях ДНК, приводящие к изменению топологии солитонов.
Днк, нелинейная динамика, топологические солитоны
Короткий адрес: https://sciup.org/147158651
IDR: 147158651
Topological solitons in the homogeneous asymmetric model of a DNA molecule
The mathematical model of the nonlinear dynamics of homogeneous asymmetric model of a DNA artificial molecule is considered. Solitonic solutions describing conformation changes in processes of functioning of DNA are obtained. Influence of dissipation on soliton dynamics is researched. It is shown that soliton propagation through the borders of homogeneous areas in artificial sequences of DNA leads to changes in soliton topology.
Текст научной статьи Топологические солитоны в однородной асимметричной модели молекулы ДНК
Развитие современной нелинейной физики привело к открытию новых элементарных механизмов, определяющих на молекулярном уровне протекание многих физических процессов в кристаллах, полимерных цепях, молекулах ДНК и других упорядоченных молекулярных системах. Развитие теории и практики вычислительного эксперимента позволяет исследовать динамику молекулярных систем, состоящих из большого числа частиц с большим выбором параметров и разнообразных условий, имитирующих реальные свойства систем. Большой интерес в связи с этим вызывает моделирование динамики ДНК с учетом ее гетерогенности и асимметрии по осно ваниям, что недоступно на нынешнем уровне развития науки аналитическим методам.
В работе [1] изучена динамика солитонов в молекуле ДНК, подробно описана численная схема метода исследования движения солитонов вдоль цепи двойной спирали ДНК. В этой работе для упрощения вычислений авторы сосредоточились только на вращательных движениях азотистых оснований вокруг сахаро-фосфатных цепей в плоскости, перпендикулярной главной оси двойной цепочки. Были найдены решения в виде уединенных волн (солитонов), описывающих открытое состояние пар оснований в ДНК; полученные решения классифицированы. Наряду с этим рассматривались устойчивость солитонов к большим тепловым колебаниям, взаимодействие между солитонами и взаимодействие солитонов с неоднородностями цепи. Для решения этих задач авторы использовали метод, эффективность которого была проверена в работах [2-4], где он использовался для анализа нелинейной динамики молекулярных цепей и полимерных кристаллов. Следует отметить, что, как было показано в цитируемых работах, солитоны могут дви гаться на всем протяжении макромолекулы с постоянной скоростью, меньшей скорости звука в рассматриваемой среде.
Целью данной работы является исследование влияния гетерогенности модели ДНК на условия зарождения и прохождения нелинейных уединенных волн.
Рис. 1. Фрагмент двойной спирали ДНК, состоящей из трех пар оснований вида GC или (АТ). Расстояние между основаниями вдоль цепи а = 3,4 А, расстояние между цепями h = 16,15 А [3,4]
В работе рассматривается дискретная модель В-формы молекулы ДНК, на рис. 1 схематически представлена часть данной модели. Белые и черные прямоугольники соответствуют парам оснований АТ или GC, а линии - скелету двойной спирали.
Плоская модель ДНК представляет две параллельные цепочки, расстояние между которыми равно h. Для удобства описания модели левой цепи будем приписывать индекс 1, а правой 2. В этой модели основания совершают вращение вокруг сахаро-фосфатного остова и всегда остаются перпендикулярными ему и соответственно этому: ф„,1 - угловое перемещение и-го основания первой цепочки, ^„ з - угловое перемещение и-го основания второй цепочки. Гамильтониан для двойной цепи имеет вид
+ Ел2$т^-_¥........^
И
^аР^Фп^пЯ
Опишем слагаемые, входящие в гамильтониан (1): первое и второе слагаемые описывают кинетическую энергию и-х пар основания (Z„j - момент инерции w-го основания первой цепи; l^ - момент инерции я-го основания второй цепи, точкой обозначено дифференцирование по времени 0- За взаимодействие соседних пар оснований вдоль цепочек отвечают третий и четвертый член в гамильтониане (1). Здесь параметр s„ , характеризует энергию взаимодействия и-го основания с (п + 1)-м основанием /-й цепи (/ = 1, 2). Параметр, характеризующий энергию взаимодей-. ствия, предположительно должен быть больше 60 кДж/моль в связи с тем, что угловое вращении оснований сопровождается преодолением барьера стекинг-взаимодействия и деформацией двугранных и валентных углов. Пятое слагаемое гамильтониана (1) описывает энергию взаимодействия между связанными парами оснований цепочек (всего имеем четыре типа пар оснований ар = АТ, ТА, CG, CG). В качестве потенциала взаимодействия возьмем потенциал вида
юарг
®ар\ ,
а ^'р
где Коф - константа потенциала взаимодействия, coap\, o)api - частоты вращательных колебаний оснований при движении в одном и в противоположных направлениях (22 и 81,5 см"1 [1,4]) соответственно.
В основе численного метода поиска солитонного решения лежит нахождение условного минимума для лагранжиана системы с соответствующими граничными условиями [1,5]:
^1,1 =^-оо,1’ ^,2 =^-оо,2’
ФмД -^00,1» Фмд
Найденные точки минимума служат начальными точками для решения системы уравнений движения:
d^nX _ дН^фпДф„ ^
пЛ dt2 5<рпХ ’
’ (« = 0, ±1, ±2 ...)
т ^фп2 _ ЭН^фпДфп2)
dr дфп2
Модель учитывает также диссипацию в системе вследствие взаимодействия молекулы ДНК с водным окружением. Интегрирование уравнений движения проводилось с помощью неявного метода Розенброка 2-го порядка точности.
Известно, что для нелинейных волновых уравнений решения в виде бегущих волн требуют начальных условий специального вида, а именно, в виде самих этих решений в начальный момент времени. Для реальных систем не менее важно знать, что происходит при достаточно произвольных решениях. Очевидно, что это может быть исследовано только численно. Рассмотрим динамику уединенных волн в случае, когда начальное импульсное возбуждение задается в произвольном месте двойной цепочки. Первое импульсное возбуждение возьмем в виде где/- параметр, который задает точку приложения начального возбуждения (зависит от п).
В таком виде данное возбуждение быстро затухает со временем, но если амплитуду возмущения увеличить в два раза, то при определенных значениях параметров (в т.ч. начальной скорости уединенной волны) данное возбуждение становится устойчивым 2я"-кинком. Другими словами, это «колоколообразное» возмущение переходит в «кинкоподобное» возбуждение Фп1 = [l + th//(n-A,'/2)]^z (рис. 2). В случае, когда амплитуда возмущения велика (множитель mmji (т > 2), возбуждение со временем всегда становится стационарным и принимает вид 2л-кинка.
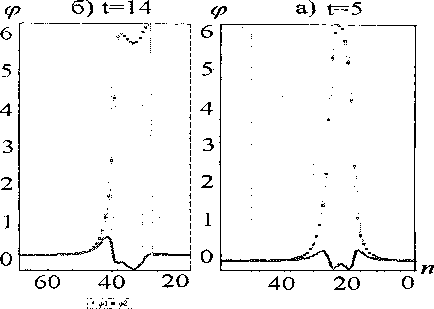
Рис. 2. Динамика возбуждения и прохождения по цепи уединенной волны топологии q=(0,1). Солитон движется справа налево. Здесь и далее значения углов поворота представлены в радианной мере
В качестве начального возбуждения нами были рассмотрены еще два колоколообразных профиля для уединенных волн:
Фм = 7---- qi ’ ф^ = 7—;------- qf i-V2.
(l + ^'(nV)) (1ф(и-/)) )
Расчеты показали, что со временем оба эти возбуждения затухают, то есть в данной модели двойной спирали ДНК не могут существовать волны с таким профилем. С увеличением амплитуды этих начальных возмущений они ведут себя аналогично рассмотренному выше. Таким образом, в рамках данной модели уединенные волны можно возбуждать не только задавая «кинко-вые» начальные условия, но и начальные условия в виде «колоколообразных» импульсов.
Солитонное решение задачи можно охарактеризовать топологическим зарядом q = (^, q^, где q, = (^ , - ф-^,^2л(р = 1,2) является целым числом (^ = 0, ±1, ±2, ...). Для нахождения солитонного решения с топологическим зарядом q нужно решить задачу на минимум лагранжиана с граничными условиями:
^=0,1 = ^-»,2 = °, ^00,1 = 2^1, ф^ = 2л q2.
В ходе численного исследования удалось установить, что в данной модели молекулы ДНК могут существовать семь типов топологических солитонов # = (1, 0), (0, 1), (-1, 0), (0, -1), (1, 1),
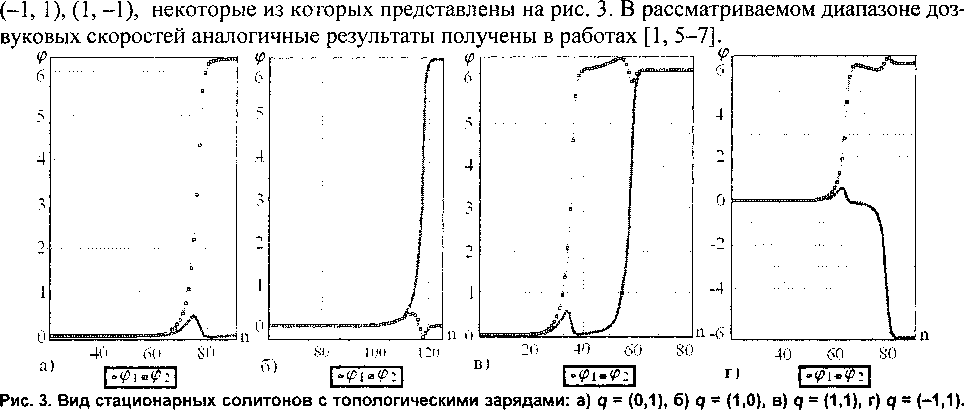
Сдвиг солитонов обусловлен разницей в энергиях взаимодействия оснований в комплементарных цепочках
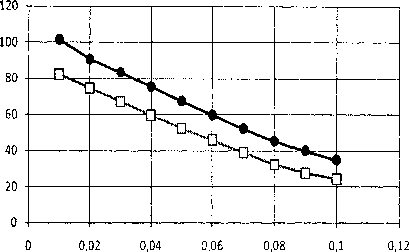
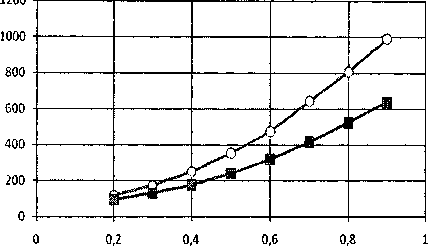
Далее нами была исследована зависимость энергии солитонов от коэффициента диссипации (рис. 4, 6) и скорости солитонов (рис. 5, 7). В качестве последовательности пар азотистых осно- ваний в модели ДНК, была взята искусственная последовательность типа AT-GC-TA-CG-AT-GC-TA-CG и т.д.
■♦■(0,1) 43-(1,0)
Рис. 4. Зависимости безразмерной энергии топологических солитонов q = (0, 1), q = (1, 0) от коэффициента диссипации
*О*|1,0) «^(0,1)
Рис. 5. Зависимости безразмерной энергии топологических солитонов q =(1, 0) и q = (0, 1) от безразмерной скорости (s)
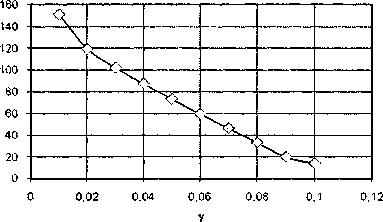
Рис. 6. Зависимость безразмерной энергии топологического солитона (1, 1) от коэффициента диссипации
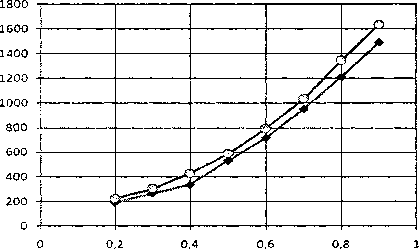
(1,1) "'О— (1,-1)
Рис. 7. Зависимости безразмерной энергии топологических солитонов (1, 1) и (1, -1) от безразмерной скорости (s)
Также в качестве реальной последовательности оснований в молекуле ДНК был рассмотрен геном Т7А\ в количестве 169 оснований. Последовательность оснований Т7А\ представлена ниже.
TTGTCTTTA ТТАА ТА САА СТС А СТА ТА A GGA GA GA САА СТТАА AGAGACTsoTAAAAGATTAATTTAAAATTTATCAAAAAGAGTATTG А СТТА А А СТСТААюоС СТ A TAGGATACTTACAGCCATCGAGAGGG ACACGGCGAATAGCCATCCisqCAATCGACACCGGGGTCAA
На данной последовательности оснований были получены следующие результаты (рис. 810).
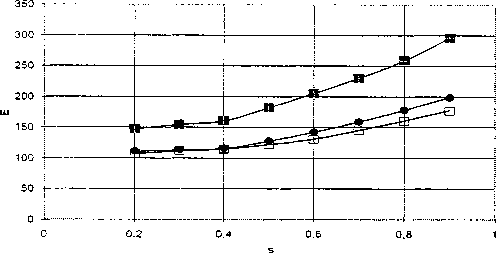
I -»-<1.0) -e-(O.D —я—<1.1)1
Рис. 8. Зависимости безразмерной энергии солитонов от безразмерной скорости (s) для 77А1
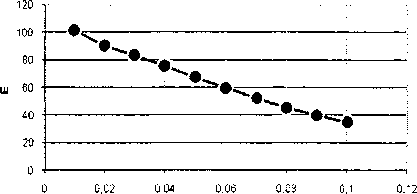
Рис. 9. Зависимость безразмерной энергии топологического солитона (0, 1) от коэффициента диссипации для 77А)
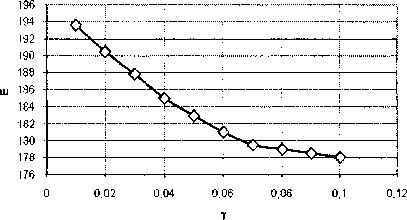
Рис. 10. Зависимость безразмерной энергии топологического солитона (1, -1) от коэффициента диссипации для 77Ai
Как и можно ожидать, в нашей модели энергия топологического солитона экспоненциально уменьшается с увеличением коэффициента вязкости. При больших значениях коэффициента диссипации (больше 0,1) происходит потеря энергии солитона со временем, пока она не достигнет близких к нулю значений.
Далее нами было рассмотрено прохождение солитонов через границу, разделяющую две однородные области. Мы составили модель молекулы ДНК таким образом, что половина цепи состоит из пар АТ-оснований, а вторая половина - из GC-оснований. В ходе численного эксперимента было установлено, что при переходе солитона из одной части (АТ) цепи в другую её часть (GC) изменяется энергия солитона. Например, при скорости солитона 5 = 500 (здесь и далее параметры заданы в безразмерных единицах), параметре кооперативности [1] g = 30, топологии q = = (1, 0), при переходе AT^GC энергия изменилась: 89,42(АТ) - 76,03(GC) = 13,39; для топологии (1, -1) при тех же параметрах: 172,79(АТ) - 147,51(GC) = 25,28. Отсюда видно, что при переходе солитона AT->GC его энергия уменьшается, а при переходе GC->AT энергия увеличивается. Объясняется это тем, что между парами GC имеется три водородные связи, а между парами АТ -две.
В ходе численного эксперимента удалось также установить минимальную скорость прохождения солитона по цепочке, так чтобы в той части цепи, где в качестве пар азотистых оснований были выбраны пары GC, волновое возбуждение не затухало, а в части цепи с парами АТ - волна затухала. Для топологии ^ = (1,0) диапазон скоростей, при которых происходит затухание уединенной волны, равен от s = 300 до s = 1980 при следующих параметрах: g = 21, К = 0,234, у = 0, и = 800 (переход AT->GC) (рис. 11); для топологии q = (0, 1) при скоростях 5 = 300 при следующих параметрах: g = 18,36, К = 0.234, у= 0, и = 800 (переход GC-»AT) (рис. 12).

Рис. 12. Прохождение топологического солитона (0,1) через границу области между двумя различными парами азотистых оснований. Граница раздела находится на 100-м узле цепи
Рис. 11. Прохождение топологического солитона (1,0) через границу области между двумя различными парами азотистых оснований. Граница раздела находиться на 100-м узле цепи
При исследовании солитонов топологии q = (1, 1) было получено следующее: при скоростях 5 = 300...500 и g = 20 в части с основаниями АТ кинки движутся вместе без сдвига (сдвиг показан на рис. 3,в), а в части с парами оснований GC топология q = (1, 1) переходит в топологию q = (0, 1), то есть в одной из цепочек (с основаниями G) уединенная волна затухает. Энергия солитона в части с основаниями АТ равна ^ij^at = 110,19, затем при переходе из части с основаниями АТ в часть с основаниями GC энергия увеличивается до E(i .цат^сс = 170,39, со временем, когда кинк в первой цепочке затухает и солитон приобретает топологию q = (0, 1), энергия равна ^(цц-ос = = 112,15 (при следующих параметрах: 5 = 300, g = 20). Если же мы рассмотрим солитон только с топологией q = (0,1) в части с основанием GC при тех же параметрах, то его энергия равна До ц. ос=77,42. ’
В случае с топологией q = (1, -1) при скоростях 5 = 300.. .500 и g = 21 переходе AT->GC кинк в первой цепочке затухает и в итоге возбуждение приобретает топологию q = (0, -1). При возбуждении уединенной волны в АТ со временем происходит отрыв кинка во второй цепочке от кинка в первой цепочке, энергия всей волны равна Е^ -щат = 143,47. На момент перехода кинком во второй цепочке границы между однородными областями энергия волны равна Д1-1)= 154,52, а когда границы достигает кинк в первой цепочке и затухает (прямо на границе раздела) энергия солитона становится равной Е^ -ц-ос = 115,22. В то же время, если рассмотрим энергию кинка q = = (0, -1), который движется только во второй цепочке в части с основаниями GC, при тех же параметрах его энергия будет равна До.-ц-ос = 79,04.
На рис. 13 и 14 представлены графики прохождения солитонов с топологиями ^ = (1, 1) и q = (1, -1) через границу однородных областей. Видно, что кинки в первых цепочках при прохождении 100-го узла (границы) затухают, а в комплементарных цепочках кинки проходят границу беспрепятственно. В итоге топология солитона меняется.
Рис. 13. Прохождение топологического солитона q = (1, 1) через границу области между двумя различными парами азотистых оснований. Граница раздела находится на 100-м узле цепи: а) кинк в первой цепочке, б) кинк во второй цепочке
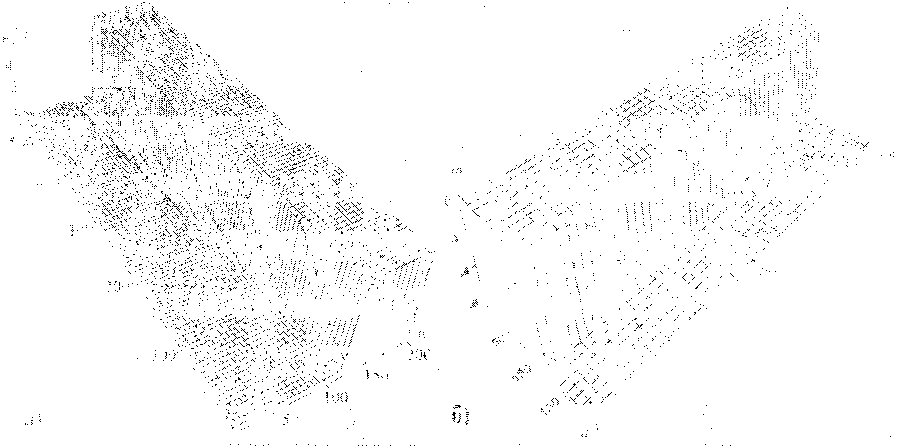
Рис. 14. Прохождение топологического солитона q = (1, -1) через границу области между двумя различными парами азотистых оснований. Граница раздела находится на 100-м узле цепи; а) кинк в первой цепочке, б) кинк во второй цепочке
Итак, в настоящей работе рассмотрена модель, учитывающая неоднородность распределения пар азотистых оснований (рассматривались различные типы чередования оснований в цепи), а также рассмотрена последовательность, соответствующая реальной последовательности в геноме бактериофага ПА ]. Рассмотрена динамика уединенных волн в случае, когда начальное импульсное возбуждение задается в любом месте двойной цепи. Показано, что солитонная динамика возможна и в случае колоколообраных начальных импульсов. Была исследована динамика топологических солитонов в зависимости от параметров цепи и скорости волны в макромолекуле. Исследована зависимость энергии топологических солитонов от безразмерной скорости s (рассматривался только дозвуковой диапазон скоростей) - с ростом скорости энергия солитона увеличивается. Полученные результаты качественно аналогичны результатам, полученным ранее на однородных цепочках, с соответствующими количественными отклонениями, объясняемыми различиями в энергиях взаимодействия разных пар оснований [6-8]. Для неоднородных двухцепочечных моделей рассмотрено влияние диссипации на динамику солитонов. Показано, что энергия солитона экспоненциально уменьшается с ростом коэффициента диссипации у. При достаточно больших значениях / происходит потеря энергии солитона до нуля. Была изучена динамика прохождения уединенной волны через границу между двумя различными парами оснований. Показано, что при этом может меняться топология солитонов.
Список литературы Топологические солитоны в однородной асимметричной модели молекулы ДНК
- Yakushevich, L.V. Nonlinear dynamics of topological solitons in DNA/L.V. Yakushevich, A.V. Savin, L.I. Manevitch//Physical Review. -2002. -E. 66.-P. 016614-1-016614-6.
- Twisted topological solitons and dislocations in polymer crystal/A.V. Savin, J.M. Khalack, P.L. Christiansen, A.V. Zolotaryuk//Physical Review В.-2002.-V. 65.-P. 054106-1-054106-10.
- Savin, A.V. Topological solitons in spiral polymeric macromolecules: A chain surrounded by immovable neighbors/A.V. Savin, L.I. Manevitch//Physical Review B. -2001. -V. 63. -P. 224303-1-224303-13
- Савин, А.В. Динамика топологических солитонов в системе слабосвязанных цепей/А.В. Савин, Е.А. Зубова, Л.И. Маневич//ВМС, серия А. -2005. -Т. 47, № 4. -С. 637-651.
- Топологические солитоны в неоднородной молекуле ДНК/Н.А. Ковалева, А.В. Савин, Л.И. Маневич и др.//ВМС, серия А. -2006. -Т. 48, № 3. -С. 454-471.
- Хамзин, С.Р. Численное исследование нелинейных уединенных волн в молекулярных цепочках методом молекулярной динамики/С.Р. Хамзин//Всероссийская школа-конференция для студентов, аспирантов и молодых ученых «Фундаментальная математика и ее приложения в естествознании»: сб. науч. тр. -Т. 3: Физика. -2008. -С. 300-303.
- Хамзин, С.Р. Численное исследование нелинейных уединенных волн в модели молекулы ДНК/С.Р. Хамзин//Всероссийская школа-конференция для студентов, аспирантов и молодых ученых «Фундаментальная математика и ее приложения в естествознании»: тезисы докладов, Физика.-2007.-С.42.
- Закирьянов, Ф.К. Топологические солитоны в двухцепочечной модели ДНК/Ф.К. Закирьянов, С.Р. Хамзин, К.Р. Юлмухаметов//Вестник ЮУрГУ. Серия «Математика, физика, химия». -2009. -Вып. 12. -№ 10 (143).-С. 8-14.
