Влияние экстремальных факторов высокогорья на ультраструктуру лейкоцитов крови крыс

Автор: Бектурганова Асель Орумбековна, Махмудова Жылдыз Акматовна, Аскалиева Нургуль Ракыевна, Ниязалиева Джамиля Карыпбековна, Таалайбекова Мээрим Таалайбековна
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 5 т.9, 2023 года.
Бесплатный доступ
В работе представлены результаты исследования по изучению ультраструктуры клеток белой крови под влиянием факторов высокогорья в сочетании с холодом. Материалы и методы: 70 белых беспородных лабораторных крыс-самцов с массой тела 200-300 г; метод электронно-микроскопического исследования, статистические методы. Результаты исследования. Установлено, что у крыс при адаптации к условиям высокогорья в сочетании с воздействием холода идут резкие изменения ультраструктуры лейкоцитов. У экспериментальных животных на 3-и сутки наблюдения уровень объемной плотности митохондрий повышался почти в 2 раза в нейтрофилах, в 1,5 в эозинофилах и в 1,4 раза в моноцитах и средних лимфоцитах. С увеличением срока пребывания в высокогорье в сочетании с холодовым воздействием объемная плотность митохондрий продолжала возрастать во всех видах клеток белой крови, и на 30-е сутки адаптации оставалась статистически значимо выше и в сравнении с контролем, и в сравнении с показателями группы животных на 3-и сутки адаптации. Объемная плотность рибосом также статистически значимо возрастала на 3-и сутки адаптации к высокогорью в сочетании с холодом в нейтрофилах, эозинофилах и лимфоцитах, и продолжала увеличиваться к 30-м суткам наблюдения. Тенденция к увеличению объемной плотности была отмечена и у вакуолей клеток: на 3-и сутки адаптации было зафиксировано возрастание показателя объемной плотности вакуолей по сравнению с контролем у нейтрофилов. К 30-му дню адаптации объемная плотность вакуолей по сравнению с животными на 3-й день адаптации статистически значимо снижалась, хотя так и не достигала показателей животных контрольной группы. Объемная плотность специфических гранил на 3-и сутки пребывания в горах в нейтрофилах и эозинофилах снизилась почти на 100%.
Ультраструктура клетки, физиологическая адаптация, горы, холод
Короткий адрес: https://sciup.org/14127974
IDR: 14127974 | УДК: 599.323.41 | DOI: 10.33619/2414-2948/90/10
Influence of extreme factors of high mountains on the ultrastructure of rat blood leukocytes
This paper presents the results of a study on the ultrastructure of white blood cells under the influence of high-altitude factors in combination with cold. Materials and methods: 70 outbred male laboratory rats weighing 200-300 g; method of electron microscopic examination, statistical methods. Research results. It has been established that in rats, during adaptation to high altitude conditions, in combination with exposure to cold, there are sharp changes in the ultrastructure of leukocytes. In experimental animals on the 3rd day of observation, the level of volumetric density of mitochondria increased almost 2 times in neutrophils, 1.5 times in eosinophils, and 1.4 times in monocytes and medium lymphocytes. With an increase in the length of stay in the highlands in combination with cold exposure, the volumetric density of mitochondria continued to increase in all types of white blood cells, and on the 30th day of adaptation it remained statistically significantly higher both in comparison with the control and in comparison, with the indices of the animal group on the 3rd and day of adaptation. The volumetric density of ribosomes also increased statistically significantly on the 3rd day of adaptation to high mountains in combination with cold in neutrophils, eosinophils, and lymphocytes, and continued to increase by the 30th day of observation. A trend towards an increase in bulk density was also noted in cell vacuoles: on the 3rd day of adaptation, an increase in the volumetric density of vacuoles was recorded compared to the control in neutrophils. By the 30th day of adaptation, the bulk density of vacuoles, compared with animals on the 3rd day of adaptation, statistically significantly decreased, although it did not reach the indicators of animals in the control group. The bulk density of specific granites on the 3rd day of stay in the mountains in neutrophils and eosinophils decreased by almost 100%.
Текст научной статьи Влияние экстремальных факторов высокогорья на ультраструктуру лейкоцитов крови крыс
Бюллетень науки и практики / Bulletin of Science and Practice
Как известно, при изучении проблемы формирования биоадаптаций необходимо базироваться не только на основе различных данных эколого-физиологических аспектов приспособительных реакций, но и учитывать морфологические признаки формирования адаптивных процессов в организме. Реакция организма на действие раздражителей, как правило, сопровождается достаточно выраженными морфологическими и физикохимическими изменениями в тканях, возникающих под воздействием экстремальных факторов внешней среды. Среди множества разнообразных факторов внешней среды одним из наиболее значимых с точки зрения эколого-физиологического воздействия, является холод. Температура воздуха оказывает огромное влияние на жизненно-важные функции организма, такие как обмен веществ, терморегуляция, дыхание, кровообращение и др. [1–4].
Цель исследования — изучение динамики изменений морфологических показателей ультраструктуры клетки белой крови у экспериментальных животных при адаптации к высокогорью в сочетании с воздействием холода.
При проведении экспериментального исследования было использовано 50 белых беспородных лабораторных крыс-самцов с массой тела 200–300 г.
-
1 группа (контроль) — 30 крыс-самцов, которые находились в низкогорье (г. Бишкек) в те же сроки (исходные данные, 3 и 30 сутки).
-
2 группа — крысы, пребывающие в высокогорье 3 суток в сочетании с холодовым воздействием;
-
3 группа — крысы, пребывающие в высокогорье 30 суток в сочетании с холодовым воздействием.
Первый этап эксперимента (контрольная группа) проводился в весеннее время в условиях низкогорья в г. Бишкек. Холодовое воздействие моделировали следующим образом: экспериментальные животные на время эксперимента содержались в помещении при температуре 4–6°С в металлических клетках с небольшим количеством подстилки из опилок. Корм и вода давались им в избытке. Вторая и третья серии эксперимента проводились в условиях пребывания животных на высокогорной базе Туя-Ашу в осеннее время в не термостатированных помещениях вивария при температурах 4–6°С в течение 30 дней. Доступ к корму и воде не ограничивался. Забор крови осуществлялся на 3 и 30 сутки пребывания в низкогорье и высокогорье.
Для электронно-микроскопического исследования использовали свежую кровь, стабилизированную гепарином, которую центрифугировали при 1000 об/мин в течение 10 минут. Выделенная лейкоцитарная пленка обрабатывалась по общепринятой методике. Фиксацию материала производили немедленно, 2,5% глутаральдегидом на буфере Миллонинга, дофиксацию проводили с использованием 1% раствора тетраоксида осмия (все используемые реактивы фирмы Sigma, США). После обезвоживания препараты заливали в эпон (Fluka, Швейцария) по общепринятой методике [5–7].
Срезы, приготовленные на ультратоме, после контрастирования 2% водным раствором уранилацетата натрия или цитратом свинца, приготовленным по Reinolds [8] просматривались с помощью трансмиссионного электронного микроскопа ПЭМ-100 (JEOL, Япония, 2007). Морфометрический анализ ультраструктуры лейкоцитов проводили по Э. Р. Вейбелю [9–11].
Полученные данные обрабатывались при помощи персонального компьютера с использованием табличного редактора Excel 2016 с помощью пакетов прикладных программ Statistica 6.0.
Статистическую значимость (достоверность) различий по количественным переменным определяли путем вычисления t-критерия Стьюдента при параметрическом распределении данных. Изменения считались статистически значимыми (достоверными) при P<0,05 [12–14].
Результаты и обсуждение
Результаты морфологического исследования лейкоцитов в условиях высокогорья в сочетании с холодом отражены в Таблице.
Ранее было показано, что у крыс при адаптации к условиям высокогорья показатели объемной плотности митохондрий к третьему дню адаптации значимо увеличивались в нейтрофилах, моноцитах и эозинофилах [15].
Таблица
МОРФОМЕТРИЧЕСКИЕ ПОКАЗАТЕЛИ УЛЬТРАСТРУКТУРЫ КЛЕТОК КРОВИ КРЫС ПРИ АДАПТАЦИИ К УСЛОВИЯМ ВЫСОКОГОРЬЯ В СОЧЕТАНИИ
С ХОЛОДОМ ПРИ t +4°С (M±m, объемная плотность в %)
|
Показатели |
Контрольная группа 1 г |
Сроки адаптации к высокогорью |
|
|
руппа (3 сутки) |
2 группа (30 сутки) |
||
|
Нейтрофилы |
|||
|
Митохондрии |
0,6±0,01 |
1,2±0,01* |
1,7±0,01**# |
|
Рибосомы |
0,9±0,01 |
1,6±0,01* |
2,6±0,01**# |
|
Вакуоли |
0,4±0,01 |
4,3±0,01* |
1,4±0,01**# |
|
Лизосомы |
7,1±0,01 |
8,9±0,01* |
9,6±0,01**# |
|
Спец. гранулы |
21,3±0,01 |
13,1±0,2* |
21,2±0,4# |
|
Эозинофилы |
|||
|
Митохондрии |
1,1±0,01 |
1,6±0,01* |
1,9±0,01**# |
|
Рибосомы |
1,3±0,01 |
1,3±0,01* |
2,9±0,01**# |
|
Вакуоли |
0,3±0,01 |
1,7±0,01* |
0,8±0,01**# |
|
Спец. гранулы |
21±0,3 |
10,0±0,3* |
21,0±0,4# |
|
Моноциты |
|||
|
Митохондрии |
2,6±0,01 |
3,6±0,3* |
5,2±0,2**# |
|
Рибосомы |
2,3±0,01 |
2,8±0,1* |
4,6±0,4**# |
|
Вакуоли |
0,6±0,01 |
1,3±0,02* |
0,8±0,01**# |
|
Лизосомы |
12,1±1,7 |
14,0±0,2* |
17±0,03**# |
|
Средние лимфоциты |
|||
|
Митохондрии |
1,9±0,01 |
2,8±0,01* |
4,7±0,02**# |
|
Рибосомы |
1,6±0,01 |
1,6±0,04 |
5,2±0,01**# |
|
Вакуоли |
0,4±0,01 |
1,2±0,01* |
0,6±0,01**# |
|
Лизосомы |
0,9±0,02 |
0,7±0,01* |
0,9±0,01# |
|
Примечание: * — |
р < 0,05 при сравнении 1 группы с |
контролем; ** — |
р < 0,05 при сравнении |
2 группы с контролем; # — р < 0,05 при сравнении 2 группы с 1 группой.
Такая же тенденция наблюдалась и у крыс при адаптации к условиям высокогорья с сочетанием холодом: на 3 сутки уровень объемной плотности митохондрий повышался почти в 2 раза в нейтрофилах, в 1,5 в эозинофилах и в 1,4 раза в моноцитах и средних лимфоцитах. С увеличением срока пребывания в высокогорье в сочетании с холодовым воздействием объемная плотность митохондрий продолжала возрастать во всех видах клеток белой крови, и на 30-е сутки адаптации была статистически значимо выше и в сравнении с контролем, и в сравнении с показателями группы животных на 3 сутки адаптации. Эти данные еще раз подчеркивают важнейшую роль митохондрий, как одного из основных регуляторов энергетического обмена в ответе организма на действие стрессовых факторов.
Анализ показателей морфометрии клеток белой крови выявил также статистически значимое увеличение объемной плотности рибосом на 3 сутки адаптации к высокогорью в сочетании с холодом в нейтрофилах, эозинофилах и лимфоцитах (Рисунок 1).
Так, показатели объемной плотности рибосом возросли от 0,9±0,014 до 1,6±0,017 (р < 0,01) в нейтрофилах и от 2,3±0,011 до 2,8±0,1 (р < 0,05) в моноцитах. Причем, на 30 сутки адаптации к высокогорной гипоксии в сочетании с воздействием холода, объемная плотность рибосом продолжал возрастать: в нейтрофилах с 0,9±0,014 до 2,6±0,017 (р < 0,001), в эозинофилах от 1,3±0,014 до 2,9±0,011 (р < 0,01), в моноцитах от 2,3±0,011 до 4,6±0,4 (р < 0,01), в лимфоцитах от 1,6±0,011 до 5,2±0,01 (р < 0,001).
Такая же тенденция к увеличению объемной плотности была отмечена и у вакуолей клеток (Рисунок 2).
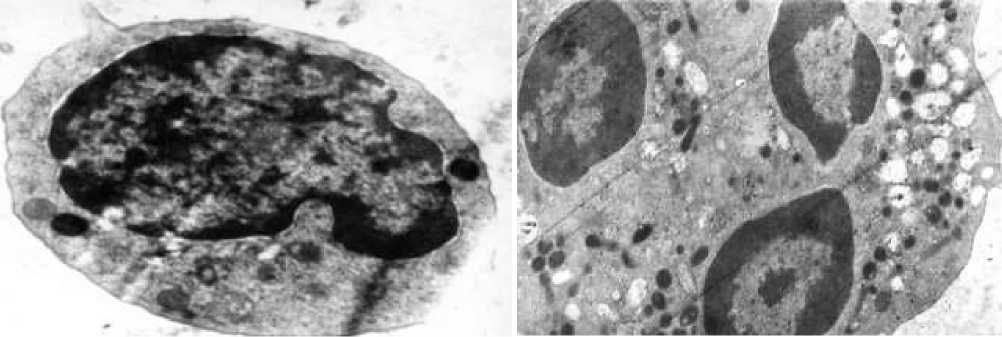
Рисунок 1. Средний лимфоцит крысы на 30 сутки адаптации в сочетании с холодом. В цитоплазме видны митохондрии и большое количество рибосом. Ядро с конденсированным и диффузным хроматином. Ув. ×10000
Рисунок 2. Нейтрофильный лейкоцит крысы на 3 сутки адаптации к условиям высокогорья в сочетании с холодом. В цитоплазме среди первичных и вторичных гранул видны многочисленные электроннопрозрачные вакуоли. Ув. ×20000
В этом случае на 3 сутки адаптации было зафиксировано возрастание показателя объемной плотности вакуолей по сравнению с контролем у нейтрофилов с 0,4±0,013 до 4,6±0,011 при р <0,05, в эозинофилах — с 0,3±0,011 до1,3±0,011 при р <0,05, в моноцитах с 0,6±0,013 до 0,9±0,011 при р <0,05. Повышение объемной плотности вакуолей объясняется тем, что идет процесс увеличения деления клеток и в связи с этим идет увеличение снабжения клеточной мембраны липидами, высвобождение различных соединений из клетки. Это могут быть гормоны или нейромедиаторы, белки и другие макромолекулы. К 30 дню адаптации объемная плотность вакуолей по сравнению с животными на 3 день адаптации статистически значимо снижалась, хотя так и не достигала показателей животных контрольной группы.
Динамика морфометрических показателей лизосом была следующей: в нейтрофилах и моноцитах объемная плотность лизосом к 3 суткам увеличивалась и продолжала возрастать к 30 суткам адаптации. В лимфоцитах этот показатель к 3 дню адаптации уменьшился с 0,9±0,017 до 0,8±0,014 при р <0,05, а к 30 дню адаптации сравнялся с показателями животных контрольной группы.
Особенно выраженные изменения при адаптации к условиям высокогорья в сочетании холодом были выявлены в отношении специфических гранул. Так, на 3 сутки наблюдения объемная плотность специфических гранул нейтрофилов по сравнению с контролем снизилась почти на 100% — с 21,3±0,014 до 13,1±0,2 при р <0,001, а у эозинофилов от 21±0,3 до 10,0±0,3 при р <0,001. Как известно, специфические гранулы содержат белки, которые обладают бактерицидным свойством.
Также в специфических гранулах содержится фермент NADPH-оксидаза. Этот фермент регулирует образование активных форм кислорода, за счет которых и реализуется бактерицидная функция специфических гранул. Снижение объемной плотности специфических гранул свидетельствует о значительном снижении сопротивляемости организма в период аварийной адаптации к гипоксии и холоду. К 30 суткам пребывания в условиях высокогорья в сочетании с холодом показатель объемной плотности специфических гранул в нейтрофилах и эозинофилах практически сравнялся с показателями животных контрольной группы.
Результаты проведенных исследований позволяют заключить, что морфологическим отражением функциональной активности клеток белой крови под воздействием факторов высокогорья в сочетании с холодовым воздействием являются повышение объемной плотности митохондрий, рибосом и вакуолей и снижение объемной плотности специфических гранул.
Список литературы Влияние экстремальных факторов высокогорья на ультраструктуру лейкоцитов крови крыс
- Colguhoun C. M. Introversion - extroversion and the adaptation of the body-temperature rhythm to high work // 5-th International Symposium on high and shift work (Rouen, 12-16 may, 1980). Chronobiologia, vol. VII, July-September 1980. Р. 428.
- Власов Ю. А. Кровообращение и газообмен человека. Новосибирск, 1983. 208 с.
- Агаджанян А. Адаптация человека к условиям Арктического Заполярья. М., 1994. 156 с.
- Казначеев В. П. Клинические аспекты полярной медицины. М.: Медицина, 1986. 208 с.
- Карупу В. Я. Электронная микроскопия. Киев: Вища школа, 1984. 208 с.
- Юрина Н. А. Гистология. М.: Медицина, 1995. 256 с.
- Мавликеев М. О. Краткий курс гистологической техники. Казань, 2020. 107 с.
- Уикли Б. Электронная микроскопия для начинающих. М.: Мир, 1975. 326 с.
- Ташке К. Введение в количественную цито-гистологическую морфологию. Бухарест, 1980. 192 с.
- Wu D. X. Lung lesions in experimental hydrostatic pulmonary edema: an electron microscopic and morphometric study // Exp. Lung Res. 1995. V. 21. №5. P. 711-730.
- Вейбель Э. Р. Морфометрия легких человека. М.: Медицина, 1970. 170 с.
- Реброва О. Ю. Статистический анализ медицинских данных. Применение пакета прикладных программ STATISTICA. М., Медиасфера. 2006. 312 с.
- Боровиков В. П. STATISTICA. Искусство анализа данных на компьютере: Для профессионалов. СПб.: Питер, 2003. С. 146-184.
- Гублер Е. В. Вычислительные методы анализа и распознавания патологических процессов. Л., 1978. 296 с.
- Бектурганова А. О. Морфофункциональное состояние лейкоцитов при кратковременной адаптации животных к климатогеографическим условиям высокогорья // Академический журнал Западной Сибири. 2023. Т. 19. №1. С. 49-52.
