Уникальное захоронение воина гунно-сарматского времени в Западно-Сибирской лесостепи: результаты палеогенетического анализа

Автор: Пилипенко А.С., Черданцев С.В., Трапезов Р.О., Чикишева Т.А., Поздняков Д.В., Молодин В.И.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Антропология и палеогенетика
Статья в выпуске: 4 т.46, 2018 года.
Бесплатный доступ
В статье представлены результаты палеогенетического исследования останков мужчины из погр. 688 памятника Сопка-2 в Барабинской лесостепи. Особенности материальной культуры и погребальной обрядности, а также монголоидный центрально-азиатский антропологический тип погребенного однозначно свидетельствуют о его инотерриториальном происхождении. Были исследованы маркеры с однородительским наследованием: митохондриальная ДНК (последовательность ГВС I и информативные позиции в кодирующей части мтДНК) и Y-хромосома (аллельный профиль 17 STR-локусов Y-хромосомы). Установлена принадлежность мтДНК исследуемого индивида к восточно-евразийской гаплогруппе D4b1, а Y-хромосомы - к восточно-евразийской гаплогруппе Q (Q1a). Таким образом, выявлен восточный вектор его глубоких генетических корней, что согласуется с палеоантропологическими данными. Результаты филогеографического анализа с использованием данных по древним и современным популяциям Евразии свидетельствуют о широком распространении выявленного нами варианта восточно-евразийской по происхождению гаплогруппы мтДНК в восточных и западных районах степного пояса. В частности, показано его присутствие в скифское время далеко за пределами Восточной Евразии -у классических скифов Северного Причерноморья. Таким образом, генетические данные не позволяют однозначно локализовать регион, из которого исследуемый индивид прибыл на территорию Барабинской лесостепи. Предложен сценарий, объясняющий противоречивость данных палеогенетики и палеоантропологии о восточных генетических корнях погребенного и преимущественно западный (сармато-аланский) круг аналогий для сопроводительного инвентаря из этого погребения. Обсуждаются возможности и ограничения использования палеогенетических методов для реконструкции различных аспектов происхождения представителей древнего населения
Митохондриальная днк, str-маркеры, y-хромосома, гунно-сарматское время, западная сибирь, степной пояс евразии, миграции
Короткий адрес: https://sciup.org/145145898
IDR: 145145898 | УДК: 575.17 | DOI: 10.17746/1563-0102.2018.46.4.123-131
Текст научной статьи Уникальное захоронение воина гунно-сарматского времени в Западно-Сибирской лесостепи: результаты палеогенетического анализа
С первой половины I тыс. до н.э. и в последующее скифское, гунно-сарматское время, а также в период раннего Средневековья кочевые племена Евразийского степного пояса играли важную роль в формировании культурного ландшафта и генетического состава населения Евразии. Один из пиков активности древних кочевников приходится на гунно-сарматскую эпоху (последние века до нашей эры – первая половина I тыс. н.э.). Масштабные перемещения популяций человека, которым сопутствовали культур- ные и генетические связи, охватили не только степь, но и обширные сопредельные территории. Не была исключением и лесостепная зона Западной Сибири, в т.ч. Барабинская лесостепь в междуречье Оби и Иртыша. Тем не менее археологических памятников, связанных с непосредственным присутствием степных кочевников гунно-сарматского времени, на юге Западно-Сибирской равнины мало. Одним из наиболее ярких таких комплексов является погребение воина в мог. 688 на разновременном погребально-ритуальном памятнике Сопка-2 в Венгеровском р-не Новосибирской обл. (рис. 1). Сопроводительный инвентарь
Рис. 1. Расположение памятника Сопка-2 в Барабинской лесостепи.
позволил датировать захоронение поздним периодом гунно-сарматского времени (IV–V вв. н.э.) [Молодин, Чикишева, 1990], что впоследствии было подтверждено радиоуглеродным анализом (подробнее см. далее). Богатство и хорошая сохранность прежде всего предметов вооружения указывают на отношение погребенного к военному делу.
Захоронение 688 могильника Сопка-2 с точки зрения материальной культуры и элементов погребальной обрядности, а также особенностей антропологического комплекса погребенного резко отличается от типичных для Барабинской лесостепи погребений гунно-сарматского времени [Молодин, Елагин, 1990], что однозначно свидетельствует об инотерриториальном происхождении индивида. Решение этого вопроса требует многостороннего анализа различных элементов рассматриваемого комплекса. Отдельные особенности погребального обряда и сопроводительного инвентаря имеют широкий круг аналогий в материалах памятников степного пояса Евразии, как его западных, так и восточных областей [Molodin, 1995]. Данные физической антропологии указывают
на принадлежность индивида к монголоидному центрально-азиатскому антропологическому типу и наиболее вероятное происхождение из южных районов Восточной Сибири [Молодин, Чикишева, 1990].
В данной работе представлены результаты молекулярно-генетического анализа о станков индивида из погр. 688 могильника Сопка-2 и их интерпретация в соответствии с полученными и опубликованными ранее археологическими [Там же; Molodin, 1995] и палеоантропологическими [Молодин, Чикишева, 1990] характеристиками элементов этого погребального комплекса.
Материалы и методы
Краткая археологическая и палеоантропологическая характеристика материалов погребения. Раскопки погр. 688, которое относится к комплексу разновременных памятников, объединенных общим названием Сопка-2 (Венгеровский р-н Новосибирской обл., место слияния рек Омь и Тартас) проведены в 1989 г. под руководством В.И. Молодина . В могиле находились останки взрослого мужчины 35–40 лет (все палеоантропологические определения выполнены Т.А. Чикишевой). Археологические и антропологические материалы из погребения хранятся в фондах Института археологии и этнографии СО РАН (г. Новосибирск). Подробная характеристика погребального комплекса опубликована ранее [Молодин, Чикишева, 1990; Molodin, 1995]. Здесь приведем лишь основные его особенности, имеющие значение для интерпретации генетических данных.
Погребение грунтовое (рис. 2). Могильная яма овальной формы ориентирована по линии ССВ – ЮЮЗ. В заполнении, в восточной части могилы, лежали кости лошади. Мужчина был погребен в вытянутом положении на спине, головой на ССВ. В могиле обнаружен богатый сопроводительный инвентарь (рис. 3–5). Почти все предметы, за исключением костяного двудырчатого псалия (см. рис. 4, 9 ), происходят с территории за пределами Западно-Сибирской лесостепи. Между погребенным и западной стенкой могилы обнаружен железный палаш (см. рис. 3, 1 ) со сложносоставным навершием (см. рис. 4, 1–4 ; 5), положенный в могилу в ножнах, от которых сохранилась массивная серебряная накладка и кусочки дерева со следами красной окраски (см. рис. 3, 2 ). Палаш крепился к поясу с помощью скобы из нефрита (см. рис. 4, 10 ). Элементы поясной гарнитуры (см. рис. 4, 5–7 ) имеют аналогии в материалах памятников западной части Евразийского степного пояса и позволяют надежно датировать рассматриваемый погребальный комплекс IV–V вв. н.э., т.е. поздним периодом гунно-сарматского времени. Это подтверждает радиоуглеродная дата, полученная
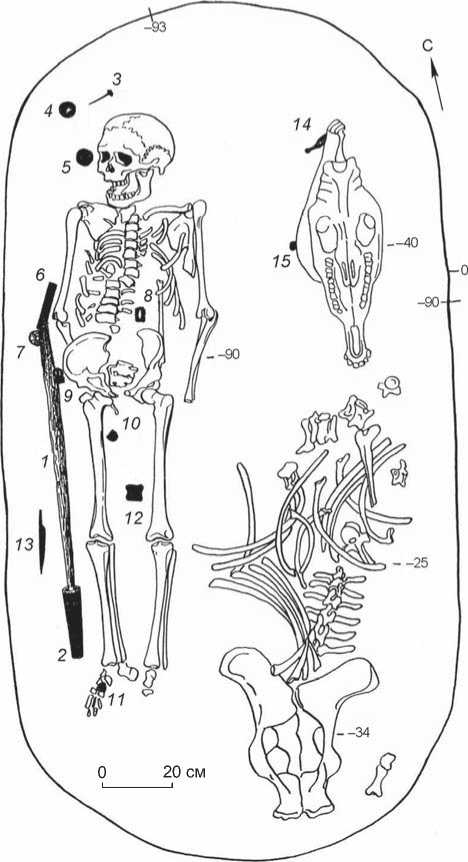
Рис. 2. План погр. 688 могильника Сопка-2.
1 – железный палаш; 2 – серебряный наконечник ножен; 3 – бронзовый гвоздь; 4 – нефритовый диск; 5 – золотое навершие; 6 – нефритовая скоба; 7 – навершие из горного хрусталя; 8 – железная обойма; 9 – серебряная пряжка; 10, 11 – серебряные наконечники ремня; 12 – лошадиная бабка; 13 – железный нож; 14 – костяной псалий; 15 – костяная застежка.
по зубу человека: ее калиброванные значения составляют 402–430 (1σ) и 393–474 (2σ) гг. н.э. (лабораторный индекс UGAMS23100/1214). В погребении обнаружены железный нож (см. рис. 4, 11 ), имеющий широкий круг аналогий, костяная застежка (см. рис. 4, 12 ), а также лошадиная бабка (см. рис. 4, 13 ), помещенная между ног погребенного.
Обобщая круг аналогий особенностям погр. 688 и найденным в нем предметам, можно отметить, что с гуннскими захоронениями Центральной Азии рассматриваемый комплекс связан лишь близкой к северной ориентацией могильной ямы и погребенного, а также

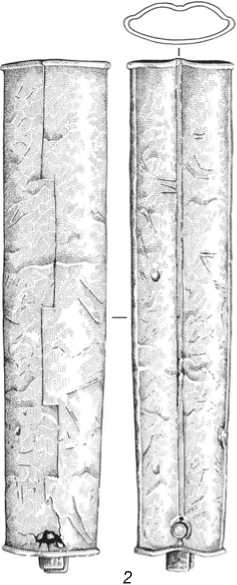
0 2 cм
Рис. 3. Железный палаш ( 1 ) и серебряный наконечник ножен ( 2 ).
Рис. 4. Сопроводительный инвентарь.
1–4 – детали навершия палаша; 5–7 – серебряная гарнитура пояса; 8 – железная обойма; 9 – костяной псалий; 10 – нефритовая скоба; 11 – железный нож;
12 – костяная застежка; 13 – лошадиная бабка.

присутствием нефритовой портупейной скобы. Основные параллели тяготеют к западу евразийских степей, к сармато-аланскому миру. Хотя в пределах ЗападноСибирской равнины аналог палаша, точнее его сложной рукояти, известен на равнинном Алтае, в Тугозво-новском захоронении [Уманский, 1978].

Рис. 5. Навершие палаша.
Морфологические особенности черепа мужчины из погр. 688 памятника Сопка-2 свидетельствуют о его принадлежности к монголоидному центрально-азиатскому антропологическому типу [Молодин, Чикишева, 1990], распространенному в более восточных районах Сибири и Центральной Азии. Этот тип не характерен для населения раннего железного века Западной Сибири и Алтая. Череп из погр. 688 находит аналогии в материалах гунно-сарматского могильника Кокэль, расположенного на территории Тувы. Краниологическое сходство в некоторых случаях доходит до тождества [Там же]. Таким образом, морфологические особенности погребенного свидетельствуют о его происхождении из расовой среды, отличной от основного населения ЗападноСибирской лесостепи гунно-сарматского времени, и указывают на возможную связь с населением во сточных районов Южной Сибири и Центральной Азии.
Палеоантропологический материал, использованный для молекулярно-генетического исследования. Костные останки погребенного характеризовались высокой степенью макроскопической сохранности, что косвенно указывало на потенциально хорошую сохранность ДНК. В качестве материала для палеоге-нетического исследования были использованы фрагменты левой бедренной кости.
Предварительная обработка палеоантропологического материала и экстракция ДНК. Молекулярно-генетические исследования выполнены в палеоге-нетической лаборатории межинститутского сектора молекулярной палеогенетики ИЦиГ СО РАН и ИАЭТ СО РАН (г. Новосибирск). Использовались методы, описанные в наших работах [Pilipenko et al., 2015, Pilipenko, Cherdantsev, Trapezov et al., 2018]. Поверхность кости обрабатывали 5%-м раствором гипохлорита натрия для разрушения возможных загрязнений современной ДНК, облучали ультрафиолетом. Поверхностный слой (~1–2 мм) удаляли механически. Из компактного костного вещества высверливали мелкодисперсный порошок, который инкубировали в 5М гуанидинизотиоционатном буфере при температуре 65 ºС и постоянном перемешивании. ДНК выделяли методом фенол-хлороформной экстракции с последующим осаждением изопропанолом.
Анализ генетических маркеров. В качестве основных информативных локусов в рамках данного исследования анализировали две системы молекулярно-генетических маркеров: митохондриальная ДНК (последовательность ГВС I и соответствующий информативный однонуклеотидный полиморфизм в кодирующей части мтДНК) и набор из 17 STR-локусов Y-хромосомы. В качестве вспомогательного локуса анализировали полиморфный фрагмент гена аме-логенина (маркер половой принадлежности останков) параллельно с набором из девяти аутосомных STR-локусов. Методы генотипирования каждой из перечисленных систем маркеров приведены ниже.
Амплификацию ГВС I мтДНК проводили двумя разными методами: четырех коротких перекрывающихся фрагментов посредством однораундовой ПЦР [Haak et al., 2005] и одного длинного фрагмента с помощью «вложенной» ПЦР (включала два раунда реакции) [Пилипенко и др., 2008]. Последовательности нуклеотидов определяли с использованием набора реактивов ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). Продукты секвенирующей реакции анализировали на автоматическом капиллярном секвенаторе ABI Prism 3130XL Genetic Analyser (Applied Biosistems, США) в центре коллективного пользования «Геномика» СО РАН . Амплификацию интересующих нас фрагментов кодирующей части мтДНК проводили с помощью олигонукле- отидных праймеров из работы [Wilde et al., 2014]. Филогенетическое положение исследуемого варианта мтДНК устанавливали на основании существующей классификации структурных вариантов мтДНК (www. [van Oven, Kayser, 2009].
Профили 17 STR-локусов Y-хромосомы определяли с помощью коммерческого набора реактивов AmpFlSTR® Y-filer® PCR Amplification Kit (Applied Biosystems, США) согласно инструкции производителя. Вероятную принадлежность исследованных STR-гаплотипов Y-хромосомы к гаплогруппам устанавливали с использованием двух программ-предикторов: Whit Athey`s Haplogroup predictor (http://www. и Vadim Yurasin`s YPredictor 1.5.0 .
Определение профилей девяти аутосомных STR-локусов и анализ полиморфизма участка гена амелогенина (маркер половой принадлежности останков) проводили с помощью коммерческого набора реактивов AmpFlSTR® Profiler® Plus PCR Amplification Kit (Applied Biosystems, США) согласно инструкции производителя.
Меры против контаминации и верификация результатов. Все работы с древним материалом выполнены в специально оборудованной для палеогенети-ческих исследований лаборатории межинститутского сектора молекулярной палеогенетики (ИЦиГ СО РАН, ИАЭТ СО РАН, г. Новосибирск). Меры против контаминации и процедуры верификации результатов описаны в нашей статье [Пилипенко, Трапезов, По-лосьмак, 2015]. Их реализация, а также особенности полученных результатов свидетельствуют о достоверности экспериментальных данных.
Результаты и обсуждение
Ранее нами было показано, что палеоантропологические материалы из археологических памятников Ба-рабинской лесостепи, демонстрирующие высокую степень макроскопической сохранности, содержат аутентичную ДНК, пригодную для проведения анализа как мтДНК, так и ядерных локусов [Пилипенко и др., 2017]. Результаты настоящего исследования подтверждают этот вывод. Используя три экстракта суммарной ДНК из материала бедренной кости индивида из погр. 688 памятника Сопка-2, мы получили воспроизводимые данные по структуре мтДНК и ядерных локусов (полиморфный участок гена аме-логенина, STR-локусы аутосом и Y-хромосомы).
Участок ГВС I контрольного района ДНК в позиции 15996–16403 содержал пять нуклеотидных замен относительно Кембриджской референсной последовательности мтДНК и характеризовался гаплотипом 16223T-16239T-16243C-16319A-16362C. Структура гаплотипа однозначно свидетельствует о принадлежности исследуемого образца мтДНК к восточно-евразийской гаплогруппе D. С этим согласуется и статус позиции 5178A в кодирующей части мтДНК, специфичный для данной гаплогруппы. Наличие замены G16319A свидетельствует о принадлежности рассматриваемой линии к подгруппе D4b1. Мы полагаем, что варианты с дополнительными заменами С16239T и T16243C составляют отдельный субкластер в рамках этой подгруппы, хотя он не был аннотирован до настоящего времени в составе единого филогенетического дерева мтДНК [van Oven, Kayser, 2009]. Установлено, что основные этапы диверсификации вариантов гаплогруппы D4b1 протекали в популяциях Центральной и Южной Сибири, а также Восточной Азии [Derenko et al., 2010].
Установленное нами присутствие обоих аллельных вариантов гена амелогенина, специфичных для X- и Y-хромосомы, однозначно свидетельствует о мужском поле погребенного, что согласуется с результатами палеоантропологического анализа скелета. Высокая степень сохранности ДНК в о станках позволила реконструировать аллельный профиль 17 STR-локусов Y-хромосомы:
DYS45616
DYS389I13
DYS39024
DYS389II28
DYS45818
DYS1913
DYS385a/b13/16
DYS39313
DYS39110
DYS43912
DYS63524
DYS39216
Y GATA H411
DYS43714
DYS43811
DYS44823
Предсказанная гаплогруппа Q (Q1a)
На основании этих данных с помощью программ- предикторов однозначно определена принадлежность исследуемого варианта Y-хромосомы к гаплогруп-пе Q, а именно к подгруппе Q1a, которая в пределах Евразии в большей степени характерна для современных восточно-евразийских популяций. Наиболее вероятным местом формирования гаплогруппы Q (и, по-видимому, субкластера Q1a) Y-хромосомы является Центральная Азия и Южная Сибирь [Huang et al., 2018].
Таким образом, общий филогенетический анализ показал, что как мтДНК, так и Y-хромосома мужчины из погр. 688 памятника Сопка-2 принадлежат к кластерам, сформировавшимся и распространенным в генофондах популяций более восточных по сравнению с Барабинской лесостепью регионов Сибири и Центральной Азии. Следовательно, рассматриваемый индивид, вероятно, имел восточные генетиче ские корни. Это согласуется с данными физической антропологии о его возможном происхождении из расовой среды более восточных по отношению к Барабе районов Евразии.
Эффективным инструментом для потенциального уточнения векторов генетических связей исследуемого индивида является анализ распространенности рассматриваемых вариантов мтДНК и Y-хромосомы в древних и современных популяциях Евразии. Ввиду наличия большого банка накопленных сравнительных материалов его проводят, в первую очередь, с использованием данных о современных популяциях. Фило-географический анализ показал, что вариант мтДНК с гаплотипом 16223T-16239T-16243C-16319A-16362C, относящийся к гаплогруппе D4b1, является редким компонентом генофонда населения Евразии. Интересно, что данный вариант встречается в современных популяциях как более восточных районов Евразии (по отношению к Западно-Сибирской лесостепи), в частности у тувинцев (а также у имеющих недавнее центрально-азиатское происхождение калмыков) [Derenko et al., 2007], так и более западных, например у башкир Волго-Уральского региона [Bermisheva et al., 2002]. Аналогичные данные получены касательно обнаруженного нами у индивида аллельного профиля STR-локусов Y-хромосомы, относящегося к га-плогруппе Q (Q1a): носители идентичных и близких по структуре аллельных профилей есть как на востоке Евразии (в частности, среди населения Китая), так и в Восточной Европе (например, у поляков).
Таким образом, хотя филогенетически данные (т. е. анализ на уровне филогенетических кластеров) по однородительским маркерам с материнским (мтДНК) и отцовским (Y-хромосома) наследованием указывают на восточные генетические корни исследуемого нами пришлого индивида, филогеографический анализ конкретных структурных вариантов мтДНК и Y-хромосомы, проведенный с использованием данных о генофонде современных популяций Евразии, не позволяет однозначно определить его возможное географическое происхождение. Это кажущееся противоречие находит объяснение, если при интерпретации результатов учитывать исторический (археологический) контекст рассматриваемых материалов, а также дополнить филогеографический анализ имеющимися данными по древним популяциям Евразии.
Как было отмечено, погр. 688 могильника Сопка-2 археологическими методами датируется IV–V вв. н.э. Возраст гаплогруппы D4b1 23,9–31,5 тыс. лет, а минимальный возраст выделенных в ее составе субкластеров существенно варьирует, но даже у самых молодых составляет не менее 2,5 тыс. лет, т.е. они значительно древнее рассматриваемого погребального комплекса [Derenko et al., 2010]. Таким образом, процессы диверсификации кластера D4b1 мтДНК, протекавшие, по филогенетическим данным, на территории более восточных регионов Евразии, происходили намного раньше совершения рассматриваемого погребения (это справедливо и для Q1a гаплогруппы Y-хромосомы). Следовательно, говоря о восточном происхождении кластеров мтДНК и Y-хромосомы, выявленных у исследуемого индивида, мы подразумеваем в первую очередь его глубокие генетические корни, связывающие, наиболее вероятно, с южными районами Сибири и Центральной Азией в целом, что согласуется с палеоантропологическими данными [Молодин, Чикишева, 1990].
После формирования интересующих нас кластеров мтДНК характер их распространения в Евразии мог существенно меняться. Сооружению погр. 688 могильника Сопка-2 предшествует целое тысячелетие (как минимум с середины I тыс. до н.э.), охватывающее скифскую и гунно-сарматскую эпохи, на протяжении которых в евразийских степях и на сопредельных территориях (включая лесостепной пояс) происходила интенсификация перемещения популяций (и соответствующих специфических генетических компонентов) в различных направлениях (см., напр.: [Боталов, 2000, с. 207–223]). Основной движущей силой этих перемещений были многочисленные группы ранних кочевников. Одним из ключевых направлений являлось распространение генетических компонентов, свойственных восточным номадам, в западные районы Евразийской степи. Свидетельства этих процессов на уровне генофонда мтДНК к настоящему моменту получены как для скифского [Unterländer et al., 2017; Pilipenko, Trapezov, Cherdantsev et al., 2018], так и для гунно-сарматского времени [Pilipenko, Cherdantsev, Trapezov et al., 2018]. Таким образом, к финальной стадии гунно-сарматского времени (к которой и относится погр. 688 могильника Сопка-2) варианты мтДНК и Y-хромосомы, изначально сформировавшиеся в Восточной Евразии, могли быть широко распространены в пределах всего Евразийского степного пояса, хотя их частота в генофондах популяций восточной части региона должна была оставаться выше, чем в западной. В связи с крайне высокой подвижностью кочевого населения перемещение генетического материала (как и археологического) нередко могло иметь характер «броуновского движения» [Боталов, 2000, с. 285, рис. 19].
Привлечение к филогеографическому анализу данных по древним популяциям Евразии позволяет проиллюстрировать описанные выше затруднения с географической локализацией происхождения конкретного мигранта по его генетическим характеристикам. Вариант мтДНК с гаплотипом 16223T-16239T-
16243C-16319A-16362C, относящийся к гаплогруппе D4b1, среди исследованных методами генетики древних популяций Сибири был выявлен у двух носителей пазырыкской культуры скифского времени в Горном Алтае [Unterländer et al., 2017] и у погребенного из могильника булан-кобинской культуры Степуш-ка-1 в предгорной части Алтая (неопубликованные данные авторов). Таким образом, получены прямые свидетельства его присутствия в генофонде населения скифского и гунно-сарматского времени сопредельных с Барабой районов Южной Сибири (к сожалению, древнее население Тувы, в т.ч. оставившее могильник Кокэль, где найдены останки индивидов с тем же краниотипом, что и у рассматриваемого погребенного из Сопки-2, пока остается практически неисследованным методами палеогенетики). Вместе с тем на вероятное распространение данного варианта мтДНК далеко на запад Евразийского степного пояса в эпохи, предшествовавшие сооружению погр. 688 могильника Сопка-2, указывает факт его обнаружения в генофонде классических скифов Северного Причерноморья (!) [Der Sarkissian, 2011].
Проведение аналогичного филогеографического анализа для выявленного нами варианта гаплогруп-пы Q (Q1a) в настоящее время затруднительно как в связи с недостаточным объемом литературных данных о разнообразии Y-хромосомы в древних популяциях различных регионов Евразии, так и из-за отсутствия у нас данных о статусе филогенетически информативных однонуклеотидных полиморфизмов (ОНП) Y-хромосомы. Их получение – задача будущих исследований. На данном этапе можно отметить, что носители различных субкластеров гаплогруппы Q1a Y-хромосомы достоверно выявлены среди представителей древних кочевых групп гунно-сарматского времени на территории Северного Китая [Zhao et al., 2010] и Горного Алтая [Allentoft et al., 2015].
Таким образом, восточный вектор глубоких генетических связей (корней) индивида из погр. 688 не вызывает сомнений. Однако с учетом результатов фило-географического анализа мы не можем однозначно сказать, из какой именно части степного пояса – западной или восточной – этот индивид прибыл на территорию Западно-Сибирской лесостепи (или локализовать его непосредственных предков), основываясь на имеющихся палеогенетических данных. Присутствие близких (и идентичных) вариантов гаплогруппы D4b1 мтДНК и гаплогруппы Q1a Y-хромосомы у представителей населения скифского и гунно-сарматского времени в сопредельных с востока районах Южной Сибири (Горный Алтай), а также конкордантность восточно-евразийских по происхождению вариантов мтДНК и Y-хромосомы, выявленных у рассматриваемого индивида, свидетельствуют в пользу его прихода из более восточных районов. Однако широкая рас- пространенность «восточных» генетических линий к позднему периоду гунно-сарматской эпохи делает возможными и другие сценарии, позволяющие объяснить видимое противоречие между данными физической палеоантропологии и генетики о восточных корнях индивида из погр. 688 могильника Сопка-2 и западными аналогиями для сопроводительного инвентаря из этого погребения. В частности, потомок тех, кто мигрировал в предшествующие эпохи из восточных районов степного пояса Евразии в западные (т.е. имеющий восточно-евразийские генетические характеристики), мог прибыть на территорию Бара-бинской лесостепи из западных степных районов, уже являясь носителем иных, в данном случае сармато-аланских культурных традиций, что отразилось в сопроводительном инвентаре рассматриваемого погребения. В таком случае археологические характеристики комплекса могут с большей точностью указывать на регион, из которого мигрировал индивид.
Заключение
Таким образом, особенности распространения генетических компонентов, возникающие в результате взаимодействия удаленных популяций, существенно усложняют определение географического происхождения конкретного мигранта (или мигрантов) методами палеогенетики. Это ограничение распространяется и на использование методов физической палеоантропологии. Данная проблема прежде всего касается древнего кочевого населения степного пояса и сопредельных территорий Евразии раннего железного века и последующих эпох, особенно в случае палеогенетического и/или палеоантропологического анализа единичных (или немногочисленных) пришлых индивидов. Одним из эффективных способов частичного преодоления этого ограничения является использование репрезентативного популяционного материала (при его наличии). Увеличить разрешающую способность генетического подхода к исследованию единичных индивидов (или небольших серий) позволяет получение их более полной генетической характеристики. Однако даже анализ ядерного генома одного индивида (или маленькой серии) не снимает ограничений полностью. Общим решением данной проблемы является проведение разностороннего исследования материала с привлечением широкого круга методов и подходов. Полноценное использование имеющегося арсенала естественно-научных методов (биологических, химических и физических) и обязательный корректный учет археологического (исторического) контекста исследуемых материалов при интерпретации результатов анализа позволят преодолеть методологические ограничения, имеющиеся у каж- дого подхода, и получить максимально объективные данные об изучаемом археологическом феномене, а следовательно, максимально приблизить нас к реконструкции конкретного исторического события.
Исследование выполнено за счет гранта Российского научного фонда (проект № 17-78-20093). Использование па-леогенетической инфраструктуры ИЦиГ СО РАН обеспечено средствами государственного задания по проекту № 0324-2018-0016.
Список литературы Уникальное захоронение воина гунно-сарматского времени в Западно-Сибирской лесостепи: результаты палеогенетического анализа
- Боталов С.Г. Поздняя древность и средневековье//Древняя история Южного Зауралья. -Челябинск: Изд-во Юж.-Урал. гос. ун-та, 2000. -Т. II: Ранний железный век и средневековье. -С. 207-430.
- Молодин В.И., Елагин В.С. Погребения сперановского этапа потчевашской культуры Барабы//Мировоззрение финно-угорских народов. -Новосибирск: Наука, 1990. -С. 140-149.
- Молодин В.И., Чикишева Т.А. Погребение воина IV-V вв. н.э. в Барабе//Военное дело древнего и средневекового населения Северной и Центральной Азии. -Новосибирск: ИИФФ СО АН СССР, 1990. -С. 161-179.
- Пилипенко А.С., Ромащенко А.Г., Молодин В.И., Куликов И.В., Кобзев В.Ф., Поздняков Д.В., Новикова О.И. Особенности захоронения младенцев в жилищах городища Чича I Барабинской лесостепи по данным анализа структуры ДНК//Археология, этнография и антропология Евразии. -2008. -№ 2. -С. 57-67.
- Пилипенко А.С., Трапезов Р.О., Полосьмак Н.В. Па-леогенетическое исследование носителей пазырыкской культуры из могильника Ак-Алаха-1 (Горный Алтай)//Археология, этнография и антропология Евразии. -2015. -Т. 43, № 4. -С. 144-150.
