Влияние даларгина на активность Na, K-АТФазы мембран синаптосом из коры головного мозга крыс при ишемии и реперфузии

Автор: Мохаммед М.Т., Кличханов Н.К.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 1-1 т.13, 2011 года.
Бесплатный доступ
Исследовано влияние даларгина на активность Na, K АТФазы мембран синаптосом из коры головного мозга крыс при ишемии и реперфузии. Ишемия, вызванная путем окклюзии двух сонных артерий, приводит к ингибированию активности фермента. Степень снижения активности Na, K АТФазы зависела от длительности ишемии (30, 60, 90 мин). В период реперфузии (60 и 90 мин), после 60 мин ишемии, активность Na, K АТФазы возрастает относительно ишемического уровня. Внутрибрюшинное введение даларгина за 30 мин до ишемии в дозе 0,5 мг/кг, но не в дозе 0,1 мг/кг, предотвращает ингибирование фермента при ишемии.
K атфаза, ишемия, даларгин, синаптические мембраны, мозг крысы
Короткий адрес: https://sciup.org/148199592
IDR: 148199592 | УДК: 577.352.3
Effect of dalargin on the activity of Na, K-ATPhase in cerebral cortex synaptosomal membranes of rats in ischemia and reperfusion
Effect of a peptide dalargin on the activity Na, K ATPase in rats cerebral cortex synaptosomal membrane during ischemia and reperfusion was studied. Ischemia caused by occlusion of the two carotid arteries, leads to inhibition of enzyme activity. The degree of decrease in activity of Na, K ATPase dependes on the duration of ischemia (30, 60, 90 min). During reperfusion (60 and 90 min), after 60 min of ischemia, the activity Na, K ATPase increases relatively ischemic level. Dalargin intraperitoneal injection (30 min) before ischemia with dose 0,5 mg/kg, but not with dose of 0,1 mg/kg, prevents the inhibition of the enzyme during ischemia.
Текст научной статьи Влияние даларгина на активность Na, K-АТФазы мембран синаптосом из коры головного мозга крыс при ишемии и реперфузии
N a, K-АТФаза является интегральным мембранным ферментом, использующим энергию гидролиза АТФ для транспорт ионов Na+ и K+ против электрохимического градиента. В клетках мозга градиенты ионов натрия и калия, создаваемые этим ферментом, необходимы для осуществления таких физиологических функций как регуляция объема, поддержание электрического потенциала, вторичный активный транспорт, а также обеспечение осмотического баланса в системе нейрон-экстраклеточный компартмент [5].
Нарушение снабжения кислородом тканей мозга, наступающее при ишемическом повреждении мозга, приводит к снижению активности Na, K-АТФазы [13]. В результате подавления ее активности нарушается транспорт ионов, развивается деполяризация мембраны, нейроны утрачивают свою важнейшую функцию — электрическую проводимость. Это может привести к чрезмерной секреции нейромедиаторов, перегрузке нейронов ионами кальция, и далее к вторичному ишемическому повреждению, т.е. активации фосфолипаз, липаз, протеаз, эндонуклеаз, неконтролируемого фосфорилирования, деградации мембран, и отёку мозга. Вместе с тем пока неясно как зависит степень ингибирования Na, K-АТФазы мозга от длительности ишемии и последующей реперфузии.
D-Ala2, Leu5, Агд6-энкефалин (даларгин) является синтетическим аналогом лей-энкефалина [3]. Этот пептид активирует р- и 6-рецепторы и проникает через гематоэнцефалический барьер только при его использовании в дозах не менее 0,5 мг/кг [9]. В экспериментах на животных с острой ишемией миокарда, вызванной окклюзией коронарной артерии, получены данные о благоприятном действии даларгина [7]. Даларгин оказывает противоишемическое действие при снижении перфузионного давления в системе кровоснабжения мозга при реконструктивных операциях на сонных и позвоночных артериях мозга [1]. Целью
данной работы было выяснение зависимости активности Na, K-АТФазы мембран синаптосом из коры головного мозга крыс от длительности ишемии и реперфузии, а также роли опиоидного нейропептида даларгина в защите фермента от ишемического повреждения.
МЕТОДИКА ИССЛЕДОВАНИЯ
Опыты выполнены на 46 белых крысах-самцах Вистар, массой 200-230 г. Экспериментальную ишемию головного мозга вызывали окклюзией двух сонных артерий в течение 30, 60 и 90 мин. У части животных после 60-минутной окклюзии двух сонных артерий моделировали реперфузию путем снятия лигатуры. Продолжительность реперфузии составила 60 и 90 мин. У животных контрольной группы воспроизводилась наркотизация, кожный разрез и выделение артерий без последующей перевязки сосудов. Все хирургические процедуры проводили под наркозом (внутрибрюшное введение тиопентала натрия в дозе 40-50 мг/кг). В период ишемии и реперфузии температура тела животного поддерживали на нормальном уровне (37°С).
Фармакопейный препарат даларгин (НПО «Микроген») вводили внутрибрюшинно в дозе 0, 1 и 0, 5 мг/кг за 30 мин до наложения лигатуры на сонные артерии. Контрольным (ложноопери-рованным) животным вводили соответствующий объем физиологического раствора.
После декапитации крыс для исследования использовалась кора больших полушарий головного мозга. Синаптосомы выделяли методом центрифугирования [16]. Полученные синаптосомы подвергали осмотическому шоку в дистиллированной воде, после чего синаптические мембраны осаждали при 20000 g в течение 30 мин. После однократной промывки 0,32 М сахарозой препарат мембран хранили при -70°С и использовали для анализа в течение недели. Активность Na, К-АТФазы в мембранах синаптосом оценивали, измеряя количество неорганического фосфата (Рн), освобождающегося в ходе гидролиза АТФ [10]. Инкубационная среда для определения общей АТФазной активности содержала (в мМ/л): 130
- NaCl, 20 — KCl, 3 — MgCl2, 30 — трис-HCl буфер, pH — 7,4, 3 — АТФ и 40 мкг мембранного белка в 1,5 мл конечного объема. При определении активности Мд2+-АТФазы среда инкубации содержала те же компоненты и дополнительно 1 мМ уабаина — специфического ингибитора Na, К-АТФазы. Активность Na, К-АТФазы рассчитывали как разность между общей и Мд2+-зависимой АТФазной активностью. Содержание белка определяли по Лоури.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Как видно на рисунке, при ишемии активность Na, К-АТФазы мембран синаптосом снижается. Ингибирование активности фермента после 30, 60, 90 мин ишемии составило 32,1%, 45,8%, 53,7% относительно контроля (ложнооперированные животные) соответственно. Таким образом, степень ингибирования фермента зависит от длительности ишемии.
Ингибирование фермента снижает трансмембранные градиенты ионов, вызывает чрезмерное поступление в нейроны Na+ с последующим притоком Cl- и воды, что приводит к клеточному отеку. Обнаружена корреляция между ингибированием Na, К-АТФазы и отёком мозга в ишемизированной области [18].
Ишемия вызывает на клеточном уровне серию быстро развивающихся во времени изменений, которые могут повлиять на Na, К-АТФазу. В их числе ацидоз, перегрузка ионами кальция, активация протеаз, аккумуляция метаболитов липидов (жирных кислот, лизолипидов) и в некоторых тканях, транслокация ферментного белка с поверхности мембраны внутрь клетки [14]. Не исключено также, что обнаруженное нами ингибирование Na, К-АТФазы связано с потерей связи фермента с цитоскелетом. Цитоплазматический домен а-субъединицы Na, К-АТФазы связан с анкирином, который связывает фермент с элементами примембранного цитоскелета (фодрино-выми фибриллами). Такая связь с цитоскелетом стабилизирует Na, К-АТФазу в плазматической мембране. а-Фодрин и анкирин являются субстратами кальпаинов — группы нелизосомальных Ca2+-зависимых протеаз. Показано, что на раннем этапе реперфузии (5 мин) после тотальной ишемии миокарда резкое снижение активности Na, К-АТФазы связано с активацией кальпаинов, что приводит к деградации цитоскелетных белков и нарушению их связи с ферментом [17]. Активация селективного протеолиза а-фодрина под действием кальпаинов обнаружена в синаптосомах из мозга крыс при ишемии и реперфузии [15].
Ингибирование активности фермента при ишемии может происходить под действием активных форм кислорода (АФК), окислительной модификации липидного микроокружения и накопления продуктов их деградации, изменения редокс-состояния окружения, фосфорилирования субъединиц [19]. При ишемии Na, К-АТФаза функционирует в условиях высокой концентрации АФК, которые приводят к ингибированию гидролитической и транспортной функции Na, К-АТФазы. На частично очищенном препарате фермента из синаптосом мозга показано, что степень ингибирования Na, К-АТФазы зависит от вида АФК [11].
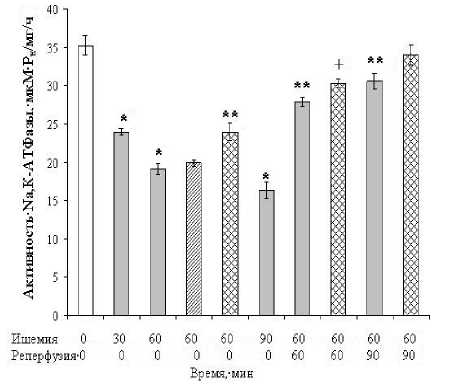
Рис. Изменение активности Na, К-АТФазы мембран синаптосом из коры головного мозга крыс при ишемии/реперфузии и введении даларгина: Светлый столбик - контрольные (ложнооперированные) животные; серые столбики - ишемия и реперфузия; столбики с косой штриховкой - введение даларгина в дозе 100 мкг/кг; дважды заштрихованные столбики - введение даларгина в дозе 500 мкг/кг. * - р < 0,05 относительно контроля; ** - р < 0,05 относительно ишемии 60 мин; +— р < 0,05 относительно ишемии 60 мин + реперфузия 60 мин.
Данные для построения рисунка
|
№ |
Группа |
Активность Na, К-АТФазы, мкмоль Р „ /мг/ч |
|
1 |
Контроль |
35,23+1,28 |
|
2 |
Ишемия 30 мин |
23,93+0,46 |
|
3 |
Ишемия 60 мин |
19,09+0,73 |
|
4 |
даларгин 100 мкг + ишемия 60 мин |
19,86+0,42 |
|
5 |
даларгин 500 мкг + ишемия 60мин |
23,95+1,14 |
|
6 |
Ишемия 90 мин |
16,31+1,12 |
|
7 |
Ишемия 60 мин+ реперфузия 60 мин |
27,86+0,62 |
|
8 |
Даларгин 500 мкг+ишемия 60 мин+ реперфузия 60 мин |
30,27+0,52 |
|
9 |
Ишемия 60 мин+ реперфузия 90 мин |
30,53+1,04 |
|
10 |
Даларгин 500 мкг+ишемия 60 мин+ реперфузия 90 мин |
33,96+1,34 |
Исследование фермента в постишемическом периоде показало, что снятие лигатуры и восстановление кровоснабжения мозга после 60 мин ишемии приводит к повышению активности Na, К-АТФазы. Через 60 мин реперфузии активность фермента возрастает на 45,9%, а после 90 мин -на 59,9% относительно ишемии. Таким образом, при ишемии/реперфузии происходит обратимое ингибирование Na, К-АТФазы. Это позволяет предположить, что ингибирование фермента при ишемии связано с изменением редокс-состояния тиоловых групп и/или фосфорилирования субъединиц, которое может привести к обратимой интернализации Na, К-АТФазы в клатрин окаймленные везикулы [12] и уменьшению количества ее молекул на синаптической мембране.
Введение даларгина в дозе 0,1 мг/кг за 30 мин до ишемии не оказало влияние на активность Na, К-АТФазы после 60 мин ишемии (рис.). В дозе 0,5 мг/кг активность фермента после 60 мин ишемии на 25% выше, чем при ишемии без введения пептида. Таким образом, даларгин, в дозе проникающей в головной мозг, защищает Na, К-АТФазу синаптических мембран от ишемического повреждения.
В связи с полученными нами результатами интерес представляют данные литературы о влиянии даларгина на сердечно-сосудистую систему. Внутривенное введение даларгина вызывало увеличение частоту сердечных сокращений как у человека (25-100 мкг/кг), так и у крыс (100 мкг/кг), и этот эффект не проявлялся в условиях блокады опиоидных рецепторов налоксоном [4, 6]. Показано, что курсовое введение даларгина вызывает снижение удельного периферического сопротивления [8], оказывает венодилятирующее действие, а также уменьшает потребление кислорода организмом. Считают, что даларгин, являясь р-агонистом опиоидных рецепторов, вызывает вазодилятацию и снижение артериального давления за счет увеличения продукции NO эндотелием [6]. Кроме того, внутривенная инфузия пептидных агонистов р-опиоидных рецепторов вызывала у наркотизированных крыс снижение общего периферического сопротивления и артериального давления. Подобных опиоид-индуцированных изменений показателей центральной гемодинамики не происходит в условиях блокады NO-синтазы. Исходя из этих данных можно предположить, что благоприятный эффект даларгина при ишемии связан как со снижением потребности клеток мозга в кислороде, так и с улучшением кровоснабжения мозга за счет интенсификации коллатерального кровоток благодаря сосудорасширяющему эффекту пептида.
Даларгин способен оказать и антиоксидантный эффект. Показано, что пептид при внутривенном введении способен снижать активность ксанти-ноксидазы мозга [2], катализирующей образование супероксиданионрадикала и пероксида водорода.
Таким образом, ишемия, в зависимости от ее длительности, приводит к ингибированию активности Na, К-АТФазы мембран синаптосом. В период реперфузии активность фермента восстанавливается. Внутрибрюшинное введение даларгина до ишемии в дозе 0,5 мг/кг, предотвращает ингибирование фермента при ишемии.
Список литературы Влияние даларгина на активность Na, K-АТФазы мембран синаптосом из коры головного мозга крыс при ишемии и реперфузии
- Казанцев В.В., Луцик А.А., Тюлькин О.Н. Защита головного мозга в хирургии экстракраниальных отделов сонных и позвоночных артерий/Сосудистые заболевания головного и спинного мозга: сб. науч. тр.; под ред. А. Ю. Савченко. Омск: Издательство ОмГПУ. 2000. С. 30-34.
- Короткина Р.Н., Шлозников Б.М., Донич С.Г., Гребенчиков О.А., Ситников А.В., Карелин А.А. Изучение активности ксантиноксидазы в ткани головного мозга на фоне миоплегии//Бюлл. экспер. биол. и мед. 1990. Т. 59, № 2. С. 145-146.
- Лишманов Ю.Б., Маслов Л.М. Опиатные нейропептиды, стресс и адаптационная защита сердца. Томск: Изд-во Том. ун-та. 1994. 352 с.
- Лишманов Ю.Б., Маслов Л.М. Опиатергическая регуляция состояния центральной гемодинамики//Пат. физиол. эксперим. терапия. 2003. №1. С. 2-10.
- Лопина О.Д. Взаимодействия каталитической субъединицы Na, К-АТРазы с клеточными белками и другими эндогенными регуляторами//Биохимия. 2001. Т. 66, вып. 10. С. 1389-1400.
- Маслов Л.В. Лишманов Ю.Б., Лопотухин Э.Ю. Даларгин -это пептидный агонист μ-и δ-опиоидных рецепторов//Клинич. фармакол. и терапия. 2004. № 4. С. 47-52.
- Маслов Л.М., Лишманов Ю.Б., Гросс Г.Дж., Стефано Дж. Феномен повышенной устойчивости сердца к аритмогенному действию ишемии и реперфузии при активации периферических опиатных рецепторов.//Вестн. аритмол. 2002. № 26. С. 77-90.
- Маслов Л.Н., Федорова Н.А., Дудко ВА., Карпов Р.С. Влияние агониста опиатных периферических рецепторов даларгина на толерантность к физической нагрузке пациентов с атеросклерозом коронарных и периферических артерий//Физиол. человека. 2002. Т. 28, № 4.С. 1-7.
- Полонский В.М., Ярыгин К.Н., Кривошеев И.Г. Место приложения (центральное или периферическое) противоязвенного действия синтетического аналога эндогенных опиоидов даларгина в экспериментальной модели цистеаминовых дуоденальных язв у крыс//Бюлл. экспер. биол. и мед. 1987. № 4. С. 433-434.
- Рожанец В.В., Козлова В.Н., Родина Р.И., Швец В.И., Глебов Р.Н. Действие противосудорожных веществ на Na, K-АТФазу синаптических мембран головного мозга животных//Биохимия. 1978. Т. 43, вып. 5. С. 892-898.
- Bogdanova A., Petrushanko I., Boldyrev A., Gassmann M. Oxygen-and redox-induced regulation of the Na/K ATPase//Current Enzyme Inhibition. 2006. Vol. 2. P. 37-59.
- Dada L. A., Chandel N.S., Ridge K. M., Pedemonte C., Bertorello A. M., Sznajder J. I. Hypoxia-induced endocytosis of Na, K-ATPase in alveolar epithelial cells is mediated by mitochondrial reactive oxygen species and PKC-ζ//J. Clin. Invest. 2003. Vol. 111. P. 1057-1064.
- Dobrota D., Matejovicova M., Kurella E.G., Boldyrev A.A. Na/K-ATPase under oxidative stress: molecular mechanisms of injury//Cell Mol. Neurobiol. 1999. Vol. 19. P. 141-149.
- Durukan A., Tatlisumak T. Acute ischemic stroke: overview of major experimental rodent models, pathophysiology, and therapy of focal cerebral ischemia//Pharmacol. Biochem. Behav. 2007. Vol. 87. P. 179-197.
- Fukuda S., Harada K., Kunimatsu M., Sakabe T., Yoshida K. Postischemic reperfusion induces α-fodrin proteolysis by m-calpain in the synaptosome and nucleus in rat brain//J. Neurochem. 1998. Vol. 70. P. 2526-2532.
- Hajos F. An improved method for the preparation of synaptosomal fractions in high purity//Brain Res. 1975. Vol. 93, №. 3. P. 484-489.
- Inserte J., Garcia-Dorado D., Hernando V., Soler-Soler J. Calpain-mediated impairment of Na+/K+-ATPase activity during early reperfusion contributes to cell death after myocardial ischemia//Circulation Res. 2005. Vol. 97. P.465-473.
- Mintorovitch J., Yang G.Y., Shimizu H., Kucharczyk J., Chan P.H., Weinstein P.R. Diffusion-weighted magnetic resonance imaging of acute focal cerebral ischemia: comparison of signal intensity with changes in brain water and Na+, K+-ATPase activity//J. Cereb. Blood Flow Metab. 1994. Vol. 14. P. 332-336.
- Wang X. Q., Xiao A. Y., Sheline C., Hyrc K., Yang A., Goldberg M.P. Apoptotic insults impair Na+, K+-ATPase activity as a mechanism of neuronal death mediated by concurrent ATP deficiency and oxidant stress//J. Cell Sci. 2003. Vol. 116. P. 2099-2110.
