Влияние блокады кисспептиновых рецепторов на показатели поведения крыс в стандартных тестах

Автор: Ткачева М.А., Инюшкина Е.М., Шарафутдинова А.Ю., Инюшкин А.Н.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 1, 2019 года.
Бесплатный доступ
Регуляторный пептид кисспептин известен как стимулятор фертильности и регулятор репродукции. Его активность опосредована специфическим взаимодействием с рецепторами GPR54 (или KisslR). Существуют указания на связь кисспептиновой системы мозга с функцией циркадианного осциллятора - супрахиазматического ядра гипоталамуса, стимулирующего клетки кисспептиновой нейронной сети антеровентрального перивентрикулярного ядра гипоталамуса, что в свою очередь приводит к высвобождению гипоталамического гонадолиберина в период времени, непосредственно предшествующий началу суточной локомоторной активности. Также имеются данные о наличии волокон, содержащих кисспептин, и экспрессии кисспептиновых рецепторов в супрахиазматическом ядре, что указывает на функцию кисспептина как сигнала обратной связи от репродуктивной к циркадианной системе. Наряду с репродуктивной функцией кисспептина, появляются данные о его участии в регуляции поведения. Цель работы - изучить влияние интраназального введения специфического антагониста GPR54 рецепторов р-234 на поведение самцов крыс в стандартных тестах «Открытое поле» и «Темносветлая камера»...
Антагонист кисспептиновых рецепторов р-234, поведенческие тесты
Короткий адрес: https://sciup.org/14116350
IDR: 14116350 | УДК: 612.826.4 | DOI: 10.34014/2227-1848-2019-1-103-109
Effect of kispeptin receptor blockade on behavioral indicators of rats in standard tests
Kisspeptin, the regulatory peptide, is known as a fertility stimulator and reproductive regulator. Its activity is mediated by specific interaction with GPR54 receptors (or Kiss1R). There are indications of correlation between the kisspeptin system in the brain and the function of circadian oscillator, the suprachiasmatic hypothalamus nucleus, which stimulates the cells of the kisspeptin neural network of the anteroventral periventricular nucleus in the hypothalamus, which in turn leads to the release of the hypothalamic gonadoliberin in the time period immediately preceding the start of the total daily locomotor activity. There is also evidence on the presence of fibers containing kisspeptin and on kisspeptin receptor expression in the suprachiasmatic nucleus, which indicates the function of kisspeptin as a feedback signal from the reproductive to the circadian system. Along with kisspeptin reproductive function, there is evidence on its importance in the behavior regulation...
Текст научной статьи Влияние блокады кисспептиновых рецепторов на показатели поведения крыс в стандартных тестах
Введение.* Регуляторный пептид кис-спептин был открыт в качестве супрессора метастатической активности [1], однако позже была доказана его роль в процессах полового созревания и гаметогенеза. Функциональная активность кисспептина объясняется его специфическим взаимодействием с рецепторами GPR54 (или Kiss1R) [2]. Взаимодействие кисспептина с рецепторами GPR54
играет важную роль в инициации секреции гонадотропин-рилизинг-гормона (GnRH) в период полового созревания [3]. Нарушение передачи сигналов через GPR54-рецепторы вследствие их мутации приводит к аутосомному рецессивному гипогонадизму [4]. Существуют указания на связь кисспептиновой системы мозга с функцией циркадианного осциллятора – супрахиазматического ядра гипоталамуса [5]. В частности, в экспериментах на грызунах показано, что супрахиазма-тическое ядро ежесуточно в любой день эстрального цикла посылает стимулирующий сигнал, передатчиком которого является кис- спептин, в направлении гипоталамических клеток, продуцирующих гонадолиберин, в период циркадианного цикла, непосредственно предшествующий началу активной локомоторной фазы [6]. Таким образом, кис-спептин способен играть важную роль в сопряжении функций циркадианной и репродуктивной систем, а время суток может оказывать влияние на характер эффектов кис-спептина. В этом плане особый интерес представляют иммуногистохимические данные о наличии большого количества волокон, содержащих кисспептин, и экспрессии кис-спептиновых рецепторов в супрахиазматиче-ском ядре [7]. В нашем исследовании in vitro на срезах гипоталамуса крыс аппликации кисспептина вызывали рост спайковой активности нейронов супрахиазматического ядра и снижение степени нерегулярности межспайковых интервалов [8]. Эти данные свидетельствуют в пользу непосредственного действия репродуктивного регулятора кис-спептина на уровне нейронов циркадианного осциллятора по принципу обратной связи.
Наряду с репродуктивной функцией кис-спептина в последнее время отмечается его участие в регуляции поведения. В частности, у самцов мышей с нокаутом гена Kiss1R обнаружено изменение уровня тревожности и исследовательского поведения [9]. Подобные исследования определяют необходимость детального изучения поведенческой функции кисспептина и его рецепторов. В настоящей работе изучали влияние интраназального введения специфического антагониста GPR54 рецепторов р-234 на поведение самцов крыс в стандартных тестах «Открытое поле» и «Темно-светлая камера».
Цель исследования. Изучение процессов регуляции циркадианного осциллятора при помощи антагониста кисспептина (р-234).
Материалы и методы. Все экспериментальные протоколы были согласованы и одобрены комиссией по биологической этике Самарского национального исследовательского университета им. академика С.П. Королева. Исследовано 12 животных контрольной и экспериментальной групп. Для экспериментов были взяты крысы-самцы «Вистар» массой тела 240–270 г. Все животные содер- жались в виварии биологического факультета Самарского университета и получали пищу и питьевую воду без ограничений. Животных содержали в условиях режима освещения «свет/темнота» 12:12 ч. В начале эксперимента в течение 10 мин изучали поведение крыс в установках «Открытое поле» и «Тёмно-светлая камера» в первой половине циркадианного дня в ZT=2–6 (ZT=0 – момент включения освещения). Далее в то же время дня в течение трёх последовательных суток интраназально в объёме 10 мкл экспериментальным животным вводили антагонист кис-спептиновых рецепторов р-234 в концентрации 10-4 М. Крысам контрольной группы аналогично вводили воду для инъекций в объёме 10 мкл. Через 15 мин после интраназального введения крыс последовательно помещали в установки «Открытое поле» и «Тёмно-светлая камера» и в течение 10 мин производили видеорегистрацию поведенческой активности. Видеорегистрацию поведения животных осуществляли при помощи камеры Panasonic fullhd hc-x810, которая была установлена на штативе. При помощи программ ANY-maze 4.99 и SigmaPlot 12.5 осуществляли автоматическую регистрацию и статистическую обработку исследуемых параметров поведения. В тесте «Открытое поле» определяли количество посещенных квадратов, количество обследованных отверстий, время нахождения в центральных и периферических квадратах, характеристики груминга, седации (замирания), количество стоек с опорой на бортик, количество стоек без опоры на бортик. В тесте «Тёмно-светлая камера» определяли продолжительность нахождения в светлом и темном отсеках, количество выходов в светлый отсек, заглядывания из тёмного в светлый отсек, заглядывания из светлого в тёмный отсек.
Значения исследуемых показателей в ходе эксперимента сравнивали в контрольной и экспериментальной группах. В случае нормального распределения данных в выборках для сравнения использовали непарный t-тест, в остальных случаях – тест Манна–Уитни. Нормальность распределения проверяли с помощью теста Шапиро–Уилка, равенство дисперсий – с помощью теста Левена. Стати- стические данные о значениях параметров, соответствующих нормальному распределению, представляли как средние арифметические ± стандартные ошибки среднего. Изменения исследуемых параметров считали статистически значимыми при р<0,05.
Результаты. В исходном состоянии исследуемые показатели поведения у животных контрольной и экспериментальной групп не различались (р>0,05). После интраназального введения антагониста кисспептиновых рецепторов р-234 значение некоторых поведен- ческих показателей отличалось относительно соответствующих значений в контроле.
В установке «Открытое поле» не было обнаружено изменений количества посещённых квадратов, количества обследованных отверстий, времени нахождения в центральных и периферических квадратах, характеристик груминга (р>0,05). Однако, как показано на рис. 1, введение антагониста вызвало статистически значимое увеличение значения показателя «седация» (замирание) относительно контроля (р=0,046).
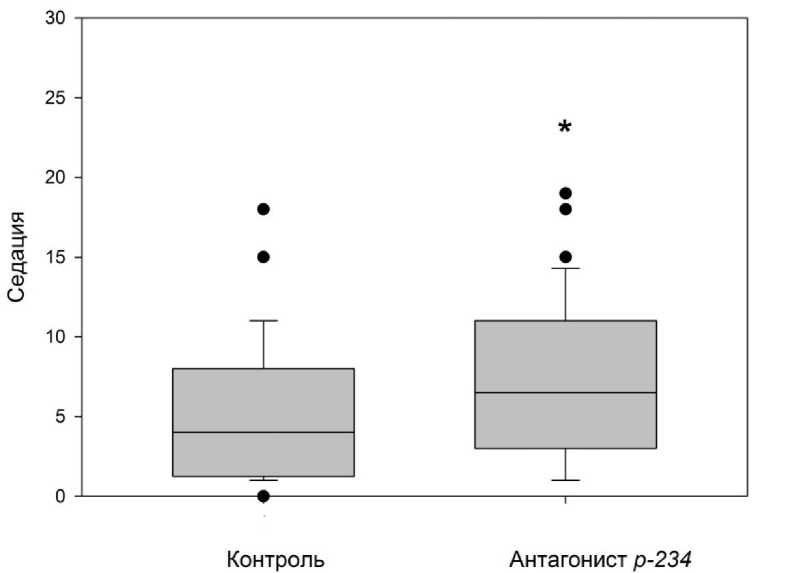
Рис. 1. Влияние интраназального введения антагониста кисспептиновых рецепторов р-234 на поведенческий параметр «седация».
Данные представлены как медианы (линия внутри боксов), квартили (верхняя и нижняя границы боксов), 10-й и 90-й перцентили (нижняя и верхняя границы усов), значения вариант за пределами этих перцентилей (точки).
* – статистически значимое различие между контрольной и экспериментальной группами (р<0,05) (на рис. 2, 3 обозначения аналогичные)
Исследование параметра «количество стоек с опорой на бортик» не выявило статистически значимых изменений (р=0,373). Вместе с тем введение антагониста привело к статистически значимому увеличению количества стоек без опоры на бортик (р=0,033; рис. 2).
Анализ параметров поведения в тесте «Темно-светлая камера» дал следующие ре- зультаты. Интраназальное введение антагониста кисспептиновых рецепторов р-234 не привело к изменению по сравнению с контролем продолжительности пребывания в темном и светлом отсеках (р>0,05). При этом у крыс, получавших антагонист кисспепти-новых рецепторов, обнаружилось увеличение количества выходов в светлый отсек по сравнению с контролем (р=0,017; рис. 3).
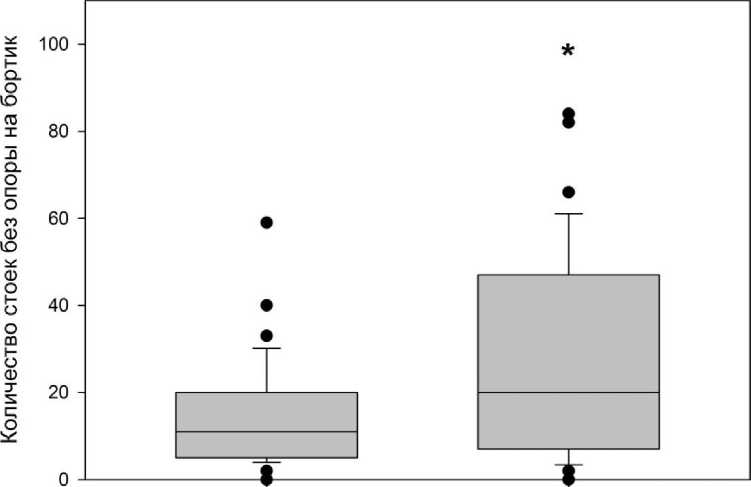
Контроль
Антагонист р-234
Рис. 2. Влияние интраназального введения антагониста кисспептиновых рецепторов р-234 на поведенческий параметр «количество стоек без опоры на бортик»
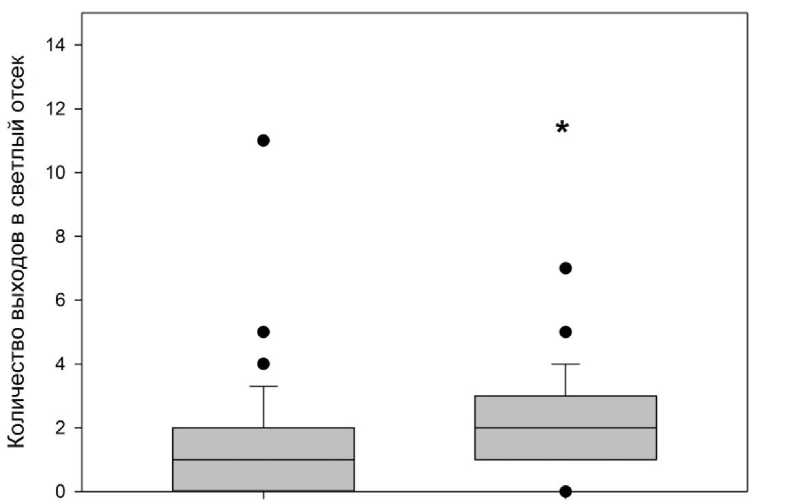
Контроль
Антагонист р-234
Рис. 3. Влияние интраназального введения антагониста кисспептиновых рецепторов р-234 на поведенческий параметр «количество выходов в светлый отсек»
Значения параметра «заглядывание из тёмного в светлый отсек» не различались в контрольной и экспериментальной группах
(p=0,146), так же как и значения параметра «заглядывание из светлого в тёмный отсек» (p=0,139).
Обсуждение. В ходе настоящей работы изучали влияние антагониста кисспептино-вых рецепторов р-234 на показатели поведения самцов крыс. При этом использовали ин-траназальное введение вещества. Данный способ введения становится все более популярным в экспериментальной практике, что объясняется целым рядом преимуществ. Вещества, вводимые интраназально, быстро проникают непосредственно в головной мозг в значительной концентрации вдоль обонятельных и тройничных нервов, минуя гематоэнцефалический барьер [10]. При этом вводимое вещество практически не попадет в кровоток [11], что дает возможность изучать центральные эффекты вещества изолированно от его периферических эффектов, возникающих при системном введении. Таким образом, в настоящей работе выявлены изменения поведенческой активности животных, возникающие в условиях блокады центральных кисспептиновых рецепторов.
Поскольку обнаружен циркадианный ритм стимуляции кисспептином структур репродуктивной оси [6], существенным является время введения вещества в суточном цикле. В нашем исследовании антагонист р-234 вводили в начале субъективного дня (ZT=2–6), когда кисспептиновые рецепторы практически не связаны с эндогенным кисспептином.
Основным результатом работы является изменение ряда поведенческих показателей в условиях блокады кисспептиновых рецепторов. В частности, в поведенческой установке «Открытое поле» обнаружено увеличение параметров «седация» и «стойка без опоры на бортик». В тесте «Темно-светлая камера» увеличилось количество выходов в светлый отсек. Такие изменения свидетельствуют о том, что введение антагониста кис-спептиновых рецепторов приводит к снижению уровня тревожности и к стимуляции исследовательского поведения. Полученные результаты хорошо согласуются с данными исследования на самцах мышей с нокаутом кисспептиновых рецепторов, у которых также было обнаружено снижение уровня тревожности и стимуляция исследовательского поведения в тесте «Приподнятый крестообразный лабиринт» [9]. Выявленные изменения поведения могут способствовать реализации уже известной функции кис-спептина как стимулятора репродукции либо выступать в качестве самостоятельного компонента в спектре физиологической активности кисспептиновой системы. Для ответа на этот вопрос необходимы дальнейшие исследования.
Заключение. Установлено, что антагонист кисспептиновых рецепторов р-234 при интраназальном введении самцам крыс вызывает снижение уровня тревожности и стимулирует исследовательское поведение.
Список литературы Влияние блокады кисспептиновых рецепторов на показатели поведения крыс в стандартных тестах
- Lee J.H., Miele M.E., Hicks D.J., Phillips K.K., Trent J.M., Weissman B.E., Welch D.R. KiSS-1, a novel human malignant melanoma metastasis-suppressor gene. J. Natl. Cancer. Inst. 1996; 88: 1731-1737.
- Ohtaki T., Shintani Y., Honda S., Matsumoto H., Hori A., Kanehashi K., Terao Y., Kumano S., Takat-su Y., Masuda Y., Ishibashi Y., Watanabe T., Asada M., Yamada T., Suenaga M., Kitada C., Usuki S., Kurokawa T., Onda H., Nishimura O., Fujino M. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature. 2001; 411 (6837): 613-617.
- Hrabovszky E., Ciofi P., Vida B., Horvath M.C., Keller T., Caraty F., Bloom S.R., Ghatei M.F., Dhillo W.S., Liposits Z., Kallo I. The kisspeptin system of the human hypothalamus: sexual dimorphism and relationship with gonadotropin-releasing hormone and neurokinin B neurons. Eur. J. Neurosci. 2010; 31: 1984-1998.
- Semple R.K., Achermann J.C., Ellery J., Farooqi I.S., Karet F.E., Stanhope R.G., O'Rahilly S., Apari-cio S.A. Two novel missense mutations in G protein-coupled receptor 54 in a patient with hypogonado-tropic hypogonadism. J. Clin. Endocrinol. Metab. 2005; 90: 1849-1855.
- Ткачева М.А., Инюшкина Е.М., Карян С.Д., Инюшкин А.Н. Роль аргинин-вазопрессина в регуляции активности нейронов циркадианных часов супрахиазматического ядра гипоталамуса. Ульяновский медико-биологический журнал. 2018; (1): 145-154.
- Kriegsfeld L.J. Circadian regulation of kisspeptin in female reproductive functioning. Adv. Exp. Med. Biol. 2013; 784: 385-410.
- Simonneaux V., Ancel C., Poirel V.J., Gauer F. Kisspeptins and RFRP-3 act in concert to synchronize rodent reproduction with seasons. Front. Neurosci. 2013; 7: 22.
- Ткачева М.А., Инюшкин А.Н. Электрофизиологические эффекты кисспептина на уровне нейронов биологических часов супрахиазматического ядра. Врач-аспирант. 2017; 80 (1): 86-92.
- Delmas S., Porteous R., Bergin D.H., Herbison A.E. Altered aspects of anxiety-related behavior in kisspeptin receptor-deleted male mice. Sci. Reports. 2018; 8: 2794.
- Frey W.H. Bypassing the blood-brain barrier to deliver therapeutic agents to the brain and spinal cord. Drug Deliv. Tech. 2002; 2: 46-49.
- Born J., Lange T., Kern W., McGregor G.P., Bickel U., Fehm H.L. Sniffing neuropeptides: a transnasal approach to the human brain. Nat. Neurosci. 2002; 5: 514-516.
