Вирулентность заразихи, поражающей подсолнечник, в Волгоградской и Ростовской областях

Автор: Антонова Т.С., Арасланова Н.М., Рамазанова С.А., Гучетль С.З., Челюстникова Т.А.
Статья в выпуске: 1 (146-147), 2011 года.
Бесплатный доступ
В Волгоградской и Ростовской областях на подсолнечнике выявлены расы заразихи E, F, G, H. При этом образцы семян растения-паразита из Волгоградской области показали значительно меньшую вирулентность, чем из Ростовской. Наблюдается пестрота по вирулентности заразихи в разных районах изученных регионов. Заразиха с разных полей одного и того же района также может значительно различаться по вирулентности, что имеет значение при размещении сортимента подсолнечника, отличающегося генами устойчивости. Это также следует учитывать и при селекции подсолнечника на устойчивость.
Подсолнечник, заразиха, расы, вирулентность, дифференциаторы, районы
Короткий адрес: https://sciup.org/142150973
IDR: 142150973 | УДК: 632.9:633.854.78
Virulence of broomrape affecting sunflower in Volgograd and Rostov regions
Broomrape races E, F, G, H were found out on sunflower in Volgograd and Rostov regions. Sample of broomrape seeds from Volgograd region were less virulent in comparison with the same one from Rostov region. There is diversity on broomrape virulence in different areas of investigated regions. The broomrape samples from the same area also can differ significantly by its virulence. It is important for placement of sunflower assortment which is differed with gens of resistance. It is also meaningful for sunflower breeding on resistance.
Текст научной статьи Вирулентность заразихи, поражающей подсолнечник, в Волгоградской и Ростовской областях
Введение. Облигатный цветковый паразит из высших растений – заразиха, Orobanche cu-mana Wallr., паразитируя на корнях подсолнечника, способна не только значительно снизить урожай семян, но и уничтожить саму культуру в целом, сделав её нерентабельной. Селекция подсолнечника на устойчивость к заразихе ведётся постоянно на протяжении почти столетия, так как в ходе сопряжённой эволюции с подсолнечником у растения-паразита периодически возникают новые вирулентные расы, преодолевающие иммунитет существующего сортимента.
На подсолнечнике известно 8 рас O. сuma-na, названных буквами латинского алфавита: A, B, C, D, E, F, G, H. Три последние из них – самые вирулентные, на протяжении последнего десятилетия были обнаружены сначала в Румынии, затем в Испании и Турции [1-5]. В этих странах были созданы дифференциаторы устойчивости подсолнечника к каждой из них.
В последние годы в регионах юга РФ, где возделывается подсолнечник, тоже начали распространяться новые расы заразихи, которые стали поражать весь отечественный сортимент и гибриды иностранной селекции. Актуальными стали все аспекты борьбы с этим злостным паразитом.
Как показали наши недавние исследования [6], особенно высоковирулентной проявила себя заразиха в ряде районов Ростовской области, в том числе, расположенных близко к Вол- гоградскому региону. Одной из основных причин ускорения формирования высоковирулентных биотипов заразихи именно в Ростовской области является интенсивное воз-делы-вание подсолнечника в этом регионе. Культуру из-за её высокой доходности стали возвращать на прежнее поле через 1-3 года, тогда как научно обоснованный севооборот для подсолнечника составляет 8 лет.
В районах Волгоградской области устойчивый ранее сортимент подсолнечника также начал поражаться, местами довольно сильно, в частности, иностранные гибриды, заявленные как устойчивые к расам Е и F.
Целью наших исследований было определение расовой принадлежности образцов семян заразихи, собранных в разных районах Волгоградской и Ростовской областей.
Материалы и методы . Образцы семян заразихи были собраны на отдельных полях в 6 районах Волгоградской области: Суровикин-ском, пригороде Волгограда, Новоаннинском, Киквидзенском, Еланском, Руднянском, в Ростовской: Азовском, Аксайском, Белокалитвин-ском, Морозовском, Обливском, Тацинском. Для идентификации их расовой принадлежности использовали линии-дифференциаторы подсолнечника: LC1002, LC1003, ВК 623, LC1093, Р 96, и две гибридных комбинации 16×25 и ВТ 62. Идентификацию проводили на центральной экспериментальной базе ВНИИМК.
Растения подсолнечника выращивали в тепличных условиях в цветочных ящиках, заполненных почвенно-песчаной смесью, смешанной с семенами заразихи каждого образца отдельно. Семена заразихи вносили из расчёта 200 мг на 1 кг почвы. Через 30 дней после появления всходов растения подсолнечника выкапывали, отмывали корни водой и подсчитывали на них клубеньки и проростки заразихи (рис. 1).
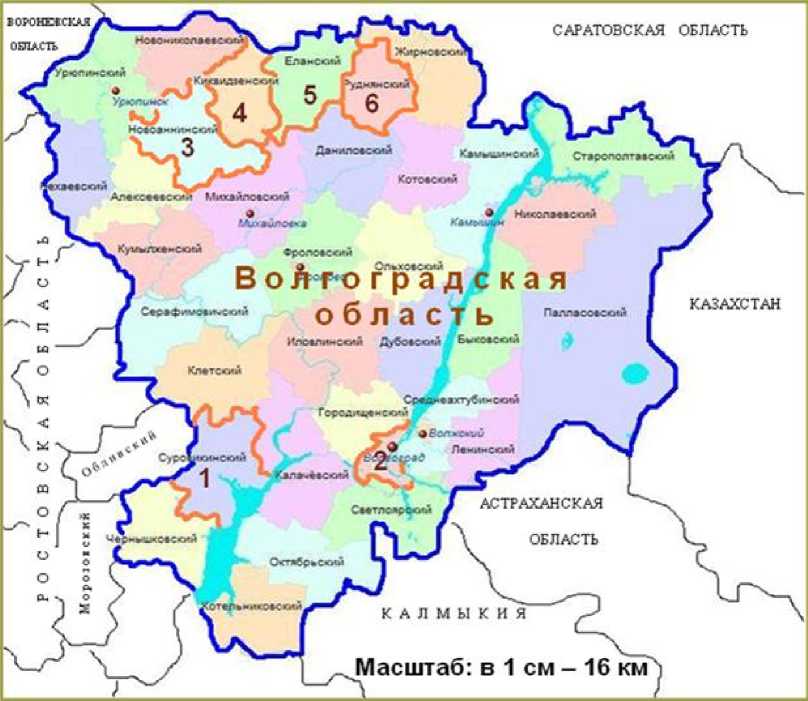
Результаты и обсуждение . Районы Волгоградской области, где были собраны семена заразихи, располагаются в южной и северной части региона (рис. 2). Карту районов Ростовской области здесь не приводим. Она была показана ранее [6]. Как видно из последнего столбца таблицы 1, гибридная комбинация, имеющая устойчивость к расе Н, практически не поразилась заразихой из Волгоградской области, то есть отсутствуют биотипы паразита с

Рисунок 1 – Поражение заразихой из Новоаннинского района Волгоградской области дифференциаторов подсолнечника с устойчивостью к расам: 1 – D; 2 – E; 3 – F; 4 – G
10МІНЕЮУШ
АСТРАХАНСКАЯ
ОБЛАСТЬ
Масштаб: в 1 см - 16 км
САРАТОВСКАЯ ОБЛАСТЬ
ОЕЛАСТЬ
ФрО.АЖ<иЙ
В а Оо»хоес<*1
о frroградская
КАЗАХСТАН область
Кпоелі**со«<* Дубо*с<««и
КлфтсммА
Рисунок 2 – Карта Волгоградской области с указанными районами, где были собраны семена заразихи, паразитирующей на подсолнечнике: 1 – Суровикинский 2 – пригород Волгограда, 3 –
Новоаннинский, 4 – Киквидзенский, 5 – Еланский, 6 – Руднянский
Таблица1 – Степень поражения* заразихой с отдельных полей из некоторых районов Волгоградской и Ростовской областей дифференциаторов подсолнечника
|
Район |
Ди |
ференциаторы устойчивости к расам заразихи: |
|||||
|
LC1002 D** |
LC1003 E |
BK 623 E |
LC1093 F |
P 96 F |
16× 25 G |
ВТ 62 H |
|
|
Волгоградская область |
|||||||
|
Суровикинский |
84 |
10 |
0 |
0 |
1 |
3 |
0 |
|
Пригород Волгограда |
24 |
20 |
5 |
7 |
2 |
11 |
0 |
|
Новоаннинский |
115 |
28 |
57 |
12 |
2 |
0 |
0 |
|
Киквидзенский |
67 |
24 |
8 |
17 |
2 |
1 |
0 |
|
Еланский |
78 |
72 |
46 |
25 |
6 |
1 |
1 |
|
Руднянский |
23 |
11 |
5 |
47 |
1 |
0 |
0 |
|
Ростовская область |
|||||||
|
Азовский |
42 |
18 |
32 |
38 |
9 |
3 |
0 |
|
Аксайский |
57 |
35 |
22 |
36 |
16 |
0 |
0 |
|
Белокалитвинский |
52 |
104 |
73 |
68 |
37 |
36 |
1 |
|
Морозовский |
71 |
69 |
80 |
62 |
28 |
4 |
3 |
|
Обливский |
83 |
67 |
30 |
26 |
10 |
2 |
1 |
|
Тацинский (поле 1) |
56 |
47 |
78 |
55 |
19 |
5 |
0 |
|
Тацинский (поле 2) |
30 |
35 |
43 |
21 |
12 |
35 |
1 |
|
* Степень поражения – количество клубеньков и проростков заразихи на одно поражённое растение подсолнечника **Наименование расы, к которой устойчив дифференциатор |
|||||||
вирулентностью выше, чем у расы Н. Заразиха же из трёх районов Ростовской области дала единичные поражения. Их можно расценивать двояко: как наличие единичных особей паразита, преодолевающих иммунитет этого гибрида, и как следствие его генетической неоднородности (примесь семян, восприимчивых к расе Н). Устойчивость гибридной комбинации к расе G преодолевается заразихой из пригорода Волгограда со степенью 11, что свидетельствует о наличии там расы Н. В трёх других районах встречаемость единичная. В то же время в популяциях заразихи из двух районов Ростовской области – Белокалитвинского и Тацин-ского – высокая концентрация этой расы (степень поражения 36 и 35 соответственно).
Линия Р 96 с совместным действием двух рецессивных генов устойчивости or6or7 к расе F почти не поражается заразихой из районов Волгоградской области, но имеет значимую степень поражения в представленных в таблице районах Ростовской. Линия LC1093 с доминантным геном Or6 устойчивости к расе F имеет высокую степень поражения заразихой из Руднянского и Еланского районов, степень поражения соответственно 47 и 25. Устойчивость, обусловленная этим геном, преодолевается и с несколько меньшей степенью заразихой из районов Киквидзенского, Новоаннинского и пригорода Волгограда. Однако степень поражения этой линии заразихой из всех районов Ростовской области значительно более высокая. Поражение обеих линий с разными генами устойчивости к расе F свидетельствует о наличии во всех районах, кроме Суро-викинского, расы G.
Заразиха из районов Новоаннинского и Еланского так же, как из всех районов Ростовской области, выделяется высокой концентрацией расы F, так как значительно поражены обе линии ВК 623 и LC1003, устойчивые к расе Е, благодаря наличию у них гена Or5. Высокая степень поражения линии LC1002 с геном устойчивости Or4 свидетельствует о наличии во всех районах расы Е. Образец семян заразихи из Суровикинско-го района мало вирулентен и представлен преимущественно расой Е. В образце из Ново аннинского района в основном смесь рас Е и F.
В таблице 1 семена заразихи из Тацинского района представлены с двух полей. Поле 2 выделяется высокой концентрацией расы Н. Поле 1 представлено преимущественно расами F и G. Это существенные различия, имеющие значение для размещения сортимента с разной устойчивостью к заразихе. Кроме того, эти данные показывают, что при использовании семян заразихи для селекционных целей необходимо предварительно проверять степень их вирулентности, даже если они собраны в одном районе.
Таким образом, в Волгоградской и Ростовской областях на подсолнечнике выявлены расы заразихи E, F, G, H. При этом образцы семян растения-паразита из Волгоградской области показали значительно меньшую вирулентность, чем из Ростовской. Наблюдается пестрота по вирулентности заразихи в разных районах изученных регионов. Заразиха с разных полей одного и того же района также может значительно различаться по вирулентности, что имеет значение при размещении сортимента подсолнечника, отличающегося генами устойчивости. Это также следует учитывать и при селекции подсолнечника на устойчивость.
