Влияние аэротехногенных загрязнений на состав сообществ эпифитных микромицетов г. Уфы

Автор: Дубовик И.Е., Шарипова М.Ю., Климина И.П.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Впервые проведено изучение таксономической структуры и закономерностей развития эпифитных микромицетов под влиянием промышленного загрязнения. В качестве объектов для стационарного исследования были выбраны пять видов древесных растений: хвойные (Picea obovata Ledeb.) и лиственные (Betulapendula Roth, Populus nigra L., Tilia cordata Mill., Sorbus aucuparia L.). Составлен таксономический список, включающий 41 вид микроскопических грибов коры древесных растений, которые в зоне нефтехимического комплекса были представлены 23 видами, а в контрольной зоне города - 28. Доминирующее положение по числу видов на коре деревьев занимают представители рода Penicillium. В фоновой зоне отмечалась большая встречаемость представителей рода Mucor, составляющая 67% общего числа эпифитных микромицетов. Выявлена специфика организации комплекса микромицетов в зависимости от условий обитания, показано, что полученные результаты могу быть использованы для биомониторинга.
Аэротехногенное загрязнение, эпифитные микромицеты, биомониторинг
Короткий адрес: https://sciup.org/147227009
IDR: 147227009 | УДК: 582.28
The impact of aerial technogenic pollution on the epiphytic community composition of the of micromycetes of the city of Ufa
The study of the taxonomic structure and patterns of development of epiphytic micromycetes under the influence of industrial pollutionwas madefor the first time. Five species of woody plants were chosen as objects for stationary research: coniferous (Picea obovata Ledeb.) And deciduous (Betula pendula Roth, Populus nigra L., Tilia cordata Mill., Sorbus aucuparia L.). A taxonomic list has been compiled that includes 41 species of microscopic fungi of bark of woody plants:in the zone of the petrochemical complexthey were represented by 23 species, and in the control zone of the city - 28.The dominant position in the number of species on the tree bark is occupied by representatives of the genus Penicillium..In the kontrolzone, a large occurrence of representatives of the genus Mucor, accounting for 67% of the total number of epiphytic micromyceteswas noted.The specifics of the organization of the micromycete complex depending on the habitat conditions are revealed, it is established that the obtained results can be used for biomonitoring.
Текст научной статьи Влияние аэротехногенных загрязнений на состав сообществ эпифитных микромицетов г. Уфы
Биоиндикация урбанизированных экосистем традиционно находится в центре эколого-ботанических исследований. Индикаторами техногенного загрязнения служат как высшие, так и низшие растения. Работы, в которых исследовался грибной компонент техногенных экосистем, касаются, в основном, узкоспециализированных групп микоризообразователей [Станкявичене, Урбонас, 2006] или ксилотрофных грибов (как классические примеры растительно- грибного взаимодействия). Рекреация и загрязняющие вещества оказывают влияние на такие показатели, как видовое богатство, доминирование, деструктивная активность ксилотрофных грибов [Волчатова и др., 2007; Сафонов, Сафонова, 2009].
В то же время большой интерес представляет обширная и менее специализированная группа микроскопических грибов, не относящихся к кси-лотрофам, но обитающих на древесной коре и наряду с водорослями и цианобактериями образующих макроскопические разрастания. Ранее были
получены данные, что фикобионты не являются индикаторами воздушного загрязнения [Дубовик, Шарипова, 2016], однако данные о микобионтах отсутствуют. В связи с этим были поставлены следующие задачи:
-
- определить видовой состав микроскопических грибов коры древесных растений, их приуроченность к разным форофитам ;
-
- сравнить микобиоту контрольной и промышленной зон на территории г. Уфы;
-
- изучить особенности сезонной динамики видового состава комплекса микромицетов;
-
- провести сравнительные исследования влияния промышленного загрязнения на комплекс микромицетов коры древесных растений, выявить возможность их использования для биоиндикации, дать оценку экологически опасным последствиям
трансформации комплексов микроскопических грибов.
Материал и методы исследования
В качестве объектов для стационарного исследования были выбраны пять видов древесных растений: хвойные (ель сибирская – Рicea obovata Le-deb.) и лиственные (береза повислая – Betula pen-dula Roth, тополь черный – Populus nigra L., липа сердцелистная – Tilia cordata Mill., рябина обыкновенная – Sorbus aucuparia L.). Выбор этих фо-рофитов обусловлен их широким распространением на данных территориях. Исследования проводились в рекреационной (фоновой) зоне города (контроль) и промышленной (территория нефтеперерабатывающего комплекса «Башнефть», расположенного в северной части г. Уфы) (табл. 1).
Таблица 1
Характеристика соcтояния воздушного бассейна по основным загрязнителям в изученный период
|
Наименование примеси |
Промышленная зона, мг/м3 |
Зона контроля, мг/м3 |
Значение ПДКс.с., мг/м3 |
|
Взвешенные вещества |
0.15 |
0.06 |
0.15 |
|
Диоксид серы |
0.005 |
0.002 |
0.05 |
|
Оксид углерода |
2.0 |
1.4 |
3.0 |
|
Диоксид азота |
0.04 |
0.02 |
0.04 |
|
Хлорид водорода |
0.04 |
- |
0.1 |
|
Формальдегид |
0.006 |
- |
0.003 |
|
Бензол |
0.02 |
- |
0.1 |
|
Четырёххлористый углерод |
0.01 |
- |
0.7 |
|
Хлороформ |
0.012 |
- |
0.03 |
|
Бенз(а)пирен |
2.8 |
- |
1.0 |
Образцы коры деревьев (всего 316) отбирали на высоте 150–155 см от поверхности почвы. Каждый образец являлся средней пробой, полученной при смешивании 10 индивидуальных с одного вида древесных растений в разные сезоны. Образцы коры размером приблизительно 5 x 5 см и толщиной 2–5 мм. При сборе проб и подготовке их к анализу соблюдали необходимые условия стерильности.
Выделение и количественный учет микромице-тов проводили по общепринятой методике посева суспензии измельчённой коры на агаризованную подкисленную среду Чапека ( в трех повторностях). Состав среды: сахароза – 30.0 г; NaNO 3 – 2.0; K 2 HPO 4 – 1.0; MgSO 4* 7H 2 O – 0.5; KCl – 0.5; FeSO 4* 7H 2 O – 0.1; агар – 20.0г; вода дистиллированная – 1000 мл. Для роста грибов среду подкисляли (0.5% молочной кислоты). Инкубирование проводили при температуре 27–28оС. Учет микро-мицетов осуществляли на седьмые сутки [Методы …, 1991]. Для определения патогенности выделенных нами видов, использовали классификацию С. Де Хога [Марфенина, 2005].
Идентификацию видов микромицетов проводили по определителям [Милько, 1974; Билай, Коваль, 1988; Билай, Курбацкая, 1990; de Hoog et al., 2000; Саттон, Фотергилл, Ринальди, 2001]. Видо- вые названия грибов уточняли по пополняемым спискам опубликованных видов в базе данных «Index fungorum» (.
Учет мицелия грибов проводили методом мембранных фильтров в модификации Демкиной и Мирчинк. Использовали фильтры марки «Сын-пор» с диаметром пор 2.5 мкм или диаметром 35 с порами 2.5 мкм и краситель дианиловый голубой. Суммарную длину гиф мицелия в 1 г почвы рассчитывали по формуле b x104 5и102 a =-----------s-,
50 PVC 108
где а – длина гиф в 1 г почвы (см); b – суммарная длина гиф на фильтре в 50 полях зрения в единицах окуляр-микрометра (мкм); S – площадь фильтра (мм2); n – разведение почвенной суспензии; P – площадь зрения (мкм2); V – объем наносимой суспензии; С – навеска почвы, коры (г).
Для установления типичных и доминантных видов определяли обилие (плотность) вида – отношение числа штаммов данного вида к общему числу всех видов. Доминантность вида определяется по его обилию (абсолютные доминанты – свыше 50% от всех имеющихся форм; доминанты первого ранга от 25 до 50%; доминанты второго ранга от 5 до 25% от всех имеющихся форм) [Мирчинк, 1988].
Для определения значимости вида применяли критерий частоты встречаемости по показателю временной частоты встречаемости вида, устанавливаемый как отношение числа моментов времени, когда вид обнаружен, к общему числу моментов отбора образцов.
При совместном использовании показателей пространственной и временной частот встречаемости Т.Г. Мирчинк [1988] предложено характеризовать структуру почвенных микромицетов следующим образом: типичные доминирующие виды – пространственная и временная частота встречаемости выше 60%; типичные частые – пространственная и временная частота встречаемости более 30%; типичные редкие – пространственная частота встречаемости ниже 30% и временная выше 30% и случайные виды – оба показателя ниже 30%. При анализе особенностей микобиоты использовался коэффициент сходства Сё-ренсена [Одум, 1986].
Статистическую обработку результатов производили с применением пакета прикладных программ Statistica V 6.0.
Результаты и их обсуждение
Был выявлен 41 видовой таксон эпифитных микромицетов, которые в зоне нефтехимического комплекса были представлены 23 видами, а в контрольной зоне города – 28. Доминирующее положение по числу видов на коре деревьев занимают представители рода Penicillium, что составляет 61% от всех выделенных видов. В промышленной зоне этот род представлен 10 видами, а в зоне контроля – 18. В фоновой зоне большая встречаемость представителей рода Mucor, составляющая 67% от общего числа эпифитных микромицетов. Представителей данного рода в промзоне обнаружено не было. Г.А.Евдокимовой, М.В. Корнейковой, Е.В. Лебедевой [2007] также была отмечена чувствительность представителей порядка Mucorales к выбросам алюминиевого предприятия. Было показано, что сопутствующая микромицетам альгофлора не зависит от аэротехногенного загрязнения [Дубовик, Климина, 2010; Дубовик, Киреева, Климина, 2012; Дубовик, Шарипова, 2016].
В наших исследованиях выявлен рост доли грибов родов Aspergillus и Trichoderma относительно микромицетов рода Penicillium и появление меланинсодержащих грибов в зоне возле промышленных предприятий. Подобные изменения наблюдали в комплексах микромицетов городских почв [Марфенина, 1999] что, возможно, указывает на единый механизм воздействия антропогенных факторов на микроскопические грибы почвенного и надпочвенного ярусов городской экосистемы. О.Е. Марфенина [1989] отмечала интенсивное развитие грибов данного рода при высоком уровне загрязнения тяжёлыми металлами. Кроме того, в зоне промышленного загрязнения отмечена большая частота встречаемости ряда видов микромицетов из рода Aspergillus .
Анализ частых, редких и случайных видов также показал качественные различия. На территории нефтеперерабатывающего предприятия доминантные формы грибов практически сохраняются, но происходит обеднение видовой структуры комплексов микромицетов за счёт выпадения числа редких видов (табл. 2).
Таблица 2
Частота встречаемости микроскопических грибов на коре древесных растений в промышленной (П) и контрольной (К) зонах
|
№ |
Таксон |
П |
К |
Уровень патогенности |
|
Класс Zygomycetes |
||||
|
1 |
Mucor sp. |
- |
Д |
|
|
2 |
Rhizopus microsporus Tiegh. |
С |
- |
|
|
Класс Hyphomycetes |
||||
|
3 |
*** Alternaria alternata (Fr.) Keissl. |
Д |
ТЧ |
BSL-2 |
|
4 |
*** Aspergillus candidus Link |
С |
- |
|
|
5 |
*** A. flavusvar.orysae (Ahlb.) Kurtz. |
ТЧ |
- |
|
|
6 |
*** A. fumigates Fresen. |
Д |
Д |
BSL-2 |
|
7 |
A. fumigatus var. griseibrunneus Rai et Singh |
ТЧ |
- |
BSL-2 |
|
8 |
*** A. nigervar.niger Tiegh. |
Д |
Д |
BSL-2 |
|
9 |
A. sulphureus Desm. |
- |
С |
|
|
10 |
** A. terreus Thom |
С |
- |
BSL-2 |
|
11 |
*** Aureobasidium pullulans (de Bary) G. Arnaud |
ТР |
- |
BSL-2 |
|
12 |
Isaria farinosa (Holmsk.) Fr. |
- |
С |
|
|
13 |
Penicillium albocinerascens Chalab. |
ТЧ |
С |
|
|
Окончание табл. 2 |
||||
|
№ |
Таксон |
П |
К |
Уровень патогенности |
|
14 |
P. aculeatum Raper&Fennell |
- |
С |
|
|
15 |
P. atrovenetum G. Sm. |
- |
C |
|
|
16 |
* P. canescens Sopp |
ТЧ |
- |
|
|
17 |
*** P. chrysogenum Thom |
С |
- |
BSL-1 |
|
18 |
*** P. citrinum Sopp |
С |
Д |
BSL-2 |
|
19 |
P. corylophiloides S.Abe |
- |
С |
|
|
20 |
P. cyaneum (Bainier&Sartory) Biourge ex Thom |
- |
С |
|
|
21 |
P. duclauxii Delacr . |
- |
ТЧ |
|
|
22 |
* P. funiculosum Thom |
- |
ТЧ |
BSL-2 |
|
23 |
*** P. glabrum (Wehmer) Westling |
С |
- |
|
|
24 |
P. implicatum Biourge |
- |
С |
|
|
25 |
* P. lanosum Westling |
ТЧ |
Д |
|
|
26 |
P. melinii Thom |
- |
С |
|
|
27 |
P. nalgiovense Laxa |
- |
ТР |
|
|
28 |
P. ochraceum Corda |
ТР |
- |
|
|
29 |
P. purpurescens (Sopp) Biourge |
- |
C |
BSL-2 |
|
30 |
*** P. purpurogenum Fleroff |
- |
ТЧ |
|
|
31 |
P. resticulosum Birk., Raistrick& G. Sm. |
- |
ТР |
|
|
32 |
* P. simplicissimum (Oudem.) Thom |
С |
- |
|
|
33 |
P. solitum var. crustosum (Thom) Bridge |
ТЧ |
- |
|
|
34 |
P. solitum var. solitum Westling. |
- |
C |
|
|
35 |
* P. velutinum J.F.H. Beyma |
- |
ТЧ |
6 |
|
36 |
* P. variabile G. Mey. |
С |
Д |
BSL-2 |
|
37 |
Trichoderma citrinoviride Bissett |
- |
ТР |
|
|
38 |
** T. harzianum Rifai |
ТЧ |
ТР |
|
|
39 |
*** T. koningii Oudem. |
С |
С |
BSL-1 |
|
40 |
** T. viride Pers. |
Д |
ТЧ |
BSL-2 |
|
Класс Coelomycetes |
||||
|
41 \ |
** Phoma glomerata (Corda) Wollenw. & Hochapfel |
С |
- |
|
|
ВСЕГО |
23 |
28 |
||
Примечание: Д – доминантные; ТЧ – типичные частые; ТР – типичные редкие; С –случайные; * – фитотоксич-ный вид; ** – оппортунистический вид; *** – условно патогенные.
Только для загрязнённой территории характерны Rhizopus microsporus, Aspergillus candidus, A. fumigates var. griseibrunneus, A. terreus, Aureo-basidium pullulans, Penicillium canescens, P. ochraceum, P. chrysogenum, P. simplicissimum, P. solitumvar. crustosum, P. glabrum, Phoma glomerata. Микромицетами, обнаруженными только в контрольной зоне, были: Mucor sp., Aspergillus sulphureus, Isaria farinosa, Penicillium aculea-tum, P. purpurescens, P. duclauxii, P. corylo-philoides, P. cyaneum, P. melinii, P. funiculosum, P. implicatum, P. nalgiovense, P. purpurogenum, P. ve-lutinum, P. resticulosum, P. solitum var. solitum, Trichoderma citrinoviride.
Сравнение частоты встречаемости сходных видов в сообществах эпифитных микромицетов загрязнённой и контрольной зон показало неоднозначный результат. Так, на двух сравниваемых участках виды Aspergillus fumigatus, A. niger var. niger являются доминирующими, они встречались практически во всех образцах коры древесных растений. Penicillium albocinerascens, входивший в группу случайных видов на незагрязненной территории, становился частым при загрязнении. Напротив, виды Penicillium citrinum, P. variabile оказались чувствительными к аэротехногенному загрязнению. При сравнении комплексов микроми-цетов промышленной и контрольной зон коэффициент Сёренсена составил 38%.
На коре древесных растений промышленной зоны численность микроскопических грибов была несколько выше, чем в контрольной (рис. 1). Так как развитие колоний грибов происходит преимущественно из спор, то увеличение количества мик-ромицетов, возможно, связано с усилением споруляции при аэротехногенном загрязнении. Очевидно, спорообразование способствует выживанию микробиоты в неблагоприятных условиях.
Измерение длины мицелия показало колебание данной величины в пределах от 0.26 до 1.64 м/г и зависимость как от места отбора пробы, так и от вида форофита, кора которого была использована для микологического анализа (рис. 2). Промышленное загрязнение нефтеперерабатывающего комплекса оказывало влияние на длину гифов, приводя к увеличению их длины в 2–5 раз. Тенденция к увеличению количества мицелия на стволах деревьев четко прослеживалась для таких пород, как береза (в 4.9 раз), тополь (в 3.8 раз). Поскольку длина мицелия является степенью активности грибов [Домр показали, что в промыщленной зоне складываются благоприятные условия для развития эпифитных микромицетов на коре всех исследованных форо-фитов. В то же время на коре ели длина грибных гиф в промышленной и контрольной зонах ниже, чем на листопадных деревьях. Возможно, наблюдаемые различия связаны с наличием в коре хвойных пород биологически активных веществ, способных влиять на рост и размножение микроорганизмов [Смирнов и др., 2002].
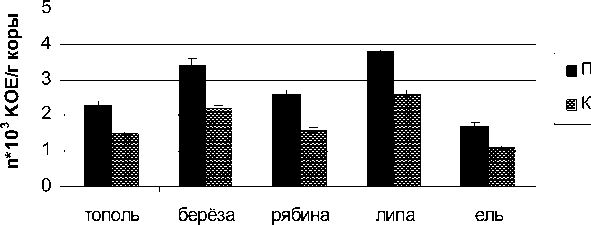
Рис. 1 . Численность микроскопических грибов на коре древесных растений, КОЕ х 10 3 /г
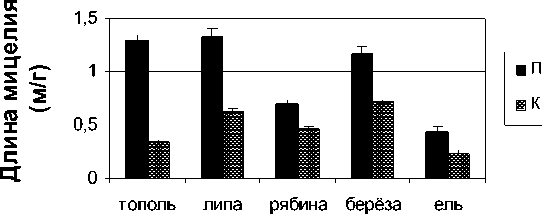
Рис. 2 . Средняя длина гиф (м/г) микромицетов, выделенных с коры деревьев, произрастающих в промышленной и контрольной зонах
Анализ видового состава микроскопических грибов показал наличие на коре деревьев оппортунистических и фитопатогенных видов. Были обнаружены условно патогенные виды грибов, относящиеся к родам Alternariа, Aspergillus, Aureo-basidium, Penicillium, Trichoderma, Phoma. Представители этих родов являются возбудителями заболеваний органов дыхания, кожных покровов, а также способствуют развитию аллергических реакций. На территории промзоны, по сравнению с контрольной, выявлено больше условно патогенных микромицетов, доля которых в контрольной зоне составляла 29%, а в зоне промышленного загрязнения – 57% от общего числа видов (рис. 3). Оппортунистические микозы человека, вызываемые условно патогенными грибами, представляют собой обширные поражения грибным мицелием слизистых оболочек или легких (реже других внутренних органов) у пациентов, страдающих иммунодефицитными состояниями или в послеоперационный период [Марфенина, 2005]. Отмечены изменения в структуре комплексов условно патогенных видов грибов на коре деревьев в фоновой и загрязнённой зонах, а именно: только в составе комплекса грибов загрязнённой зоны выявлены условно патогенные виды: Aspergillus candidus, A. flavus var.orysae, A. terreus, Aureo-basidium pullulans, Penicillium chrysogenum, P. gla-brum, Phoma glomerata.
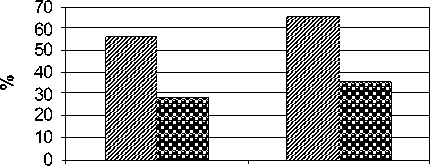
оппортунистические фитопатогенные промы шленная зона контрольная зона
Рис. 3 . Соотношение фитопатогенных и оппортунистических видов микромицетов на коре древесных растений
Наибольшим обилием в исследуемых зонах среди других аэрофильных оппортунистических видов мик-ромицетов отличились Alternaria alternata, Aspergillus fumigatus, A. niger . Данная группа микромицетов представлена эврибионтными видами, выделяемыми из различных местообитаний; они часто обнаруживались в деградированных и химически загрязненных почвах [Терехова, 2007]. Также О.Е. Марфенина, Г.М. Фомичёва [2007] указывали на эвритопность таких потенциально патогенных микромицетов, как Al-ternaria alternata, Aspergillus niger . Все три вида могут выступать в качестве аллергенов; кроме того, Aspergillus fumigatus может вызывать микозы у больных с ослабленной иммунной системой, а Alternaria alternata – известный фитопатоген согласно литературным данным [Соболев, 1999; de Hoog et al., 2000; Саттон, Фотергилл, Ринальди, 2001; Зачиняева, Лебедева, 2003].
Многие из выделенных нами видов грибов по опасности их для человека или по уровню патогенности относятся к 2 группам: BSL-1 и BSL-2 (см. табл. 2).
Грибы группы BSL-1 – это, в принципе, безопасные для здоровья людей. Но при нарушении кожных покровов они могут попадать в организм человека и вызывать локализованные микозы на фоне иммунодефицита. Грибы группы BSL-2 могут попадать в организм здорового человека и сохра- няться в нём, вызывая локализованные микозы. В ослабленном организме они могут распространяться более широко и проявлять свойства оппортунистов. В промышленной зоне их доля от видового разнообразия составляет 56%, тогда как в контроле – только 32%.
Аналогичным образом реагировали на загрязнение и фитопатогенные микромицеты. Как неблагоприятное воздействие загрязнения на экосистему можно расценивать и появление дереворазрушающего гриба Phoma glomerata на коре рябин промышленной зоны. А.Б. Кулько и О.Е. Марфениной [2001] также отмечено увеличение частоты встречаемости данного тёмноокрашенного пикнидиаль-ного гриба в снеговом покрове придорожной зоны.
Распространение потенциально патогенных микромицетов имеет определённую сезонную динамику. Наибольшее число оппортунистических микромицетов было обнаружено в летний и осенний периоды, 9 и 11 видов соответственно, а наименьшее (6 видов) в зимний период (табл. 3). По-видимому, низкие температуры воздуха оказываются лимитирующими для развития микробиоты. Схожие данные получены О.Е. Марфениной [2002], которая отмечает, что летний и осенний периоды являются самыми благоприятными временами года для развития опасных плесеней.
Обилие условно патогенных микромицетов
Таблица 3 коры древесных растений в разные сезоны
|
Виды микромицетов |
Время года |
|||||||
|
Весна |
Лето |
Осень |
Зима |
|||||
|
П |
К |
П |
К |
П |
К |
П |
К |
|
|
Alternaria alternata |
0.75 |
- |
1.44 |
1.93 |
2.71 |
1.9 |
1.52 |
- |
|
Aspergillus candidus |
- |
- |
- |
- |
1.2 |
- |
- |
- |
|
A. fumigatus |
2.15 |
1.95 |
3.6 |
0.6 |
3.35 |
1.7 |
2.33 |
0.85 |
|
A. nigervar.niger |
9.4 |
8.3 |
8.52 |
6.96 |
3.35 |
1.6 |
2.0 |
1.2 |
|
A. flavus var. oryzae |
- |
- |
1.6 |
- |
1.16 |
- |
- |
- |
|
A. terreus |
- |
- |
0.12 |
- |
- |
- |
- |
- |
|
Aureobasidium pullulans |
0.43 |
- |
- |
- |
- |
- |
- |
- |
|
Penicillium chrysogenum |
- |
- |
- |
- |
1.6 |
- |
- |
- |
|
P. citrinum |
- |
0.3 |
0.18 |
0.8 |
- |
0.3 |
0.12 |
0.12 |
|
P. glabrum |
- |
- |
- |
- |
0.5 |
- |
- |
- |
|
P. purpurogenum |
- |
0.75 |
- |
- |
- |
- |
- |
0.6 |
|
Trichoderma viride |
0.24 |
- |
0.7 |
0.96 |
0.57 |
0.12 |
0.5 |
- |
|
T. koningii |
1.6 |
- |
2.33 |
- |
1.16 |
0.75 |
- |
- |
|
T. harzianum |
- |
- |
- |
- |
2.0 |
0.24 |
- |
- |
|
Phoma glomerata |
- |
- |
0.12 |
- |
- |
- |
- |
- |
|
Всего |
6 |
4 |
9 |
5 |
10 |
7 |
5 |
4 |
|
Всего видов |
8 |
9 |
11 |
6 |
||||
Заключение
Таким образом, проведенные исследования образцов коры древесных растений позволили выявить 41 вид микроскопических грибов. Изучен- ные древесные растения по степени уменьшения видового разнообразия можно расположить в следующем порядке: Betula pendula Roth. – 23 вида → Populus nigra L. – 21 → Tilia cordata Mill. – 16 → Рicea obovata Ledeb. – 9→ Sorbus aucuparia L. – 6.
В зоне промышленного загрязнения наблюдалось упрощение видовой структуры комплекса микромицетов, при этом отмечено увеличение длины гиф и биомассы грибного мицелия на стволах древесных растений вблизи нефтеперерабатывающего комплекса. Также на загрязнённой территории обнаруживалось большее число условно патогенных для человека и животных видов микро-мицетов, чем на контрольной территории. Аналогичным образом реагировали на загрязнение и фи-топатогенные микромицеты, в промышленной зоне их количество возрастало по сравнению с таковыми фоновой территории.
Полученные результаты могут быть использованы для биоиндикации. Нами показано, что промышленное загрязнение нефтехимического комплекса приводит к изменениям в комплексах микроскопических грибов, обитающих на коре древесных растений. Это может способствовать накоплению грибного мицелия на коре с перераспределением доминирования к небольшому числу устойчивых видов, представляющих потенциальную опасность для здоровья человека.
Список литературы Влияние аэротехногенных загрязнений на состав сообществ эпифитных микромицетов г. Уфы
- Билай В.И., Коваль Э.З. Аспергиллы. Киев: Наук. думка, 1988. 204 с
- Билай В.И., Курбацкая З.А. Определитель токси-нобразующих микромицетов. Киев: Наук. думка, 1990. 234 с
- Волчатова И.В. и др. Микогенный ксилолиз в условиях антропогенного загрязнения // Лесоведение. 2007. № 5. С. 27-31
- Домрачева Л.И. и др. Альго-микологические и фи-тотоксические комплексы при химическом загрязнении почвы // Экология и почвы: лекции и докл. XIII Всерос. школы. Пущино, 2006. Т. 5. С. 88-98
- Дубовик И.Е., Климина И.П. Эпифитные водоросли в городских экосистемах Предуралья // Водоросли и цианобактерии в природных и сельскохозяйственных экосистемах: материалы Междунар. науч.-практ. конф. Киров, 2010. С. 107-110
- Дубовик И.Е., Киреева Н.А., Климина И.П. Эпифитные сообщества цианопрокариот, водорослей и микроскопических грибов древесных растений г. Уфы и возможность их использования в биоиндикации // V1 Междунар. конф. «Актуальные проблемы современной альгологии»: тез. докл. Киев, 2012. С. 104-105.
- Дубовик И.Е., Шарипова М.Ю. Наземные и водные эпифитные цианопрокариоты и водоросли и возможность их использования в оценке состояния окружающей среды // Теоретическая и прикладная экология. 2016. № 1. С. 51-55
- Евдокимова Г.А., Корнейкова М.В., Лебедева Е.В. Сообщества микромицетов в почвах в зоне воздействия алюминиевого завода // Микология и фитопатология. 2007. Т. 41, вып. 1. С. 20-28
- Зачиняева А.В., Лебедева Е.В. Микромицеты загрязненных почв северо-западного региона России и их роль в патогенезе аллергических форм микозов // Микология и фитопатология. 2003. Т.37, № 5. С. 69-74
- Кулько А.Б., Марфенина О.Е. Распространение микроскопических грибов в придорожных зонах городских автомагистралей // Микробиология. 2001. Т. 70, № 5. С. 709-713
- Марфенина О.Е. Реакция микроскопических грибов на загрязнение почв тяжёлыми металлами // Биологические науки. 1989. № 9. С. 89-93
- Марфенина О.Е. Антропогенные изменения комплексов микроскопических грибов в почвах: автореф. дис.... д-ра биол. наук. М., 1999. 50 с
- Марфенина О.Е. Опасные плесени в окружающей среде // Природа. 2002. № 11. С. 33-38
- Марфенина О.Е. Антропогенная экология почвенных грибов. М.: Медицина для всех, 2005. 196 с
- Марфенина О.Е. Фомичева Г.М. Потенциально патогенные мицелиальные грибы в среде обитания человека. Современные тенденции // Микология сегодня. М.: Национальная академия микологии, 2007. Т. 1. С. 235-266
- Методы почвенной микробиологии и биохимии / под ред. Д.Г. Звягинцева. М.: Изд-во МГУ, 1991. 304 с
- Милько А.А. Определитель мукоральных грибов. Киев: Наук. думка, 1974. 302 с
- Мирчинк Т.Г. Почвенная микология. М.: Изд-во МГУ, 1988. 220 с
- Одум Ю. Экология. М.: Мир, 1986. Т. 2. 376 с
- Саттон Д., Фотергилл А., Ринальди М. Определитель патогенных и условно патогенных грибов. М.: Мир, 2001. 468 с
- Сафонов М.А., Сафонова Т.И. Варьирование характеристик микоценозов в зависимости от уровня антропогенной нагрузки // Вестник Оренбургского государственного университета. 2009. № 6. С. 332-334
- Смирнов В.Ф. и др. Действие терпеноидов на физио-лого-биохимическую активность грибов-деструкторов промьппленньгх материалов // Химия растительного сырья. 2002. № 4. С. 29-33
- Соболев А.В. Значение микромицетов в патологии легких у человека // Проблемы медицинской микологии. 1999. Т. 1, № 3. С. 4-9
- Станкявичене Д., Урбонас В. Разнообразие агари-ковых грибов и эктомикоризы в градиенте антропогенного загрязнения в лиственном лесу // Микология и фитопатология. 2006. Т. 40, № 2. С. 108-116
- Терехова В.А. Микромицеты в экологической оценке водных и наземных экосистем. М.: Наука, 2007. 215 с
- de Hoog G.S. et al. Atlas of clinical fungi. Centraal-bureau voor Schimmelcultures. Utrecht: Universi-tat Rovira i Virgili, 2000. 1126 p
