Видовая принадлежность возбудителя фомопсиса подсолнечника в выборке изолятов, собранных на некоторых полях в окрестностях Краснодара

Автор: Арасланова Н.М., Антонова Т.С., Старина А.А., Науменко Д.Д., Саукова С.Л., Ивебор М.В.
Рубрика: Агрохимия, агропочвоведение, защита и карантин растений
Статья в выпуске: 3 (191), 2022 года.
Бесплатный доступ
Распространенность опасного карантинного патогена - возбудителя фомопсиса подсолнечника (Phomopsis helianthi Munt.-Cvetk. Mihaljc. & M. Petrov), варьирует в зависимости от погодных условий и запаса его инфекционного начала в почве. В 2010 г. было предложено сохранить название возбудителя Diaporthe, которое было озвучено в литературе раньше всех потенциальных синонимов. К настоящему времени из 14 видов Diaporthe, которые могут быть потенциальными возбудителями фомопсиса подсолнечника в России, кроме D. helianthi, обнаружены виды: D. phaseolorum и D. gulyae, D. monetii и D. vangoghii sp. (подтверждено молекулярногенетическими исследованиями). Существует вероятность распространения этих видов фомопсиса в России с зараженным посевным материалом. В посевах подсолнечника окрестностей Краснодара были отобраны образцы стеблей и семянок с характерными для Diaporthe helianthi симптомами поражения с целью уточнения видовой принадлежности возбудителя болезни. Всего было выделено в чистую культуру 39 изолятов возбудителя болезни. Исследование культуральных признаков проводили согласно методикам экспериментальной микологии. Изоляты возбудителя различались по культуральным характеристикам, однако в пикнидах формировались только P-споры, что характерно для Diaporthe helianthi. Выполнена идентификация видовой принадлежности 17 изолятов возбудителя болезни в отделе секвенирования ЦКП «Биотехнология». Из них 16 изолятов с 99-100%-ной вероятностью были определены как Diaporthe helianthi, и один - как Xylaria ellisii.
Подсолнечник, фомопсис, изолят, возбудитель болезни, видовая принадлежность, культуральные признаки, анализ днк
Короткий адрес: https://sciup.org/142236122
IDR: 142236122 | УДК: 633.854.78:632.4:575 | DOI: 10.25230/2412-608X-2022-3-191-50-59
Species membership of the sunflower stem blight isolates collected in some fields near Krasnodar
The prevalence of a dangerous quarantine pathogen, the sunflower stem blight pathogen (Phomopsis helianthi Munt.-Cvetk. Mihaljc. & M. Petrov), varies depending on weather conditions and the reserves of its infectious matter in the soil. In 2010, it was proposed to keep the name of the pathogen Diaporthe, which was used in the literature before all potential synonyms. Currently, of the 14 species of Diaporthe that can be potential pathogens of sunflower stem blight in Russia, there have been identified species other than D. helianthi: D. phaseolorum and D. gulyae, D. monetii and D. vangoghii sp. (confirmed by molecular genetic studies). There is a possibility that these stem blight species will spread in Russia with infected seed material. We selected stem and seed samples with the affection symptoms characteristic of Diaporthe helianthi in sunflower sowings from the fields near Krasnodar to clarify the species membership of the pathogen. We identified a total of 39 isolates of the pathogen as a pure culture. We conducted the study of cultural features according to the methods of experimental mycology. Isolates of the pathogen differed in cultural characteristics, but only р-spores were formed in pycnidia, which is characteristic of Diaporthe helianthi. We carried out the identification of the species membership of 17 pathogen isolates in the sequencing department of TsKP "Biotechnology". Of these, 16 isolates were identified with 99-100 % probability as Diaporthe helianthi and one isolate as Xylaria ellisii.
Текст научной статьи Видовая принадлежность возбудителя фомопсиса подсолнечника в выборке изолятов, собранных на некоторых полях в окрестностях Краснодара
Введение. Фомопсис распространен на подсолнечнике ( Helianthus annuus L.) во всех странах мира, возделывающих эту культуру [1; 2] . Вредоносность болезни зависит от погодных условий. Потери урожая могут составлять от 5 до 35 %, а в годы эпифитотий – до 10 %. С 1986 г . возбудитель этой болезни Phomopsis helianthi Munt.-Cvetk. Mihaljc. & M. Petrov входит в «Единый перечень карантинных объектов, ограниченно распространенных на территории Евразийского экономического союза» [2].
Впервые возбудитель фомопсиса был обнаружен в США в 1932 г. и определен как Phomopsis arctii Lasch на видах подсолнечника: H. annuus и H. giganteus [3]. В 1933 г. Вехмайер, основываясь на морфологии и ассоциации с растениями - хозяевами, высказал предположение, что возбудителем болезни может быть комплекс видов Diaporthales [4].
По результатам таксономической ревизии в международной базе данных по фитопатогенам возбудителем фомопсиса подсолнечника зарегистрирован гриб Phomopsis helianthi Munt.-Cvetk. Mihaljc. & M. Petrov с телеоморфной стадией Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov [5] . Этот вид возбудителя долгое время считался единственным, вызывающим фомопсис подсолнечника в регионах РФ [6] . Позже из семян и стеблей подсолнечника был выделен Phomopsis arctii Lasch , который ранее был обнаружен на сорных растениях в Краснодарском крае [6; 7].
С обнаружением новых видов возбудителя фомопсиса на подсолнечнике, которые были представлены анаморфами Diaporthe ( Ascomycota, Diaporthales,
Valsaceae ), предложено сохранить название возбудителя Diaporthe, которое озвучено в литературе раньше всех потенциальных синонимов [8; 9].
Филогенетический анализ, проведенный в некоторых странах, выявил новые виды возбудителя фомопсиса, которые в GenBank ошибочно числились как D. helianthi: D. gulyae Shivas, D. kochmanii Shivas, D. kongii Shivas [10], D. longicolla (Hobbs), D. stewartii Harrison [12; 13; 14] и другие.
Установлено, что виды фомопсиса D. helianthi и D. gulyae доминируют по распространенности в мире [12; 13; 14] . По сравнению с другими видами D. helianthi чаще встречается в странах Европы и Южной Африки и поражает дикорастущий и культурный подсолнечник, а также Xanthium strumarium , Arctium lappa и Vitiss vinifera, но не был обнаружен в Австралии [12]. D. gulyae распространен в США и Канаде, поражает, кроме подсолнечника, сою ( Glycine max L.) и чертополох ( Carduus tribulus ) [13; 14].
Симптомы болезни при поражении стеблей подсолнечника видом D. gulyae идентичны, вызываемым D. helianthi: пятно неправильной формы от бледно - до темно - коричневого цвета с центром в листовых узлах с повреждением сердцевины и с часто встречающимся переломом в этом месте [12; 13] . Однако разная видовая принадлежность была подтверждена анализом нуклеотидной последовательности ДНК [15].
Подробное описание культуральных характеристик D. helianthi было сделано югославскими учеными в 80-е годы прошлого столетия [5]. Позже проведено сравнительное исследование географических изолятов из разных стран [6]. Изоля-ты из Югославии, Болгарии, Украины, юга РФ в культуре на КГА (картофельноглюкозный агар) формировали белый, компактный, порошистый, полупогру-женный в субстракт мицелий с темной пигментацией в центре колонии. Изолят из Франции имел слабый, белый воздушный мицелий и выделялся крупными, высокими пикнидами. В пикнидах гео- графических изолятов присутствовали только β-споры [5; 6].
В 2018 г. были описаны характерные культуральные признаки для D. helianthi , паразитирующего на подсолнечнике в США: колонии на картофельно - декстрозном агаре (КДА) имеют белый мицелий наряду с узкими зеленовато желтыми участками. На обратной стороне цвет культуры от белого до коричневого со светло - коричневыми рассеянными пятнами вначале, которые позже становятся темно - коричневыми. Образовавшиеся пикниды обычно агрегированные, реже одиночные. Показано, что изоляты этого вида могут отличаться содержанием разного типа спор в пикнидах. В некоторых присутствуют только одноклеточные, нитевидные β - споры. В других встречаются два типа спор: β - и одноклеточные, бесцветные, эллиптические α - споры [14; 15] . Например, у изолятов D. helianthi из штатов США Миннесота, Северная Дакота и Южная Дакота в пикнидах культур формировались только β - споры, которые не прорастали в лабораторных условиях, перитеции D. helianthi формируются на перезимовавших растительных остатках. Аски бесцветные, 8 - споровые, мешковидные, аскоспоры бесцветные, эллиптические [15] . Колонии изолятов D. gulyae на КДА имели скудный воздушный мицелий и несколько более темные центры. Пикниды часто разбросаны по поверхности мицелия, почти шаровидные, до 3 мм в диаметре, иногда с носиками до 1 мм длиной, α - споры шаровидные, полушаро-видные, эллипсоидные, овальные или обратнояйцевидные, гиалиновые, β - споры не наблюдались в культуре на КДА [15].
В Российской Федерации в последние два десятилетия распространение болезни было локальным и не достигало 40 % [1]. Однако из 14 видов Diaporthe, которые могут быть потенциальными возбудителями фомопсиса подсолнечника в РФ, кроме D. helianthi, обнаружено еще два вида: D. phaseolorum и D. gulyae [16; 17]. В 2022 г. на подсолнечнике выявлены еще три вида Diaporthe: D. eres, D. monetii sp. и D. vangoghii sp. [18]. Частота встречаемости этих видов в регионах РФ не изучена. Велика вероятность их распространения зараженными семенами.
В связи с обнаружением в РФ новых видов возбудителя фомопсиса на подсолнечнике стала необходимой идентификация видовой принадлежности грибов рода Diaporthe в посевах подсолнечника окрестностей г. Краснодара с применением традиционных и современных молекулярных методов.
Цель нашей работы – определить видовую принадлежность возбудителя болезни, выделенного из растений и семян подсолнечника с полей ВНИИМК с признаками поражения фомопсисом.
Материалы и методы. Исследования проводили в 2021–2022 гг. в лаборатории иммунитета центральной экспериментальной базы ФГБНУ ФНЦ ВНИИМК.
На полях подсолнечника с разных генотипов в окрестностях Краснодара в период налива семян было собрано 37 фрагментов стеблей и два образца семянок с различными симптомами поражения фомопсисом. Высечки (длиной 1 см) из фрагментов пораженных стеблей подсолнечника промывали водопроводной водой в течение 2 мин и стерилизовали в 95%- ном спирте, ополаскивали стерильной водой, обжигали в пламени спиртовки, раскладывали в чашки Петри (диаметр 9 см) на КДА с добавлением антибиотика ампициллина. Инкубировали в термостате при температуре 26 оС до начала образования пикнид. Диаметр колонии измеряли на 3 - й и 7 - й дни культивирования, на 14 - й день подсчитывали количество образовавшихся пикнид. Тип спор определяли микроскопированием экссудата пикнид с помощью цифрового микроскопа Motic.
Идентификация видовой принадлежности 17 изолятов возбудителя фомопси-са была проведена на коммерческой основе в Федеральном государственном бюджетном научном учреждении «Всероссийский научно-исследовательский институт сельскохозяйственной биотехнологии» (ЦКП «Биотехнология» ВНИИСБ, Москва). Для идентификации патогена были использованы области внутренних транскрибируемых спейсеров рДНК (ITS-локус). Секвенирование образцов проводили по методу Сэнгера на генетическом анализаторе 3500 (Applied Biosystems, США). Нуклеотидные последовательности участков генов изучаемых изолятов были проанализированы с помощью программного обеспечения «BioEdit» и базы данных.
Результаты и обсуждение. В условиях одного и того же года распространенность и интенсивность болезней связаны с разной степенью индивидуальной устойчивости растений и разнообразием генотипов подсолнечника. Типичные симптомы поражения вегетирующих растений подсолнечника возбудителем фо-мопсиса в 2021 г. представлены на рисунке 1.

а б в

г д е ж
Рисунок 1 – Поражение подсолнечника фомопсисом ( Diaporthe helianthi Munt.-Cvetk. Mihaljc. & М. Petrov ): а , б, в – обширные участки поражения стебля, различаются по цвету; г, д, е – небольшие участки поражения у основания стебля; ж – перелом стебля в средней части (ориг.)
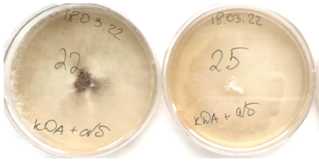
Выделенные из пораженных участков стеблей подсолнечника изоляты возбудителя фомопсиса различались по скорости роста мицелия в культуре и формирования пикнид. В таблице 1 представлены характерные культуральные признаки, свойственные изученным изолятам возбудителя болезни. Колонии большинства изолятов имели белый, порошистый мицелий, а у двух – войлочный (рис. 2).
Таблица 1
Характерные культуральные признаки изолятов возбудителя болезни, выделенных из растений с симптомами поражения DiaporthehelianthiMunt.-Cvetk.Mihaljc.& M. Petrov на 14 - й день выращивания
|
Характеристика культуры |
Диаметр колонии, мм |
Количество пикнид, шт. на 14-й день |
|
|
3-й день |
7-й день |
||
|
Мицелий порошистый, пикниды простые и сложные, в стадии формирования |
25 |
85 |
40 |
|
Мицелий порошистый, пикниды мелкие, простые, расположены на поверхности и в субстрате, в стадии формирования |
30 |
85 |
~ 1000 |
|
Мицелий порошистый, тянется вверх, пикниды простые и сложные, зрелые |
25 |
85 |
30 |
|
Мицелий порошистый, пикниды простые и сложные расположены на поверхности, зрелые |
35 |
85 |
80 |
|
Мицелий войлочный, пикнид нет |
15 |
40 |
0 |
|
Мицелий войлочный, пикниды простые и сложные |
25 |
80 |
40 |
|
Мицелий порошистый, пикниды простые и сложные, в начальной стадии формирования |
20 |
75 |
20 |
30.3 \
б
а
Рисунок 2 – Колонии изолятов гриба Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov , выделенного из фрагментов стеблей подсолнечника на питательной среде: а, б – войлочный мицелий;
в – порошистый мицелий (ориг.)
в
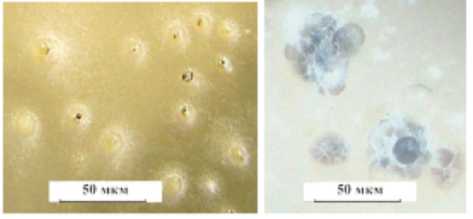
Размер колоний на третьи сутки составлял от 15 до 35 мм. На 7 - й день колонии большинства изолятов выровнялись и составили 80–85 мм. Пикниды формировались простые и сложные (многокамерные) (рис. 3 ). На 14 - й день у многих изолятов наблюдался выход пикноспор, что указывало на их зрелость, тогда как у других пикниды были в стадии формирования. В основном количество пикнид варьировало от 20 до 60, а у восьми изо-лятов их было больше 1000 штук. Выделялся изолят, который отличался отсутствием пикнид в культуре.
а б
Рисунок 3 – Пикниды простые (а) и сложные многокамерные (б), образовавшиеся в культурах изолятов фомопсиса на картофельно - декстрозном агаре (ориг.)
Известно, что для изолятов D. helianthi из разных географических зон характерно присутствие в пикнидах либо только β -, либо в разном соотношении β - и α - спор [2; 10; 13] . Как уже отмечено выше, у D. gulyae на КДА в пикнидах формируются только α - споры, а у других видов присутствуют как α - , так и β - споры [14; 15] . В пикнидах всех изученных нами изолятов фомопсиса присутствовали только β - спо - ры (рис. 4)
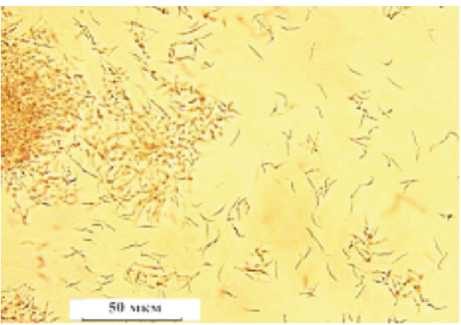
Рисунок 4 – Выход β - спор Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov из пикнид ( ориг .)
Для образования совершенной стадии спороношения гриба, после хранения фрагментов стеблей при -20 оС, их поместили во влажную камеру, где на 20 - е сутки образовались перитеции с аскоспорами. Перитеции образовались и на семянках подсолнечника, которые не подвергались замораживанию (рис. 5).
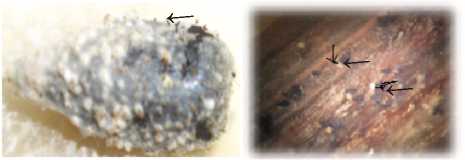
Рисунок 5 – Перитеции Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov на семянке и фрагменте стебля подсолнечника (ориг.)
Аски и вышедшие из них аскоспоры были типичными для D. helianthi. Один изолят отличался от всех остальных отсутствием пикнид в культуре. Вместо них на колонии появились очень похожие на перитеции образования (рис. 6).

Рисунок 6 – Колония гриба, выделенного из стебля подсолнечника с признаками поражения Diaporthe helianthi Munt.-
Cvetk. Mihaljc. & M. Petrov (ориг.)
По изученным культуральным характеристикам изоляты возбудителя болезни, кроме одного, у которого отсутствовали пикниды, можно отнести к Diaporthe helianthi.
С применением анализа ДНК изолят, который выделялся отсутствием пикнид в культуре, был определен как Xylaria ellisii J.B. Tanney, Seifert & Y.M. Ju. X. Ellissi относится к эндофитным грибам семейства Xylariaceae, часто выделяемым из Picea и Pinus, а также разлагающихся ветвей или стволов Acer saccharum [19]. Из вегетирующего растения подсолнечника с симптомами фомопсиса X. ellisii выделен нами впервые.
Результаты идентификации части образцов в отделе секвенирования ЦКП «Биотехнология» представлены в таблице 2.
Шестнадцать изолятов вобудителя болезни с 99– 100%- ной вероятностью были отнесены к виду Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov . У остальных 22 изолятов анализ нуклеотидных последовательностей ДНК не проводился. Однако их культуральные признаки (в частности, наличие в пикнидах только β - спор) совпадают с представленными в таблице 1. Поэтому мы полагаем, что и эти изоляты можно отнести к виду Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov.
Заключение. Симптомы проявления фомопсиса на вегетирующих растениях подсолнечника в полях окрестностей г. Краснодара характерны для Diaporthe helianthi Munt.-Cvetk. Mihaljc & M. Petrov. Выделенные из пораженных фомопсисом стеблей и семянок подсолнечника изоля-ты различались в культуре на КДА скоростью роста мицелия и количеством сформированных пикнид. Однако в пикнидах у всех изолятов не было α - спор и присутствовали только β - споры, что является характерным признаком D. helianthi. Анализ нуклеотидных последовательностей ДНК 16 изолятов возбудителя подтвердил с 99– 100%- ной вероятностью принадлежность их к виду Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov.
Таблица 2
Нуклеотидные последовательности ДНК исследуемых изолятов возбудителя болезни, выделенных из вегетирующих растений и семян подсолнечника с симптомами поражения Diaporthe helianthi Munt.-Cvetk. Mihaljc. & M. Petrov
Отдел секвенирования ЦКП «Биотехнология»
|
Последовательность |
Видовая принадлежность |
|
GGAGGGATCATTGCTGGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGC CGGCCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGA GTTAAAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATA AGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGT TCGAGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAG CCCGCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCCTGAA AATTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAG |
Diaporthe helianthi KU315382.1 100 % 5 41 п.о |
|
GAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGCCCCCCCTGGGGG CCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAGTTAAAAACACAAATG AATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAG AATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAAC CCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGA GCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCCTgAAAATTTgACCTCGGATCA |
Diaporthe helianthi KU315382.1 100 % 500 п.о |
|
GCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGCCCCCCCTGGGGGCCCC CTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAGTTAAAAACACAAATGAATC AAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATT CAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAACCCTC AAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGAGCGT AGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCC |
Diaporthe helianthi KU315382.1 100 % 475 п.о |
|
GCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGCCCCCCCTGGGGGCCCC CTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAGTTAAAAACACAAATGAATC AAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATT CAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAACCCTC AAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGAGCGT AGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCCTgAAAATTTgACCTCGGATCA |
Diaporthe helianthi KU315382.1 100 % 496 п.о |
|
CCSCCCTGGGGRCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACaGTGGATCTcTGAGTTAA AAACACAAaTGAATCAAwACTTTCAACAACGGATCTCTtGGTTCTGGCATCGATGAAGAACGCAGCGAAATgCGATAAGTAAW GgTGAATTGCAGAATTCAGTGAATCATCGAATCTTTSAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCKGTTCGAG CGTCATTTCAACCCTCAAGCCtGGSTTGGTGATGGGGCACTGCCKGTGACAGGGCAGGCCCYGAAATCCAGCGGCGAgCCCSCC GGGMCCCCGAGCGTAGTAGKAACTTCTcGCTCCGGAAGGCCCTGGCGGCGCCCTGCCGTTAAACCCCCACTCCTGAAAATTTG ACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGCATATCAWAAAGCGGAGGA |
преобладание Diaporthe helianthi KU315382.1 99 % 473 п.о |
|
CTTGTTTCTMCGTGGATCTCTGAKTTAWAAACACWATGAATCMAACTTTCAACAACGGATCTCTTGGKTCTGGCATCGATGAA KAACGCARCGAAATGCGATAAGTAATGTGAATTGCASAATTCAKYGAATCATCSAATCTTTGAACGCACATTGCGCCCTCTGG TATTCCGGAGGGCATGCCTGTTCAGCGYCATTTCAACCCTCARCCTGGCTTGGTGATGGGGCACTGCCTGYGACAGGGCAGGM CCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCTGGCGGCGCCCTGCCG TTAAACCCCCACTCCTGAAAATTTGACCTCGGATCAKGTASGAATACCCGCTGAACTTAAGCATATCATAAGCGGAGG |
Diaporthe helianthi KU315382.1 100 % 5 41 п.о |
|
GGAGGGATCATTGCTGGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGC CGGCCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGA GTTAAAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATA AGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGT TCGAGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAG CCCGCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCCtGAA AAKTtGACCTCGGATCAGGTAGGAATACCCGCTGAACT |
Diaporthe helianthi KU315382.1 100 % 410 п.о |
|
GGAGGATCATGCTGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGG CCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCaAACTCTTGTTTCTACAGTGGATCTCTGAGTT AAAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGT AATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCG AGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCC GCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTaAACCCCCAACTCCTgAAAATT TGACCTCGGATC |
Diaporthe helianthi KU315382.1 100 % 5 12 п.о |
Продолжение таблицы 2
|
GGATCATAAAGAGTTCTATAACTCCCAAACCCATGTGAACaTACCTTACGTTGCCTCGGCaGGTCGCGCCTACCCCGTAACGTC CTACCCTGTAGGACCTACCCGGTAGACGCGGGTAAGCCTGCCGGCGGCCCACGAAACTCTGTTTAATATTGAATTCTGAACCT AAAACTAAATAAGTTAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAAT GTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCATTAGTATTCTAGTGGGCATGCCTGTTCGAGC GTCATTTCAACCCTTAAGCCTCTGTTGCTTAGTGTTGGGAGCCTACGGCACCCGTAGCTCCTCAAAGTTAGTGGCGGAGTCGGT TCACACTCTAGACGTAGTAATTCTTATCTCGCCTATCAGTTGgACCGGTCCCCTgCCGTAAAACCCCCCATTTTTAAAGGTTGAC CTCGGATCAGGTAGGAATACCCGCTGAACTAG |
Xylaria ellisii MN219731.1 100 % 535 п.о |
|
CGGAGGATCATGCTGGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCC GGCCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAG TTAAAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAA GTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTT CGAGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGC CCGCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAaCTCCTGAAA ATTTGACCTCGGATCATGYACGAATACCCGCTGAACTAG |
Diaporthe helianthi KU315382.1 99 % 538 п.о |
|
CTGGAACGCGCCCCCGGCGCACCCAGAAACCCtTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGCCCCCCCTGGG GGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTcTGAGTTAAAAACACAAA TGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGC AGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCA ACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCC GAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAaCTCCTGAAAATTTGACCTCGGAT CAGGTASGAATACCCGCTGAACTTAGCA |
Diaporthe helianthi KU315382.1 100 % 529 п.о |
|
GAACGCGCCCCCGGCGCACCCAGAAACCCTTGTGAACTTATACCTATCTGTTGCSTCGGCGCAGGCCGGCCCCCCCTGGGGGC CCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTgAGTTAAAAACACAAATGA ATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGA ATTCAGTGAATCATCGAATCTTTgAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAACC CTCAaGCCTGGCtTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCYGAAATCCAGCGGCGAGCCCRCCGGGACCCCGAGC GTAGTAGTAACTTCTCGCTCCGGAAGGCCCTGGCGGCGCCCTGCCGTTAAACCCCCAACTCCTGAAAATTTGACCTCGGATCAT GTASGAATACCCGCTGAACTA |
Diaporthe helianthi JF430484.1 99 % 52 1 п.о |
|
GCGGAGGGATCATTGCTGGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCG GCCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAGTTAAA AACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTG CAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAACCC TCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGAGCGTAG TAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAACCCCCAACTCCTgAAAATTTGACCTCGGATCAGGTAGGAATA CCCGCTGAACTTAAG |
Diaporthe helianthi KU315382.1 100 % 5 43 п.о |
|
GGGATCATTGCTGGAACGCGCCCCCCGGCGCRCCCAGAAACCYTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGG CCCCCCCTGGGGGCCCCCTGGGAACcAGGGAGCAGCCCGCCGGCGGCCGACCaAACTCTTGTTTCTACAGTGGATCTcTGAGTT AAAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGT AATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCG AGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCC GCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTAAaCCCCCAACTCCTGAAAAT TTGACCTCGGATMGGGTACGAATACCCGCTGAAC |
Diaporthe helianthi KU315382.1 100 % 534 п.о |
|
GAGGATCATGCTGAACGCGCCCCCGGCGCACCCAGAAACCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGcC CCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGCCGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTcTGAGTTA AAAACACAAATGAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTA ATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTtGAACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGA GCGTCATTTCAACCCTCaAGCCTGGCTTGGTGATGGGGCACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCG CCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGAAGGCCCtGGCGGCGCCCTgCCGTTaAACCCCCAACTCCTgAAAATTT gACCTCGGATCAGTAGATACCCGCTGAAC |
Diaporthe helianthi JF430484.1 99 % 530 п.о |
|
GAAGTAAAAGTCGTAACAAGGTCTCCGTTGGTGAACCAGCGGAGGGATCATTGCTGGAACGCGCCCCCGGCGCACCCAGAAA CCCTTTGTGAACTTATACCTATCTGTTGCCTCGGCGCAGGCCGGCCCCCCCTGGGGGCCCCCTGGGAACAGGGAGCAGCCCGC CGGCGGCCGACCAAACTCTTGTTTCTACAGTGGATCTCTGAGTTAAAAACACAAATGAATCAAAACTTTCAACAACGGATCTC TTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGA ACGCACATTGCGCCCTCTGGTATTCCGGAGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCCTGGCTTGGTGATGGGGC ACTGCCTGTGACAGGGCAGGCCCTGAAATCCAGCGGCGAGCCCGCCGGGACCCCGAGCGTAGTAGTAACTTCTCGCTCCGGA |
Diaporthe helianthi AJ312362.1 AJ312354.1 99.83 % |
Список литературы Видовая принадлежность возбудителя фомопсиса подсолнечника в выборке изолятов, собранных на некоторых полях в окрестностях Краснодара
- Обзор Фитосанитарного состояния посевов сельскохозяйственных культур в Краснодарском крае в 2021 году. Прогноз развития вредных объектов в 2022 году. Министерство сельского хозяйства РФ. Филиал Федерального государственного бюджетного учреждения «Российский сельскохозяйственный центр» по Краснодарскому краю. - Краснодар, 2022. - 108 с.
- Якуткин В.И. Фитосанитарная обстановка с фомопсисом на посевах подсолнечника в России и странах СНГ, влияние источников инфекции болезни на ее состояние // The scientific heritage. - 2020. - No 50. - P. 5-15.
- Harveson R., Mathew F., Gulya T., Markell S., Block Ch. and Thompson S. Sunflower stalk diseases initiated through leaf infection. DOI: 10.1094/PHP-12-17-0083-DG.
- Wehmeyer L.E. The genus Diaporthe nitschke and its segregates. - Ann Arbor, University of Michigan Press, 1933. - P. 23-33.
- Muntanola-Cvetkovic M., Mihaljcevic M., Petrov M. On the identity of causative agent of a serious Diaporthe disease in sunflower plants // Nova Hedwigia. - 1981. - No 34. - P. 417-435.
- Долженко Е.Г. Биология гриба Phomopsis helianthi и меры борьбы с ним в условиях Краснодарского края: автореф. дис. канд. биол. наук / Евгения Григорьевна Долженко. - Краснодар: КубГАУ, 2000. - 25 с.
- Пивень В.Т., Алифирова Т.П., Шуляк И.И., Мурадасилова Н.В., Саенко Г.М. Семена подсолнечника - источник сохранения и распространения фомопсиса // Защита и карантин растений. - 2010. - № 1. - С. 36-40.
- Santos J.M., Correia V.G., Phillips A.J.L. Primers for mating-type diagnosis in Diaporthe and Phomopsis: their use in teleomorph induction in vitro and biological species definition // Fungal Biology. - 2010. - Is. 114 (2-3). - Р. 255-70. DOI: 10.1016/j.funbio. 2010.01.007.
- Herr L.J., Lipps P.E., Watters B.L. Diaporthe stem canker of sunflower // Plant Disease. - 1983. - 67. - P. 911-913.
- Yang S.M., Berry R.W., Luttrell E.S., Vongkaysone T. A new sunflower disease in Texas caused by Diaporthe helianthi // Plant Disease. - 1984. - 68. - P. 254-255.
- Catalogue of Life: 2020-04 Beta. Annual Checklist: [Электронный ресурс]. - Режим доступа: https://www.catalogueoflife.org/.
- Thompson S.M., Tan Y.P., Young A.J., Neate S.M., Aitken E.A.B., Shivas R.G. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species // Persoonia. -2011. - No 27. - P. 80-89. DOI: 10.3767/003158511X617110.
- Mathew F.M., Alananbeh K.M., Balbyshev N., Jordahl J.D., Meyer S.M., Castlebury L., Gulya T. and Markell S.G. Phomopsis stem canker: A reemerging threat to sunflower (Helianthus annuus) in the United States // Phytopathology. -2017. - 195. - P. 900-907. DOI: 10.1094/PHYTO-11-14-0336-FI.
- Olson T.R., Kontz B., Gulya T.J. and Mathew F.M. First report of Diaporthe stewartii causing Phomopsis stem canker of sunflower (Helianthus annuus) in Minnesota // Plant Dis. -2017. - Vol. 101. - No 2. - P. 382. DOI: 10.1094/PDIS-08-16- 1122-PDN.
- Mathew F.M., Olson T.R., Marek L.F., Gulya T.J., Markell S.G. Identification of sunflower (Helianthus annuus) accessions resistant to Diaporthe helianthi and Diaporthe gulyae // Plant Health Prog. - 2018. - Vol. 19. - P. 92-96. DOI: 10.1094/PHP- 10-17-0060-RS.
- Гомжина М.М., Ганнибал Ф.Б. Первая находка гриба Diaporthe phaseolorum на подсолнечнике в России. - 2018. - Т. 5. - № 1. - С. 59-64.ttps:// Microbiology Independent Research Journal. - 2018. - Vol. 5. - No 1. - P. 59-64. DOI: 10.18527/2500-2236-2018-5-1-59-64.
- Сурина Т.А., Скрипка О.В., Ручка Е.Р. Изучение генетических особенностей возбудителя фомопсиса подсолнечника // Фитоса-нитария. Карантин растений. - 2021. - № 1 (5). - С. 42-48.
- Гомжина М.М., Ганнибал Ф.Б. Виды Diaporthe, поражающие подсолнечник (Helianthus annuus) в России, с описанием двух новых видов // Mycologia. - 2022. - 114 (3). - P. 556-574. DOI: 10.1080/0027514. 2022.2040285.
- Helaly S.E., Thongbai B. & Stadler M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales // Natural Product Reports. - 2018. - Is. 9 (35). - P. 1014. DOI: 10.1039/C8NP00010G.
