Видовые особенности гистологического строения аорты у неясыти обыкновенной, совы полярной и ястреба-тетервятника

Автор: Фоменко Людмила Владимировна, Хонин Геннадий Алексеевич
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Ветеринарные науки
Статья в выпуске: 1 (21), 2016 года.
Бесплатный доступ
Описано гистологическое строение аорты и плечеголовных стволов у неясыти, совы полярной и ястреба-тетеревятника. Установлены морфологические особенности в строении сосудов в зависимости от условий обитания и типов полета. В результате проведенных гистологического исследований аорты, плечеголовного ствола, у хищных птиц установлено, что изученные артерии имеют хорошо выраженные внутреннюю, среднюю и наружную оболочки. Установлено, что аорта исследованных птиц эластического типа, имеет внутреннюю оболочку, состоящая из столбчатых эндотелиоцитов, под которыми находится тонкая прослойка из коллагеновых и эластических волокон, создающих с пучками гладких миоцитов крупноячеистый рисунок. У неясыти обыкновенной и ястреба-тетеревятника эндотелий имеет столбчатый вид. Высота клеток равна их ширине или немного превышает ее. У ястреба-тетеревятника соседние эндотелиоциты аорты между собой контактируют неплотно с появлением между ними узких полостей под эндотелием в виде пузырьков.
Гистология, аорта, плечеголовные стволы, птицы
Короткий адрес: https://sciup.org/142199118
IDR: 142199118 | УДК: 619:611.13:611.737:636.5
Specific features of histological stucture of aorta in ordinary owls, owls and hawks polar teterevyatnika
Described the histological structure of the aorta and brachiocephalic trunk at the owl, owls and polar goshawk. The morphological features in the structure of blood vessels, depending on the environmental conditions and type of flight. As a result of histological examination of the aorta, brachiocephalic trunk, birds of prey found that studied the artery have well-defined inner, middle and outer shell. It was found that the aorta studied birds elastic type, it has an inner shell consisting of columnar endothelial cells, which is a thin layer of collagen and elastin fibers, creating a smooth muscle bundles of the coarse pattern. At Owl ordinary and goshawk endothelium has a columnar appearance. The height of the cell is equal to the width or slightly exceed it. In goshawk aortic endothelial cells adjacent to each other in contact with the emergence of loosely between the narrow cavities under the endothelium in the form of bubbles.
Текст научной статьи Видовые особенности гистологического строения аорты у неясыти обыкновенной, совы полярной и ястреба-тетервятника
Птицы занимают особое положение в мире животных и представляют наиболее многочисленную, хорошо обособленную группу теплокровных позвоночных с интенсивным обменом веществ. Эволюцию птиц следует рассматривать как процесс адаптации, в результате которого они приобрели способность к полету, это коренным образом отличает их от других групп позвоночных.
В дикой природе дневные и ночные хищные птицы как ценная группа охотничьих птиц имеют большое значение для сельского хозяйства и являются санитарами природных зон РФ. Множество диких птиц, находясь в неволе в зоопарках и вольерах, испытывают значительное влияние антропогенных факторов.
Сердце и сосудистое русло органов и тканей обеспечивает их метаболические функции. У хищных диких птиц, приспособленных к полету, имеются приспособительные морфологические механизмы для регуляции гемодинамических функций всего организма. Несмотря на значительные достижения современной морфологии, до сих пор остается ряд проблем, касающихся гистологического строения аорты у хищных диких птиц. В руководствах и пособиях по морфологии птиц, как отечественных [1], так и в зарубежных [2; 3], практически отсутствуют сведения о гистоархитектонике аорты.
Тема исследований определена интересом к хищным диким птицам, обладающим способностью к полету, в частности изучением видовых особенностей строения аорты на гистологическом уровне.
Для гистологического изучения использован классический метод изготовления препаратов [4]. Морфологическую структуру сосудов выявляли с помощью окрашивания срезов гематоксилином Ганзена и эозином. Коллагеновые волокна – окраской по Маллори, а эластические волокна – по Вейгерту. Морфометрические измерения микроструктур проводили с использованием винтового окуляра-микрометра МОВ-1-16. На основании полученных данных рассчитали отношение толщины стенки к диаметру ее просвета. Цифровой материал был подвергнут статистической обработке с использованием компьютерной программы «Microcoft Exсel».
Объекты и методы
Всего изучено 9 трупов птиц из отрядов Совообразные (неясыть обыкновенная и сова полярная) и Соколообразные (ястреб-тетеревятник), относящихся к разным таксономическим группам, приспособленным к различным условиям обитания и использующим разные типы полета. В результате проведенных исследований изучена ангиоархитектоника дуги аорты и плечеголовного ствола
В результате проведенных гистологического исследований аорты, плечеголовного ствола у хищных птиц установлено, что изученные артерии имеют хорошо выраженные внутреннюю, среднюю и наружную оболочки.
У неясыти обыкновенной аорта эластического типа с толщиной стенки 712,0 ± 0,4 мкм. У интимы неглубокие продольные складки с фрагментами гладких мышечных клеток. Одиночные капилляры локализуются близко к эндотелию (рис. 1). Эндотелий имеет различную форму клеток – от уплощенной до почти круглой, что хорошо видно по форме ядер. Под эндотелием находится тонкая прослойка рыхлых соединительных волокон, создающих ячеистый рисунок. Далее лежит внутренняя эластическая мембрана, представленная четкими, толстыми, длинными и параллельными относительно друг друга волокнами. Она на препаратах выделяется своей яркостью и плотностью. Между внутренней эластической мембраной и медией – крупноячеистая структура подэндотелиального слоя. Здесь встречаются пучки гладкой мышечной ткани, ориентированные, судя по форме ядер миоцитов, вдоль аорты, под углом к эндотелию. Между эндотелием и первым слоем медии весь объем занят редко расположенными тонкими, паутинообразными волокнами, занимающими наибольший объем, и фрагментами умеренно окрашенных соединительнотканных волокон. В интиме ядра соединительнотканные клетки локализуются значительно реже, чем в медии и адвентиции.
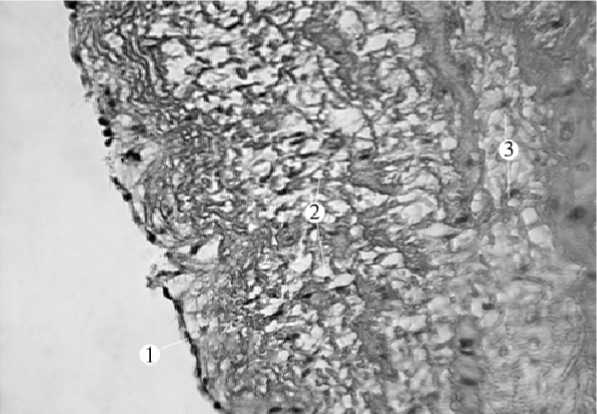
Рис. 1. Интима аорты неясыти обыкновенной (гематоксилин-эозин, ув. х 400); 1 – эндотелий; 2 – кровеносный капилляр; 3 – волокнистая соединительная ткань; 4 – пучки гладких миоцитов.
Таким образом, установлено, что аорта исследованных птиц эластического типа имеет внутреннюю оболочку, состоящую из столбчатых эндотелиоцитов, под которыми тонкая прослойка из коллагеновых и эластических волокон, создающих с пучками гладких миоцитов крупноячеистый рисунок. У неясыти обыкновенной и ястреба-тетеревятника эндотелий столбчатого вида. Высота клеток равна их ширине или немного ее превышает. У ястреба-тетеревятника соседние эндотелиоциты аорты контактируют неплотно, с появлением узких полостей под эндотелием в виде пузырьков.
Гладкая мышечная от соединительной ткани в медии легко дифференцируется, с ровным, умеренно окрашенным фоновым красителем цитоплазмы гладких миоцитов и их ядер. Ядра гладких миоцитов даже при их поперечном сечении сохраняют прозрачность кариоплазмы, но при косом и продольном в ядрах гладких миоцитов четко просматриваются ядрышки и крупные зерна хроматина. Кариолемма ядер гладких миоцитов тонкая и хорошо просматривается только при их продольном сечении. На ровном фоне цитоплазма гладких миоцитов и ее волокнистая структура выражены слабо.
Ядра соединительнотканных клеток, в отличие от ядер гладких миоцитов, всегда окрашиваются плотно. Поэтому ядра с прозрачной кариоплазмой встречаются редко, даже если ядро соединительнотканной клетки имеет прозрачную кариоплазму, ядрышки, хроматин и кариолемма неразличимы. Ядра фибробластов и фиброцитов расположены на слабоокрашенном, рыхлом фоне, состоящем из волокон различной толщины и неупорядоченного направления. Медиа представлена чередующимися слоями соединительной и гладкой мышечной тканей. В направлении к адвентиции толщина мышечных слоев увеличивается, а прослойки соединительной ткани между ними истончаются. Если соединительнотканные прослойки широкие, то коллагеновые и эластические волокна создают ячеистую структуру, подобную той, что в интиме, но размер ячеек в медии значительно меньше, а соединительнотканные волокна более плотные. Ближе к адвентиции в узких соединительнотканных прослойках коллагеновые и эластические волокна ячеек не создают и ориентируются циркулярно относительно просвета сосуда. В структуре стенки медии слои соединительной ткани чередуются со слоями гладкой мышечной. Между слоями мышечной ткани имеются прослойки, соединяющие соседние слои. На фоне ячеек соединительной ткани, особенно на фоне гладкой мышечной, легко обнаруживается большое количество кровеносных капилляров.
Адвентиция тонкая от медии отличается иной характеристикой соединительнотканных волокон и отсутствием гладких миоцитов. В ней есть участки, построенные из сочетания коллагеновых и эластических волокон, создающих ячейки разной величины, в которых встречаются кровеносные сосуды. Кровеносные и лимфатические капилляры локализуются близко к медии, а артерии и вены дальше, у ее периферии. Основной объем адвентиции занят толстыми коллагеновыми волокнами, не образующими ячеистых структур.
У наружной оболочки – самая рыхлая структура. Соединительная рыхлая волокнистая ткань адвентиции характеризуется наличием большого количества толстых, слабоокрашен-ных волокон и тонких волокон или пластинок, которые просматриваются в виде серых контрастных теней. Основной объем в ней занимают кровеносные и лимфатические сосуды, расположенные или в средней ее части, или на периферии.
У совы полярной и ястреба тетеревятника внутренняя оболочка аорты складчатая, выстлана эндотелием с высотой эндотелиоцитов, равной их ширине (рис. 2). Кариолемма соседних эндотелиоцитов хорошо заметна, поэтому между соседними клетками имеются полости, простирающиеся до базальной мембраны. Ядра эндотелиоцитов разной формы, одни в виде широких полосок, другие неправильно-овальной формы. Между эндотелием и первым слоем средней оболочки – рыхлые, тонкие, паутинообразные волокна соединительной ткани, формирующие ячеистую структуру. Ядра соединительнотканных клеток – самые плотно окрашенные структуры интимы. Внутренняя эластическая мембрана представлена плотно располагающимися эластическими волокнами.
В средней оболочке широкие слои соединительной ткани чередуются с узкими прослойками гладкой мышечной ткани. В направлении от интимы к адвентиции ширина соединительнотканных прослоек уменьшается, а количество гладкой мышечной ткани увеличивается. Всего насчитывается 16–17 слоев гладких миоцитов и 17–18 – соединительнотканных волокон (рис. 3, 4). Кровеносные капилляры умеренно дифференцируются лишь в средней и наружной частях медии. Между слоями гладкой мышечной ткани – прослойки, по которым гладкая мышечная ткань из одного слоя распространяется в другую. Форма ядер гладких миоцитов имеет косоциркулярное направление.
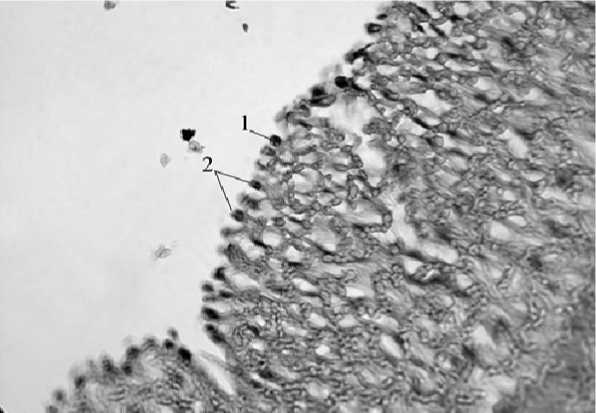
Рис. 2. Столбчатая форма эндотелиоцитов стенки аорты ястреба-тетеревятника (окраска гематоксилин-эозин, ув. х 400); 1 – эндотелиоцит с непрозрачным ядром;
2 – умеренно плотные ядра эндотелиоцитов
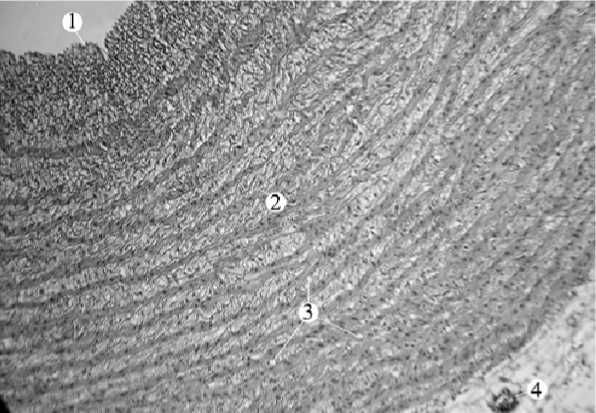
Рис. 3. Стенка аорты ястреба-тетеревятника (окраска гематоксилин-эозин, ув. х 100): 1 – внутренняя оболочка; 2 – средняя оболочка; 3 – капилляры в средней оболочке;
4 – мелкие кровеносные сосуды в наружной оболочке
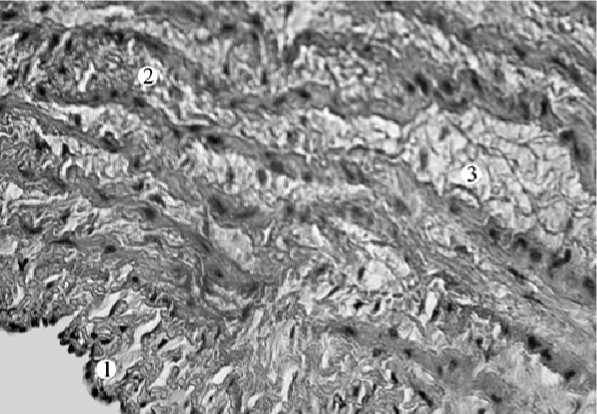
Рис. 4. Стенка аорты совы полярной (окраска гематоксилин-эозин, ув. х 400); 1 – эндотелий; 2 – мышечноэластическая средняя оболочка; 3 – прослойка, соединяющая соседние слои гладкой мышечной ткани
Наружная оболочка рыхлая, представлена крупноячеистой соединительнотканной структурой с базофильным оттенком окраски волокон; часто встречаются мелкие кровеносные сосуды и нервы. Большая часть лимфатических сосудов и вен находится возле медии, а нервы и мелкие артерии от плотной медии ориентированы вдоль сосуда.
Мы согласны с утверждениями [5], что аорта, состоящая из эластических и коллагеновых волокон при прохождении пульсовой волны своим расширением, сужением и скручиванием сосуда оказывает механическое давление на стенки, стимулируя продвижение крови. По нашим данным, преобладание соединительной ткани над гладкой мышечной позволяет аорте при резком взлете птиц отвечать на гидродинамические удары тока крови, предотвращая разрыв стенки, являясь легкорастяжимой при функциональной нагрузке, выполняя роль эластического резервуара и превращая толчки давления, вызываемые систолой левого желудочка, в потенциальную энергию эластического напряжения сосудистых стенок.
Сопоставление морфометрических показателей толщины стенки аорты и ее просвета свидетельствует, что относительно тонкостенной является аорта с наименьшими показателями у ястреба-тетеревятника – 12,89%. Наибольшие показатели отмечаются у совы полярной – 16,46%, у неясыти обыкновенной – 16,56%. Отношение стенки к диаметру ее просвета (индекс Кернога-на) у птиц, использующих маневренный и парящий полет: у ястреба-тетеревятника 1:7,8, у неясыти обыкновенной и совы полярной 1:10,6.
Авторы [6] считают, что в аорте млекопитающих средняя оболочка состоит из трехмерного комплекса – сети гладких мышечных, коллагеновых и эластических волокон. Ориентация и близкая взаимосвязь между волокнами составляет непрерывную большую силу, упругость и способность сопротивления в продольном и циркулярном направлениях. Кроме того, эластические волокна разделяют среднюю оболочку на разное количество концентрически укрепленных волокон. По нашим данным, основную часть стенки аорты составляет средняя оболочка, занимающая от 91,10 (у совы) до 92,10% (у ястреба-тетеревятника) всей толщины стенки.
При анализе морфометрических данных мы отмечаем, что наружная оболочка аорты развита слабо. Ее наименьшие показатели – у неясыти обыкновенной 3,5%, у ястреба-тетеревятника 3,6%, у совы полярной 3,8%, от всей толщины стенки. Результаты наших исследований позволяют согласиться с мнением [7] о том, что наружная оболочка предохраняет аорту от сильных растяжений и разрывов и участвует в регулировании сосудистого тонуса. Под эндотелием волокна соединительной ткани более плотные сливаются в сплошную пластину, формируя внутреннюю эластическую мембрану. У изученных птиц ядра соединительнотканных клеток внутренних слоев медии перпендикулярны к поверхности интимы.
Возможно, столбчатая форма эндотелиоцитов является результатом сокращения площади их эластической мембраны, при которой площадь основания клетки уменьшается, а сохраняющийся объем вытесняется в сторону просвета сосуда, подтверждая прочную связь цитолеммы эндотелиоцитов с внутренней эластической мембраной. Различная форма и плотность окраски ядер эндотелиоцитов объясняется их положением в плоскости среза. Наличие ядер, имеющих форму пластинки, ориентированных под острым углом или почти перпендикулярно к внутренней эластической мембране, свидетельствует о том, что они легко смещаются в цитоплазме эндотелиоцитов. Мы считаем, что эндотелий играет большую роль, являясь барьером с регулирующими свойствами между кровью и основными тканями. Поэтому степень механической прочности последовательно увеличивается в стенке артериального сосуда, в том числе и в аорте от эндотелия к наружной эластической мембране. Учитывая, что у птиц в покое сердечный выброс, как считает [8], в 1,5 раза выше, чем у млекопитающих с такой же массой тела, во время полета эта величина возрастает в несколько раз.
Кровь из аорты поступает в правый и левый плечеголовные стволы. Пульсовая волна из аорты за счет гидростатического и гидродинамического давления оказывает воздействие на стенку правого и левого плечеголовных стволов, которые лежат почти горизонтально в грудной полости и имеют определенные углы расхождения между собой.
Плечеголовной ствол неясыти обыкновенной с толщиной стенки 445,9 ± 0,1 мкм эластического типа. Внутренняя оболочка плечеголовного ствола совы, неясыти на поперечном сечении сосуда имеет выпячивания в сторону просвета сосуда . Ядра эндотелиоцитов располагаются близко друг к другу. Внутренняя и наружная части медии более плотные за счет большого количества прослоек гладкой мышечной ткани, в которой миоциты ориентированы циркулярно или косоциркулярно. Средняя часть медии построена из волокнистой соединительной ткани, поэтому более рыхлая. В ней только одну четвертую часть занимают прослойки гладкой мышечной ткани. Ее наиболее толстые прослойки локализуются в наружной трети медии, а самые тонкие – в средней части, как и у ястреба-тетеревятника и цесарки. Прослоек между слоями гладкой мышечной ткани больше в наружной трети средней оболочки. Во всех слоях медии направление пучков гладких миоцитов, даже в соседних пучках, может быть различным. Всего в медии насчитывается 8–9 гладких мышечных и 10 соединительнотканных слоев.
Интима неоднородная по плотности, длине и конфигурации коллагеновых волокон. Наиболее плотные и длинные волокна локализованы около эндотелия и по мере удаления от него длина и толщина соединительнотканных волокон уменьшается, поэтому около медии адвентиция более рыхлая.
В направлении от адвентиции к интиме количество гладкой мышечной ткани уменьшается, но увеличивается объем соединительной ткани, состоящей из большого количества эластических волокон. Прослойки гладкой мышечной ткани становятся тоньше. Установлено 14–15 гладких мышечных и 15–16 соединительнотканных слоев. На участках, близких к ад-вентиции, гладкие миоциты ориентированы косоциркулярно, а в направлении к интиме увеличивается количество гладких миоцитов, ориентированных вдоль просвета сосуда.
Гладкая мышечная ткань, находящаяся близко к интиме, создает волнистый рисунок. Возле адвентиции гладкая мышечная ткань имеет ровный, однородный фон окраски цитоплазмы.
В средней оболочке отмечают кровеносные капилляры и мелкие лимфатические сосуды, которые локализуются по всей толщине медии и на всех поперечных срезах стенки сосуда имеют щелевидную или неправильно удлиненную форму в связи с продольной ориентацией капилляров.
Для наружной оболочки плече-головного ствола характерны неплотные соединительнотканные волокна, ориентированные в разных направлениях. В адвентиции локализуется большое количество артериальных и венозных капилляров.
Плечеголовной ствол ястреба-тетеревятника эластического типа с толщиной стенки 465,3 ± 0,3 мкм. Интима тонкая, окрашивается слабее, чем адвентиция, но интенсивнее, чем медиа. Внутренняя часть медии начинается короткими, отдельными фрагментами гладкой мышечной ткани, разделенными широкими прослойками соединительной, количество которой увеличивается от интимы к адвентиции. В средней части медии пучки гладких миоцитов имеют вид длинных, тонких прослоек. Между ними располагаются широкие слои соединительной ткани, создавая больший, чем у мышечной, объем соединительной ткани. Для на-ружнай трети медии характерно наличие узких прослоек соединительной ткани, а основной объем составляет гладкая мышечная ткань. Насчитывают 14–15 гладких мышечных и 15–16 соединительнотканных слоев. На всех участках медии пучки гладких миоцитов ориентированы относительно просвета сосуда косоциркулярно, формируя спиралевидную структуру гладкой мышечной ткани. Отмечено расположение витков спирали с взаимоперпендикулярным направлением в левую и правую стороны.
Адвентиция развита слабо, представлена соединительнотканными волокнами.
При морфометрическом анализе плечеголовных стволов мы отмечаем, что по отношению к просвету сосуда внутренняя оболочка более тонкая и составляет у ястреба-тетеревятника 15,31%. Средние показатели внутренней оболочки у неясыти обыкновенной – 17,15%. Наиболее толстая оболочка – у совы полярной 18,1%. Индекс Керногана стенки плечеголовного ствола у неясыти обыкновенной – 1:5,8 и у ястреба-тетеревятника и совы полярной – 1:6,5 и 1,66. Средняя оболочка плечеголовных стволов – толстая у птиц, использующих парящий, маневренный полет, зафиксированы показатели у неясыти обыкновенной и совы полярной 89,28% и у ястреба-тетеревятника – 91,95%.
Внутренняя оболочка плечеголовного ствола тонкая, занимает от 4,2 (у ястреба-тетеревятника) до 5,1% (у совы полярной) толщины стенки сосуда. Средняя оболочка – самая толстая в стенке плечеголовных стволов. Данные морфометрического анализа показывают, что в отношении средней оболочки к толщине стенки сосуда у птиц, использующих парящий маневренный полет, наименьшие показатели – у неясыти обыкновенной – 89,28%, наибольшие – у ястреба-тетеревятника – 91,95%.
Пучки гладких миоцитов в виде тонких, длинных слоев имеют больший объем в средней оболочке. В наружной части средней оболочки отмечено наличие узких прослоек соединительной ткани; основной объем составляют гладкие миоциты. У неясыти и ястреба-тетеревятника 14–15 гладких миоцитов. Что касается соединительнотканных слоев, то у неясыти и ястреба-тетеревятника их 15–16.
Заключение
В результате проведенных исследований нами установлено, что аорта и плечеголовные артерии у неясыти обыкновенной, совы полярной и ястреба-тетеревятника сохраняют общий план строения сосудов, характерный для млекопитающих. Относятся к сосудам эластического типа строения. Средняя оболочка состоит из трехмерного комплекса – гладких мышечных, коллагеновых и эластических волокон. Ориентация и близкая взаимосвязь между этими волокнами составляет непрерывную большую силу, упругость и способность к сопротивлению в продольном и циркулярном направлениях. Кроме того, эластические волокна разделяют среднюю оболочку на разное количество концентрически укрепленных волокон, обеспечивающих им высокую механическую прочность. Чередование прослоек гладких миоцитов и соединительнотканных мембран подтверждает различную сократительную способность слоев.
Список литературы Видовые особенности гистологического строения аорты у неясыти обыкновенной, совы полярной и ястреба-тетервятника
- Папченко, И.В. Некоторые особенности гистологического строения артерий грудной конечности кур/И.В. Папченко//Эколого-экспериментальные аспекты функциональной и возрастной морфологии домашних птиц. -Воронеж, 1988. -С. 24 -27.
- Salomon, V.F. Lehrbuch der Geflugelanatomie/V.F. Salomon. -Sttudgart, 1993. -S. 100 -158, 290 -300.
- Konig, H.E. Anatomia der Vogel/H.E. Konig., R. Korbel, H.-G. Liebich. -Sthatteauer GmbH, 2008. -S. 64 -88, 78 -92, 184 -200.
- Меркулов, Г.А. Курс патологической техники: практ. пособие/Г.А. Меркулов. -Л.: Медгиз, 1969. -424 с.
- Holzapeel, G.A. A new constitutive frame work for arterial wall mechanics and a comparative study of material model/G.A. Holzapeel, T.C. Gasser//Journal of Elasticity. -2000. -V. 61. -Р. 1 -48
- Wagenseil, J.E. Vascular extracellular matrix and arterial mechanics/J.E. Wagenseil, R.P. Mecham//Physiol. Rev. -2009. -V. 89. -P. 957 -989.
- Lawrence, A.R. Transmural pressur and axial loading intraactively regulare arterial remodeling ex vivo/A.R. Lawrence, K. J. Gooch//Am. J. of Physiology. -2009. -V. 297, № 1 -P. 475 -484.
- Rayner, J.M. Wake structure and forse degeneration in avian flapping flight/J.M. Rayner//Acta 20 Cong. Jnt. Ornithol., Christchurch, 2-10 Dec., 1990. -Wellington, 1991. -V. 2. -P. 702 -715.
