Влияние элементов питания на накопление и жирнокислотный состав липидов семян тыквы

Автор: Надежкин С.м, Голубкина Н.А., Киселева Т.В.
Журнал: Овощи России @vegetables
Рубрика: Проблемы качества овощной продукции. Овощи - здоровье нации
Статья в выпуске: 3 (20), 2013 года.
Бесплатный доступ
Изучено влияние органических и минеральных удобрений на накопление липидов семенами тыквы крупноплодной и жирнокислотный состав липидов. Установлено влияние характера питания на размер семян, содержание липидов и полиненасыщенных жирных кислот.
Тыква крупноплодная, семена, тыквенное масло, липиды, жирные кислоты, минеральные и органические удобрения
Короткий адрес: https://sciup.org/14025032
IDR: 14025032 | УДК: 631.531:635.621:581.19
Effect of fertilizer elements on lipids accumulation and fatty acids composition of pumpkin seeds
Effect of organic and mineral fertilizers on pumpkin seeds lipids accumulation and their fatty acids composition is investigated. The influence of nutrition's composition on the seeds size, lipids content and concentration of polyunsaturated fatty acids was shown.
Текст научной статьи Влияние элементов питания на накопление и жирнокислотный состав липидов семян тыквы
Семена тыквы известны своей пищевой ценностью и широким спектром биологического действия. Они содержат значительные количества эссенциальных жирных кислот (до 50%), белки (около 30%), сахара, фитостерины, витамины группы B, аскорбиновую кислоту, фитин, лецитин, каротиноиды, тирозин, салициловую кислоту, камедь и микроэлемент цинк (Rezig et al, 2012; Jariene et al, 2007). В ряде стран семена составляют традиционные компоненты салатов, заправок для супа, добавок в кондитерские изделия.
В медицине семена тыквы используют как эффективное средство против гельминтов, для лечения гипертонии, гиперплазии предстательной железы и воспаления мочевого пузыря (Mafakheri et al., 2004), а также для лечения рожистого воспаления (Messiaen CM, Fagbayide, 2004). Высокое потребление семян тыквы препятствует образованию почечных камней, снижает риск развития рака желудочно-кишечного тракта, груди, легких и прямой кишки (Huang et al, 2004).
Наибольший интерес привлекает тыквенное масло (Lazos, 1986; Elmadfa&Fritzsche, 1999), главными жирными кислотами которого, составляющими около 90% от общего количества, являются олеиновая, пальмитиновая, стеариновая и линолевая (Tsaknis et al, 1997; Glew et al, 2006). Тыквенное масло содержит также омега-3-жирные кислоты; последние помогают предотвратить атеросклероз, гипертонию и сердечно-сосудистые заболевания, а также стимулируют метаболизм жиров.
На уровень накопления липидов и жирнокислотный состав оказывают влияние различные факторы, включая вид (Mi Young Kim et al, 2012) и сорт тыквы (C^erniauskiene', 2011), климатические условия выращивания (C^ erniauskiene' , 2011), количество вносимых минеральных удобрений (Oloyede et al, 2012) и технологию выращивания (Bahrami et al, 2009).
Установлена отрицательная корреляция между содержанием в семенах липидов и белка (r=-0,997, P<0,01), а также липидов и суммой активных температур в месяце (C^ erniauskiene' , 2011), что непосредственно связано с замедлением процесса фотосинтеза и продукции белков при снижении количества поступающей солнечной энергии с одновременным возрастанием количества жиров (Mailer, Cornish,1987). Таким образом, тесно связанные конкурирующие процессы синтеза жиров из глицерина и жирных кислот и синтез белков из аминокислот имеют место при созревании семян. Кроме того, в период созревания семян синтез жиров обычно требует больше воды, чем синтез белков, поэтому плохой полив в этот период снижает уровень липидов (Bahrami et al, 2009). Отмечается, что в условиях невысоких температур семена тыквы содержат больше липидов с большим процентом олеиновой кислоты и стерола, однако, масса семян в таких условиях уменьшается. Увеличить количество липидов в семенах, а также массу семян можно, используя рассадный способ и технологию прищипывания молодых побегов (Bahrami et al,2009). Однако в этой работе авторы не исследовали жирнокислотный состав липидов. Наконец, установлено, что микроэлементный состав семян тыквы является четким отражением места произрастания тыквы, однако не оказывающего влияния на количество масла (Bandoniene et al, 2013).
Среди факторов, влияющих на жирнокислотный состав липидов тыквы, следует выделить температуру и доступность влаги. Литературные данные свидетельствуют о том, что соотношение между насыщенными и ненасыщенными жирными кислотами зависит от климата. В северных райо- нах липиды семян тыквы содержит больше ненасыщенных жирных кислот, что является проявлением защитной реакции растения к низким температурам. В самом деле, обнаружена прямая корреляция между количеством насыщенных жирных кислот и суммой активных температур в течение вегетационного периода (r=+0,752, P<0,01) (Mailer, Cornish,1987). Отрицательная корреляция установлена между количеством осадков за вегетационный период и количеством ненасыщенных жирных кислот.
Данные влияния особенностей питания растений на количество липидов семян тыквы и их жирнокислотный состав весьма фрагментарны. Так, установлено, что высокие дозы минеральных удобрений (более 100 кгNPK/га) снижают выход липидов (Oloyede et al, 2012).
Целью настоящей работы было исследование влияния последействия использования минеральных и органических удобрений на накопление липидов семян тыквы и их жирнокислотный состав.
Материалы и методы
Тыкву крупноплодную сорта Россиянка выращивали на опытном поле ВНИИССОК в Московской области. Схема опыта: (3х2)х4 со следующими факторами и градациями: фактор А – последействие органических удобрений: 1 – без удобрений (контроль); 2 – навоз полуперепревший (15 т/га на 1 га севооборотной пашни); 3 – зеленая масса пласта козлятника (57,2 т сырой или 9,9 т/га сухой массы); фактор В – применение минеральных удобрений: 1 – без удобрений, 2 – N60P45K60. Минеральные удобрения вносили в виде аммиачной селитры, двойного суперфосфата и хлористого калия, повторность в опыте четырехкратная, варианты размещались рендомизированно. Посев тыквы проведен по схеме 70х70 см. Общая площадь делянок – 28 м2, учетная – 20 м2. Учет урожая – сплошной поделяночный.
Органические удобрения были заделаны в паровом поле в 2009 году. Опыт заложен на дерново-подзолистой почве, пахотный слой которой имел в 2009 году следующую агрохимическую характеристику: содержание гумуса 2,05 %, Р205 – 450 мг/кг, К20 – 357, N щелочногидролизуемый – 108 мг/кг почвы, рН – 6,8, сумма обменных оснований – 18,9 мг-экв, гидролитическая кислотность – 0,96 мг-

Количество элементов питания, внесенных с органическими и минеральными удобрениями суммарно за 4 года составило (кг/га):
Для оценки биохимических показателей семян отбирали по 5 плодов после созревания с каждого варианта, семена выделяли, подсушивали, кожуру отделяли, взвешивали, ядро экстрагировали смесью хлороформ – метанол, 10:1, растворитель удаляли и высушивали до постоянной массы при 50°C. Гомогенизированные образцы хранили до начала анализа в холодильнике.
Определение жирнокислотного состава липидов семян осуществляли по времени выхода соответствующих метиловых эфиров с капиллярной стеклянной колонки газового хроматографа Carlo Erba Strumentazione при температурной программе хроматографа: 80 °C – 10 минут, 175 °C – 15 минут, 200 °C – 5 минут, 225 °C – 70 минут. Температура инжектора – 260 °C, детектора – 240 °C.
Содержание селена устанавливали флуорометрически (Alfthan, 1984), используя мокрое сжигание образцов смесью азотной и хлорной кислот, восстановление шестивалентного селена до Se+4 действием 6Н соляной кислоты и конденсацию образующейся селенистой кислоты с 2,3-диаминонафталином.
Статистическую обработку результатов осуществляли с использованием критерия Стьюдента.
Результаты исследований
В таблице 1 приведены результаты определения биометрических показателей при выращивании тыквы с использованием органических и минеральных удобрений. Доля семян в плодах тыквы варьировала от 0,93 до 6,58%. Семена сильно различались по размеру: показатель массы 1000 семян находился в интервале от 206 до 378 г. Содержание липидов в семенах было наибольшим (более 40%) в варианте с использованием навоза, однако, с учетом общей массы семян с плода наибольший выход липидов установлен при выращивании тыквы с применением в качестве сидерата – козлятника восточного с использованием минеральных удобрений (около 1% с одного плода). Варианты с навозом и сидератом козлятника отличались меньшей долей выполненных семян (39-45 %) (табл.1).
Соотношение массы плаценты к массе семян уменьшалось в ряду: минеральные удобрения > органические удобрения (навоз и козлятник) > совместное использование минеральных и органических удобрений (рис.1).
Между содержанием липидов в семенах тыквы и долей ядра (от общей массы семени) наблюдалась обратная корреляция (коэффициент корреляции 0,80, P<0,005) (рис.2).
Взаимосвязь величины накопления липидов семенами и уровня аккумулирования природного антиоксиданта селена описывалась полиномиальным уравнением: Y=-0,016X2 + 1,846X – 10,41, где Y – содержание липидов %, Х – концентрация селена, причем, во всех вариантах ис-
1. Выход липидов при выращивании тыквы с использованием органических и минеральных удобрений
|
Вариант |
Урожай семян, % от массы плода |
% Выполненных семян |
М 1000 семян, г |
Доля ядра в семени % |
Содержание липидов % |
Выход масла с 1 плода % |
|
1 |
1,68 |
88 |
239 |
72,2 |
38,9 |
0,376 |
|
2 |
2,07 |
97,2 |
228 |
73,1 |
36,85 |
0,151 |
|
3 |
1,3 |
45,1 |
206 |
71,7 |
40,5 |
0,126 |
|
4 |
3,34 |
100 |
227 |
76,0 |
37,7 |
0,265 |
|
5 |
0,93 |
38,8 |
359 |
76,9 |
36,1 |
0,297 |
|
6 |
6,58 |
100 |
378 |
71,8 |
39,3 |
0,977 |
|
научно- |
практический журнал |
[ 45 ] |
овощи |
россии № 3 (20) |
2013 |
|
-
2. Жирнокислотный состав масла семян тыквы (% от суммы)
Жирная кислота
Вариант опыта
1
2
3
4
5
6
С14:0
0,42
0,22
0,33
0,08
0,2
0,24
С16:0
16,95
20,46
23,77
12,20
22,60
19,93
С18:0
6,40
7,81
6,44
5,27
5,60
7,47
С18:1
18,33
21,55
16,95
14,90
15,70
21,21
С18:2
55,47
48,25
50,62
66,36
54,34
49,12
С18:3
0,82
0,78
0,86
0,59
0,69
0,97
С17:0
0,55
0,39
0,43
0,28
0,42
0,41
С20:2
0,35
0,14
0,20
С20:0
0,28
0,34
0,21
0,18
0,18
0,20
С16:1
0,23
0,21
0,26
0,14
0,27
0,24
С20:1
0,20
Следы
Следы
Следы
Следы
Следы
НЖК*
24,60
29,22
31,18
18,01
29,00
28,25
МНЖК**
18,76
21,76
17,21
15,04
15,97
21,45
ПНЖК***
56,64
49,03
51,62
66,95
55,03
50,29
ННЖК****
75,4
70,79
68,83
81,99
71,00
71,74
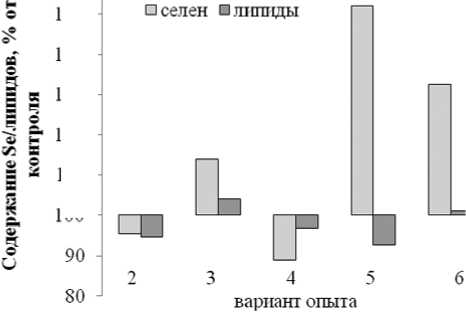
*НЖК – насыщенные жирные кислоты, **МНЖК – мононенасыщенные жирные кислоты, ***ПНЖК – полиненасыщенные жирные кислоты, ****ННЖК – ненасыщенные жирные кислоты пользование NPK – удобрений снижало уровень аккумулирования микроэлемента (рис.3).
Нормированные результаты уровня аккумулирова- ния семенами липидов и селена относительно контроля указывают на наибольшее накопление микроэле- мента в варианте с использованием в качестве орга- нического удобрения
козлятника (варианты
5,6)(рис.4).
Среди жирных кислот липидов семян тыквы лидируют по содержанию олеиновая (С 18:2; 48-66%), пальмитиновая (C16:0; 12-24%), линолевая (C18:1; 1522%) и стеариновая (C 18:0; 5-9%) (табл.2). Количество омега-3-жирной кислоты (С18:3) не превышало 0,61% (табл.2). Наибольшее количество насыщенных жирных кислот установлено в семенах, полученных при использовании навоза (более 30 %, вариант 3).
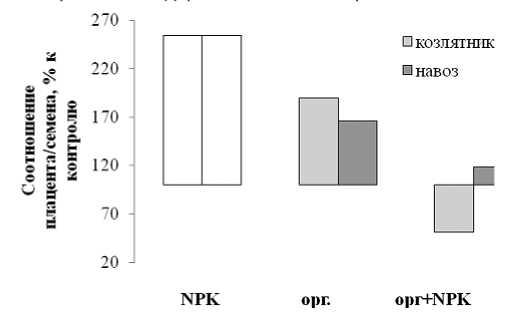
Соотношение массы плацента/семена в
Рис.1.
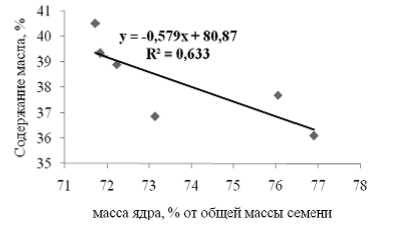
Рис. 2. Взаимосвязь между содержанием липидов и долей ядра в общей массе семени
зависимости от использования органических и минеральных удобрений
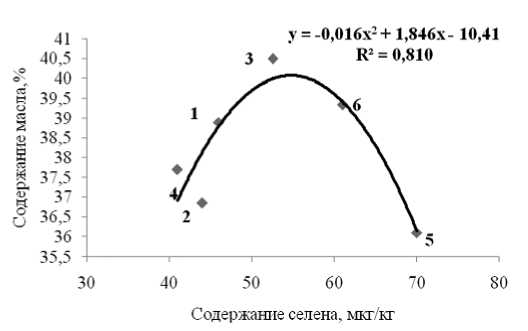
Рис. 3. Зависимость содержания липидов от уровня аккумулирования селена селенами тыквы
160 л
150 -
140 -
130 -
120 -
НО -
100 -
Рис.4. Содержание селена и липидов в семенах тыквы
Наибольший уровень полиненасыщенных жирных кислот был характерен для семян растений, выращенных с использованием навоза и NPK (вариант 4) (табл. 2).
Нормированные показатели содержания жирных кислот относительно данных для контрольных растений представлены на рис.5, из которого видно, что раздельное использование минеральных и органических удобрений приводит к возрастанию содержания пальмитиновой кислоты. Совместное действие навоза и туков обеспечивало увеличение интенсивности синтеза олеиновой кислоты, а применение козлятника и NPK – (рис.5) приводило к возрастанию содержания омега-3-жирных кислот.
Обсуждение результатов

Выявление закономерностей аккумулирования липидов семенами тыквы имеет первостепенное значение для прогнозирования и эффективного регулирования качества урожая.
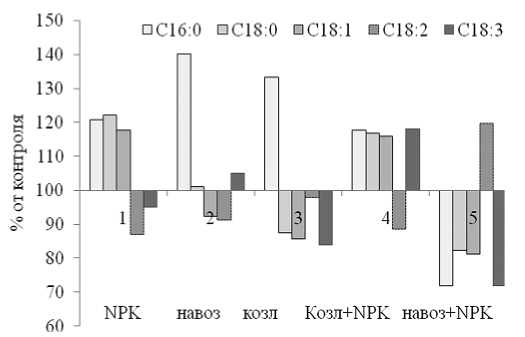
Рис. 5. Нормированные показатели содержания жирных кислот в липидах тыквы
Биометрическиепоказатели
Использование органических и минеральных удобрений при выращивании крупноплодной тыквы сорта Россиянка позволило установить, что характер питания влияет на количество и качество семян, а также соотношение веса плаценты к весу семян. Данные рис.1 свидетельствуют о том, что применение минеральных удобрений более чем в два раза увеличивает соотношение массы плаценты к массе семян. Влияние органических удобрений менее выражено, однако также имеет место возрастание исследуемого показателя, причем для применения козлятника эффект более интенсивен, чем при использовании навоза (1,9 по сравнению с 1,6 раз). Использование сидерата козлятника с NPK-удобрениями достоверно уменьшало соотношение массы плаценты к массе семян по сравнению с контролем, а при использовании навоза с ми- неральными удобрениями соотношение выбранных показателей лишь не на много отличалось от контроля. В связи с этим следует отметить, что максимальная выполненность семян выявлена именно для совместного использования минеральных и органических удобрений и именно для этих вариантов урожай семян был максимален (табл.1). При сравнении урожая семян разных вариантов обращает внимание, как разная величина семян, так и значимое различие в доле ядра. Наиболее крупные семена были при использовании козлятника как раздельно, так и с минеральными удобрениями, наиболее мелкие – при использовании навоза.
Как видно из данных рис.2, содержание липидов в семенах определяется не общей массой семени, а соотношением массы ядра к массе оболочки. Таким образом, оказалось, что лидировали по содержанию липидов самые маленькие семена (вариант 3) и самые большие (вариант 6), для которых доля ядра была наименьшей (71,7-71,8%).
Жирнокислотный состав
Данные таблицы 2 свидетельствуют о том, что использование органических и минеральных удобрений может существенно влиять на жирнокислотный состав масла семян тыквы. Обращает внимание перспективность совместного применения навоза и минеральных удобрений, позволяющее достоверно увеличить содержание в липидах ненасыщенных жирных кислот. Из таблицы 2 и рисунка 5 видно, что влияние совместного использования органических и минеральных удобрений не является арифметической суммой влияния на жирнокислотный состав липидов тыквы отдельных элементов питания. Особенно важным с позиций практики представляется факт большего накопления насыщенных жирных кислот при раздельном применении NPK-удобрений, навоза или сидерата козлятника.

Селен
Принимая во внимание антиоксидантные антистрессовые свойства микроэлемента селена (Голубкина, Папазян, 2006), представляло интерес установить возможность взаимосвязи между уровнем аккумулирования микроэлемента семенами и количеством образующихся липидов и жирнокислотным составом последних. Ранее была показана прямая корреляция между содержанием селена и масла в семенах льна, а также установлено, что обогащение растений селенатом натрия достоверно снижает уровень насыщенных жирных кислот в льняном масле (Голубкина и др, 2012). Для тыквы, однако, взаимосвязь между содержанием селена и масла в семенах оказалась не линейной, а описывалась квадратным уравнением (рис.2), таким образом, что максимальное содержание липидов соответствовало не максимальным, а средним показателям концентрации микроэлемента. Нормирование полученных показателей относительно данных для контрольного варианта (рис.5) позволило установить, что уровень аккумулирования селена семенами регулируется, прежде всего, содержанием его в почве и использованием минеральных удобрений. На рисунке 3 видно, что минеральные удобрения во всех случаях снижают уровень селена в семенах, что хорошо согласуется с известными литературными данными (Голубкина, Папазян, 2006). Максимальное количество селена в семенах наблюдается при использовании в качестве сидерата козлятника, что и следовало ожидать, поскольку бобовые культуры известны своей способностью накапливать более высокие концентрации микроэлемента, чем большинство других сельскохозяйственных растений (Голубкина, Папазян, 2006). Несоответствие полученных результатов с описанными закономерностями для семян льна (Голубкина и др., 2011) скорее всего, связано с тем, что в семенах льна липиды образуют комплекс с белками, в том числе с селеносодержащими. Для семян тыквы существование такого комплекса неизвестно и, согласно литературным данным, отмечается отрицательная корреляция между концентрациями белка и масла (C^ erniauskiene’ , 2011). В отличие от содержания селена в семенах льна (Голубкина и др., 2012), в семенах тыквы отсутствует взаимосвязь между концентрацией селена и жирнокислотным составом масла.
Заключение и выводы
Таким образом, выявлено, что жирнокислотный состав липидов семян тыквы крупноплодной зависит от способа использования органических и минеральных удобрений – раздельно или совместно, что позволяет изменять соотношение насыщенных и ненасыщенных жирных кислот. Показано, что содержание липидов обратно пропорционально доле ядра в семенах. Соотношение масс плацента/семена зависит от раздельного или совместного использования органических и минеральных удобрений.
Список литературы Влияние элементов питания на накопление и жирнокислотный состав липидов семян тыквы
- Curcic Z., Nagal N., et al. Evaluation of genetic diversity among open-pollinated sugar beet populations using quantitative root traits.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.84.
- Del Piano L., Dato F., et al. Genetic variability in N.tabacum as revealed by morphological traits and molecular markers.//Prebreeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.77.
- Gailite A., Bumane S., et al. Evaluation, characterization and utilization of Latvian pea (Pisum sativum L.) genetic resources.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.100.
- Glogovac S., Takac A., et al. Molecular evaluation of tomato (Lycopersicon esculentum L.) genotypes using microsatellite markers.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.78.
- Hegay S., Geleta M., et al. Introducing host plant resistance to seed-borne diseases anthracnose (Colletotricum lindemuthianum) and bean common mosaic virus (BCMV) in Kyrgyz common bean through marker-aided and inoculation-based back-crossing.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.83.
- Jorgensen R., Frenck G., et al. Adaptation of oilseed rape, barley, and Brassica rapa to the future climate and increased CO2.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.17.
- Khazaei H., Street K., et al. The focused identification of germplasm strategy (FIGS): an approach to pre-breeding for water-limited environments.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.42.
- Lebeda A., Kristkova E., et al. Prickly lettuce enormous source of variation unexploited in lettuce breeding.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.87.
- Mba C., Guimaraes E., et al. Enhancing global capacity for pre-breeding.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.24.
- McCouch S., Wright M., et al. Genome wide association studies (GWAS) help develop blueprint for better utilization natural variation in rice.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.15.
- Orabi J., Starr G., et al. A large scale association study for aroma wheat components in bread wheat.//Pre-breeding fish ing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.28.
- Pietrusinska A., Czembor J. Wild wheat as source of resistance to leaf rust and powdery mildew.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.145.
- Poulsen G., Maggioni L., et al. Molecular markers for botanic identification among Brassica species.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.73.
- Sasnauskas A., Gelvonauskiene D. Assessment of resistance to apple scab (Venturia inaequalis) of apple genetic resources and breeding applications at the institute of horticulture, LRCAF.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.88.
- Sumalan R., Popescu S. et al. Studies on biodiversity of local onion (Allium cepa L.) landraces cultivated in saline areas from western Romania through phenotypic and molecular analysis.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.79.
- Suprunova T., Pishnaya O. et al. Practical use of molecular markers of pungency in breeding program of pepper (Capsicum annuum L.).//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.67.
- Svirshchevskaya A., Malysheva O. et al. SSR analysis of sugar beet (Beta vulgaris L.) parental forms of different origin for predicting possible heterosis effects.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.72.
- Terzic S., Dedic B. Interspecific sunflower progenies selection for resistance white rot caused by Sclerotinia sclerotiorum (Lib.) de Bary.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.132.
- Toker C., Ikten F. et al. Association between heterosis and genetic distance based on morphological traits and SSR markers in Cicer species.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.85.
- Uzun B., Yol E. et al. Screening, testing, and molecular diagnosis for resistance against phyllody diseases in sesame.//Prebreeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.82.
- Vilanova S., Gramazio P. et al. Facilitating the fishing in the gene pool of the wild eggplant relative Solanum incanum by developing introgression lines.//Pre-breeding fishing in the gene pool». Abstract of oral presentation and posters of the European Plant Genetic Resources Conference 2013, NordGene, SLU, Alnarp, Sweden. Pp.128.
