Влияние низких концентраций аденозина и гистамина на сокращения мышцы трахеи и бронхов

Автор: Смирнова Любовь Евгеньевна, Кирилина Валентина Михайловна, Федин Анатолий Николаевич, Кривченко Александр Иванович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 4 (149), 2015 года.
Бесплатный доступ
Аденозин и гистамин являются важными биологически активными соединениями для клеточных структур нижних дыхательных путей. В настоящее время недостаточно изучено действие аденозина и гистамина на структуры нижних дыхательных путей в физиологически низких концентрациях, соответствующих естественному фоновому уровню организма. Нашей задачей было изучить влияние низких доз аденозина (10 мкг/мл) и гистамина (0,1 мкг/мл и 10 мкг/мл) на активность гладкой мышцы трахеи и бронхов крысы. В исследовании применялась электрическая стимуляция постганглионарных нервов и блокада гистаминовых рецепторов. При влиянии аденозина в низкой концентрации наблюдается больший дилатационный эффект, чем при действии гистамина, что связано с воздействием аденозина на различные структуры. С учетом того, что искусственно введенный в препараты трахеи и бронхов гистамин равен по концентрации эндогенно выработанному лаброцитами гистамину, следует заключить, что основной констрикторный эффект в системе нижних дыхательных путей связан с гистамином и не связан с действием аденозина или иных медиаторов тучных клеток.
Аденозиновые рецепторы, гистаминовые рецепторы, дегрануляция тучных клеток, тахикинины
Короткий адрес: https://sciup.org/14750893
IDR: 14750893 | УДК: 57.054
Adenosine and histamine low concentration influence on muscle contraction in trachea and bronchi
Adenosine and histamine are important and biologically active molecules of the cellular structure of the lower respiratory tract. Currently, the influence of adenosine and histamine on low airways in physiologically low concentrations is not studied. The purpose of this study was to investigate the influence of low doses of adenosine (10 mkg/ml) and histamine (0,1 mkg/ml и 10 mkg/ml) on the activity of smooth muscles of the trachea and bronchi. In the course of our research, we employed both electrical stimulations of the postganglionic nerves and blockages of histamine receptors. The study revealed that adenosine leads to a much greater dilatation effect than histamine. This effect is conditioned by the broader influence of adenosine on different structures causing dilation. We concluded that he main bronchoconstriction effect is associated with the histamine and is not conditioned by the influence of adenosine or other mediators of the mast cells.
Текст научной статьи Влияние низких концентраций аденозина и гистамина на сокращения мышцы трахеи и бронхов
Аденозин и гистамин являются важнейшими медиаторами тучных клеток. С учетом новых биохимических данных следует рассматривать роль этих веществ в сократительной активности гладкой мускулатуры трахеи и бронхов, а соответственно и в патогенезе распространенных обструктивных заболеваний нижних дыхательных путей, к числу которых, безусловно, относятся хроническая обструктивная болезнь легких и бронхиальная астма.
Аденозин синтезируется после активации тучных клеток и является трансмиттером поздней фазы дегрануляции. Также аденозин появляется во внеклеточных пространствах вследствие разрушения клеток. При действии различных по- вреждающих факторов из клеток во внеклеточное пространство высвобождается АТФ. АТФ быстро превращается в АМФ при участии внеклеточной эктонуклеотиддифосфорилазы (экто-апиразы, или CD39). АМФ быстро трансформируется в аденозин при участии экто-5′-нуклеотидазы (CD73) [6]. Внеклеточный аденозин транспортируется обратно в клетки при участии нуклеозидного транспортера, такого как ENT1 [12]. Аденозин является как аутокринной, так и паракринной сигнальной молекулой и взаимодействует с трансмембранными рецепторами 4 подтипов: A1R, A2AR, A2BR и A3R. Соотношение этих рецепторов и локализация на мембранах различных клеток достаточно вариабельны [2], [11].
Наиболее хорошо изученными рецепторами системы GPCR (Gprotein coupled receptors) оказались аденозиновые рецепторы А1, А2 и А3. Рецепторы А1, А2 и А3 взаимодействуют с различными G-белками: рецепторы А1 и А3 – с Gi/o-белком, а рецептор А2 – с Gs-белком. Кроме того, имеются данные, что все аденозиновые рецепторы могут взаимодействовать и с другими G-белками [3], [4]. Рецепторы А1 и А3 являются ингибиторами аденилатциклазы, а рецепторы А2 – стимуляторами. После активации G-белков активируются ферменты и ионные каналы и запускается каскад сложных биохимических превращений [10]. В тучных клетках присутствуют аденозиновые рецепторы А2В и А3, которые, будучи активированными, облегчают дегрануляцию тучных клеток. Использование неселективного антагониста аденозиновых рецепторов теофиллина широко применяется при лечении астмы, хотя механизм его действия на систему GPCR до конца не ясен [4]. Гетерогенность адено-зин-ассоциированных механизмов определяется прежде всего субтипом рецептора. Взаимодействие с рецепторами тучных клеток субтипа A2B приводит к высвобождению гистамина, который ведет к сокращению гладкой мускулатуры [2]. Сформулированы представления об изменении соотношения количества пуринергических рецепторов разных подтипов. В частности, K. Varani et al. показано увеличение уровней транскриптов для A2AR и A3R при снижении A2BR у больных хронической обструктивной болезнью легких [8]. В результате присоединения аденозина в качестве лиганда к рецепторам тучных клеток А1 и А3 активируется мембраносвязанный фермент – фосфолипаза С. В результате присоединения аденозина в качестве лиганда к рецепторам тучных клеток А2 активируется аденилатциклаза. Фосфолипаза С и аденилатциклаза катализируют реакции с образованием соответственно инозитол-1,4,5-трифосфата, 1,2-диацилглицери-на и цАМФ. Инозитол-1,4,5-трифосфат и цАМФ обеспечивают фосфорилирование и активацию Са2+-связывающего белка кальмодулина, мобилизующего Са2+ из эндоплазматического ретикулума клеток в цитоплазму, в присутствии которого при участии цАМФ и 1,2-диацилглицерина активируется протеинкиназа С. Протеинкиназа С осуществляет фосфорилирование и активацию ряда других внутриклеточных ферментов, в частности Са2+-зависимой фосфолипазы А2. При этом за счет Са2+-индуцированного сокращения микротрубочек гранулы «подтягиваются» к плазматической мембране, а 1,2-диацилглицерин, продукты его расщепления (моноацилглицерин, лизофосфатидиловая кислота) и активации фосфолипазы А2 (лизофосфатидилхолин) обусловливают слияние депонирующих гранул тучной клетки со стенкой мембраносвязанных канальцев и цитоплазматической мембраной, через которые медиаторы гранул (первичные) и медиаторы, образующиеся при активации клеток, высвобождаются наружу. Медиаторы быстрой фазы дегрануляции воздействуют на гладкую мускулатуру, вызывая сокращение миоцитов.
Механизм влияния аденозина на рецепторы гладкой мускулатуры отличается от такового в тучных клетках. При присоединении аденозина к А1- и А3-рецепторам миоцитов запускаются механизмы ингибирования аденилатциклазы – фермента, обеспечивающего гидролиз АТФ до цАМФ с высвобождением пирофосфата. Инактивация аденилатциклазы приводит к уменьшению концентрации цАМФ в цитоплазме миоцитов. При низкой концентрации цАМФ не происходит фосфорилирования киназы легких цепей миозина, что является условием активности этого фермента. При одновременном увеличении концентрации ионов кальция в цитоплазме активируется белок цитоплазмы миоцитов – кальмодулин. Активация осуществляется за счет связывания кальмодулина с катионами кальция. Активный комплекс – кальций-кальмодулин – связывается с киназой легких цепей миозина и активирует этот фермент вследствие удаления с ингибиторного участка фермента ингибирующих его активность протеинов и фосфат-анионов. Активная киназа легких цепей миозина фосфорилирует легкую цепь миозина, вследствие чего последняя соединяется с тяжелой цепью миозина, далее миозиновые нити соединяются с нитями актина, образуя акто-миозиновый комплекс и вызывая гладко-мышечное сокращение [1].
Существуют данные о влиянии аденозина на эпителий и неадренэргические нехолинэргичес-кие чувствительные к капсаицину С-волокна. На С-волокна аденозин оказывает возбуждающее действие, вследствие чего волокна синтезируют медиаторы возбуждающего действия – тахикинины. Основным тахикинином является субстанция Р (SP), которая повышает сократительную активность гладкой мускулатуры трахеи и бронхов.
Другой важнейший медиатор тучных клеток, выбрасываемый в большом количестве с самого начала дегрануляции, – гистамин – оказывает прямое действие на гладкую мускулатуру, эпителий, нейроны метасимпатических ганглиев, капсаицин-чувствительные С-волокна и сами тучные клетки. Гистамин действует на данные структуры через гистаминовые рецепторы – Н1 и Н2. Н1-рецепторы в системе нижних дыхательных путей локализованы на мембране постсинаптических нервных клеток, гладкой мускулатуре, эпителии, тучных клетках. Активация этих рецепторов приводит к увеличению продукции цГМФ и повышению концентрации внутриклеточного кальция и развитию сокращения гладкой мускулатуры трахеи и бронхов. Н2-рецепторы имеют представительство на гладкой мускулатуре, эпителии и тучных клетках, восприимчивы к низким концентрациям гистамина и опосредуют расслабление мускулатуры нижних дыхательных путей. Действие ГА через эти рецепторы реализуется при повышении внутриклеточной концентрации цАМФ. Н2-рецепторы при их активации также ингибируют высвобождение серотонина, способствующего сократительной активности, и увеличивают синтез вазоактивного интестинального пептида (VIP), являющегося медиатором неадренэргической нехолинэргической системы, обеспечивающим дилатацию гладкой мускулатуры [7]. Все эти последствия активации Н2-рецепторов определяют их дилатационный эффект.
При влиянии гистамина на эпителий респираторного тракта запускается синтез простагландинов (ПГЕ2 и ПГЕ2а), оказывающих двойственный эффект на миоциты. ПГЕ2 индуцируют снижение сократительной активности, а ПГЕ2а - ее увеличение [5].
Известно, что гистаминергические реакции гладкой мышцы опосредуются через трахеобронхиальные быстро- и медленноадаптирующиеся стретч-рецепторы, расположенные в эпителиальном и мышечном слоях. Возбуждение трехео-бронхиальных стретч-рецепторов приводит к рефлекторному сокращению гладкой мускулатуры дыхательных путей и кашлевому рефлексу [7].
Таким образом, можно отметить, что действие аденозина и гистамина в нижних дыхательных путях оказывается тесно связанным. Влияние аденозина и гистамина на гладкую мышцу трахеи и бронхов опосредуется разными типами гистаминовых и аденозиновых рецепторов, которые могут быть расположены на различных структурах респираторного тракта, в том числе непосредственно на гладкомышечных клетках, чувствительных окончаниях С-волокон, дыхательном эпителии, тучных клетках. Характер реакции гладкой мышцы при этом будет определяться балансом активирующих и тормозных влияний, приходящих от рецепторных полей, локализованных на указанных структурах, и исходным состоянием гладкой мышцы. В настоящее время остаются недостаточно изученными вопросы взаимодействия аденозина и гистамина в низких концентрациях, соответствующих естественному фоновому уровню организма. Нашей задачей было изучить влияние низких доз аденозина (10 мкг/мл) и гистамина (0,1 мкг/мл и 10 мкг/мл) на активность гладкой мышцы трахеи и бронхов, связанную с эфферентными нервными структурами (постганглионарными нервными окончаниями). Также в задачи входило сопоставление эффектов аденозина и гистамина с целью установления роли каждого из этих трансмиттеров. Для этого были изучены реакции мышцы, вызванные электростимуляцией постганглионарных нервов, при действии гистамина и аденозина на фоне действия блокады Н1- и Н2-рецепторов.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектом исследования являлись 20 крыс линии Вистар обоего пола с массой тела 180-250 г. Подготовленные препараты трахеи и бронхов помещались в камеры с физиологическим раствором Кребса - Хензелайта, где поддерживался необходимый уровень кислорода и температурный режим. В ходе эксперимента экзогенно в виде аппликаций вводились следующие вещества: аденозин (С = 10 мкг/мл; V = 0,2 мл) для активации тучных клеток, С-волокон и аденозиновых рецепторов гладкой мышцы; гистамин (0,1 мкг/мл; 10 мкг/мл; V = 0,2 мл) для активации гистаминовых рецепторов. Кроме искусственного введения гистамина в препараты, происходило естественное поступление гистамина в систему вследствие дегрануляции тучных клеток, вызванной аденозином. Концентрации эндогенного и экзогенного гистамина являются равными. Таким образом, сравнивая результаты серии опытов «аденозин + естественно выработанный гистамин» с результатами экспериментов по влиянию на гладкомышечные сокращения одного только гистамина, можно выявить физиологический эффект аденозина и установить его роль в сокращении гладкой мускулатуры трахеи и бронхов крысы. В ходе всех экспериментов применялось раздражение препаратов электрическим полем с помощью стимулятора ЭСЛ-2. Серебряные электроды располагались вдоль продольной стенки ванночки по обе стороны препарата. Применялась стимуляция постганглионарных нервов с частотой 30 стим./с, длительность 0,5 мсек, амплитуда 20 В, продолжительность стимуляции 10 с.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Влияние гистамина на ответы трахеи и бронхов при блокаде Н2-рецепторов
Мы заблокировали тормозные гистаминовые Н2-рецепторы блокатором циметидином. Применение 10 мкг циметидина не изменяло сокращений трахеи и бронхов, вызванных электростимуляцией постганглионарных нервов. Гистамин на фоне блокады Н2-рецепторов вызывал дозозависимое усиление сокращений на бронхах - до 109 ± 2,2 % при 0,1 мкг (P < 0,05). На трахее при дозе 10 мкг повышение амплитуды сокращения достигало 121,2 ± 1,5 % (рис. 1). Ответы трахеи и бронхов имели значимые различия (P < 0,05). Увеличение сокращения в ответ на введенный гистамин является закономерным, поскольку препарат действовал в основном на оставшиеся незаблокированными Н1-рецепторы, опосредующие констрикторный эффект.
Влияние гистамина на ответы трахеи и бронхов при блокаде Н1-рецепторов
Для блокады активирующих гистаминовых рецепторов использовали Н1-антагонист супрастин. Блокада Н1-рецепторов устраняла конс-трикторный эффект, связанный с влиянием на
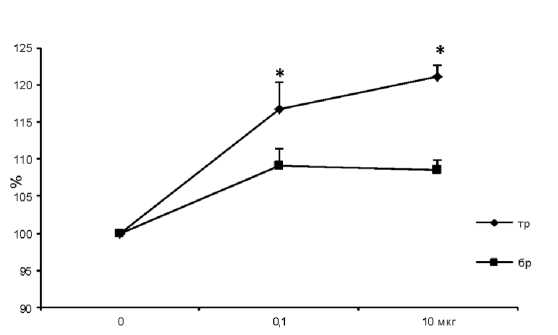
Рис. 1. Действие гистамина на сокращения гладкой мышцы трахеи и бронхов на фоне блокады Н2-рецепторов.
По оси абсцисс – концентрация гистамина в мкг, по оси ординат – изменения ответов в %. За 100 % принимается величина сокращений препаратов на фоне циметидина.
* – достоверные (P < 0,05) отличия эффекта гистамина. Сокращения: «тр» – трахея, «бр» – бронхи эти структуры гистамина, и позволяла выявить его влияние на оставшиеся незаблокированными звенья стенки респираторного тракта (Н2- и Н3-рецепторы). На фоне действия 100 мкг супрастина величина сокращений значимо снижалась (до 84,2 ± 2,5 % на трахее и до 87,6 ± 2,5 % на бронхах (P < 0,05)). Гистамин на фоне блокады Н1-рецепторов также достоверно (P < 0,05) понижал ответы трахеи и бронхов (рис. 2). Влияние гистамина на трахею носило дозозависимый характер (уменьшение ответов до 73,6 ± 3,4 % при 10 мкг), на бронхах дозозависимости не наблюдалось (амплитуда сокращений составляла 91 % при обеих дозах гистамина). Ответы трахеи и бронхов на действие гистамина имели достоверные различия (P < 0,05). Снижение сокращений респираторной мышцы связано с устранением ведущих брон-хоконстрикторных акцепторов к гистамину – Н1-рецепторов и его взаимодействием с рецепторами Н2 и Н3, опосредующими дилатацию.
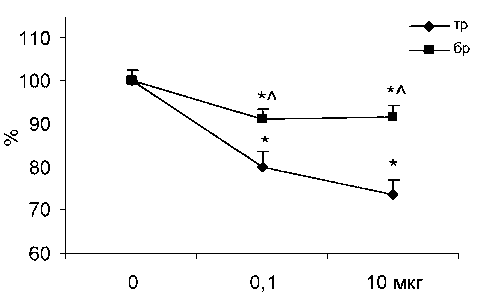
Рис. 2. Действие гистамина на сокращения гладкой мышцы трахеи и бронхов на фоне блокады Н1-рецепторов.
По оси абсцисс – концентрация гистамина в мкг, по оси ординат – изменения ответов в %. За 100 % принимается величина сокращений препаратов на фоне супрастина. * – достоверные (P < 0,05) отличия эффекта гистамина.
л - достоверные (P < 0,05) отличия между ответами препаратов трахеи и бронхов. Сокращения: «тр» – трахея, «бр» – бронхи
Влияние аденозина на ответы трахеи и бронхов при блокаде Н2-рецепторов
Для опытов с аденозином из респираторного тракта избирались препараты трахеи и бронхов с ганглиями (в местах бифуркации трахеи и бронхов) и без них (сегменты вне бифуркаций). Подобное разделение препаратов позволяло изучить влияние аденозина непосредственно на нейроны интрамурального метасимпатического ганглия и установить наличие или отсутствие аденозиновых рецепторов в нейрональных структурах ганглия. При блокаде Н2-рецепторов циметидином на фоне постганглионарной стимуляции аденозин оказывал двухфазный эффект: сначала наблюдалось увеличение сократительных ответов гладкой мышцы, а затем – уменьшение. Ответы трахеи с ганглиями повышались до 108,9 ± 2,4 % (рис. 3), что было достоверно (P < 0,05) выше действия одного аденозина. Ответы трахеи без ганглиев повышались почти на такую же величину (107,1 ± 2,6 %) и превышали (P < 0,05) эффект одного аденозина. Фаза снижения сокращения на этих препаратах равнялась 102,2 ± 3,1 % и 95,1 ± 2,8 % соответственно. При действии аденозина на фоне блокады Н2-рецепторов бронхи с ганглиями и без ганглиев показывали усиление сократительных ответов (116,1 ± 4,9 и 116,3 ± 5,2 % соответственно), которое было больше по величине, чем на трахее. Фаза снижения сокращения на бронхах с ганглиями составляла 96,7 ± 2,8 %, на бронхах без ганглиев - 97,4 ± 2,4 %. Существенных различий между препаратами трахеи и бронхов с ганглиями и без них не наблюдалось, что, вероятно,
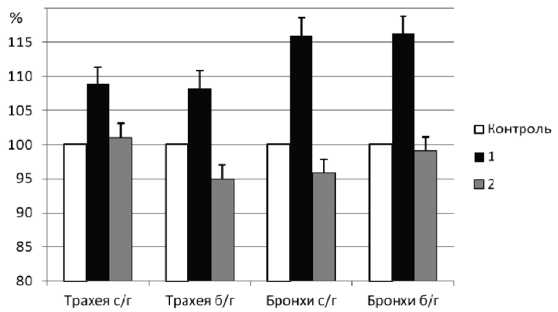
Рис. 3. Влияние блокады гистаминовых Н2-рецепторов циметидином на сократительные ответы гладкой мускулатуры трахеи и бронхов, обработанных аденозином при постганглионарной стимуляции.
По оси абсцисс обозначены используемые препараты. По оси ординат обозначены изменения ответов гладкой мышцы в %. За 100 % приняты ответы гладкой мышцы, полученные при стимуляции постганглионарных нервов на фоне действия аденозина. «Трахея с/г» – трахея с ганглиями. «Трахея б/г» – трахея без ганглиев. «Бронхи с/г» – бронхи с ганглиями. «Бронхи б/г» – бронхи без ганглиев. «1» – первая фаза действия аденозина на фоне блокады Н2-рецепторов циметидином. «2» – вторая фаза действия аденозина на фоне блокады Н2-рецепторов циметидином.
* – достоверное (P < 0,05) отличие от контроля указывает на отсутствие рецепторов к аденозину в нейронах интрамуральных ганглиев.
Влияние аденозина на ответы трахеи и бронхов при блокаде Н1-рецепторов
Блокада гистаминовых Н1-рецепторов супрастином на фоне постганглионарной стимуляции с применением блокады Н2-рецепторов уменьшала сократительные ответы гладкой мышцы всех используемых препаратов на применение аденозина. Ответы трахеи с ганглиями достоверно (P < 0,01) снижались по сравнению с контролем до 43,9 ± 1,9 % (рис. 4). Ответы трахеи без ганглиев снижались почти на такую же величину (42,1 ± 2,2 %). Действие аденозина при блокаде Н1-рецепторов на бронхи с ганглиями оказывало аналогичный эффект на сократительные ответы (42,5 ± 1,9). На бронхах без ганглиев снижение сокращения достигало 43,3 ± 2,1 %. Различий между препаратами с ганглиями и без них не наблюдалось.
%
□ Контроль
■ Супрастин
LILI J J
Трахея с/г Трахея б/г Бронхи с/г Бронхи б/г
Рис. 4. Влияние блокады Н1-рецепторов супрастином на сократительные ответы ГМ трахеи и бронхов, вызванные постганглионарной стимуляцией, на фоне блокады Н2-рецепторов циметидином при аппликации аденозина.
По оси абсцисс обозначены используемые препараты. По оси ординат обозначены изменения ответов гладкой мышцы в %. За 100 % приняты ответы гладкой мышцы, полученные при постганглионарной стимуляции на фоне аппликаций аденозина. «Трахея с/г» – трахея с ганглиями. «Трахея б/г» – трахея без ганглиев. «Бронхи с/г» – бронхи с ганглиями. «Бронхи б/г» – бронхи без ганглиев.
* – достоверное (P < 0,05) отличие от контроля
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
При блокаде Н2-рецепторов гистамин вызывал большее увеличение ответов в препаратах трахеи с ганглиями по сравнению с действием аденозина, что может быть связано дилатационным эффектом аденозина в системе нижних дыхательных путей. Однако в препаратах бронхов констрикторный эффект гистамина, напротив, был меньше такового эффекта аденозина. Такой результат может свидетельствовать о различном качественном и количественном составе аденозиновых рецепторов в трахее и бронхах, на что указывалось в работах K. Varani и G. Vass [8], [9].
При блокаде Н1-рецепторов происходило уменьшение сокращения мышцы трахеи и бронхов как при действии гистамина, так и при действии аденозина. Однако при влиянии аденозина наблюдался гораздо больший дилатационный эффект, чем при действии гистамина. Большая дилатационная реакция в ответ на аппликацию аденозина, вероятно, связана с более широким его действием, по сравнению с гистамином, на структуры, опосредующие дилатационный эффект. Это может быть связано с действием аденозина на А2-рецепторы гладкой мышцы, стимулирующие аденилатциклазу и приводящие к инактивации киназы легких цепей миозина, что является причиной гладкомышечного расслабления [3]. Так же аденозин мог воздействовать на С-волокна неадренэргической нехолинэргичес-кой системы, запуская высвобождение оксида азота и вазоактивного интестинального пептида, опосредующих дилатацию. Кроме вышеупомянутых воздействий аденозина нельзя исключать влияния аденозина на тучные клетки с запуском последующей дегрануляции и высвобождением низких доз эндогенного гистамина. Выработанный тучными клетками гистамин оказывает влияние на Н2-и Н3-рецепторы, опосредующие дилатацию [6]. Влияния аденозина на нейроны интрамуральных ганглиев не наблюдалось, так как различия в величинах гладкомышечного сокращения препаратов с ганглиями и без них отсутствовали. По-видимому, нейрональные структуры метасимпатического ганглия не имеют рецепторов к аденозину.
ВЫВОДЫ
Основное констрикторное действие при влиянии гистамина и аденозина опосредовано активацией Н1-рецепторов, а основное дилатирующее – активацией Н2-рецепторов. Однако при влиянии аденозина в применяемой концентрации наблюдается больший дилатационный эффект, чем при действии гистамина, что связано с воздействием аденозина на различные структуры. С учетом того, что искусственно введенный в препараты трахеи и бронхов гистамин равен по концентрации (5–10 мкг/мл) эндогенно выработанному лаброцитами гистамину [5], следует заключить, что основной констрикторный эффект в системе нижних дыхательных путей связан с гистамином и не связан с действием аденозина или иных медиаторов тучных клеток, выделяемых при их частичной дегрануляции.
Таким образом, можно отметить, что действие аденозина и гистамина в нижних дыхательных путях оказывается тесно связанным. Влияние аденозина и гистамина на гладкую мышцу трахеи и бронхов опосредуется разными типами гистаминовых и аденозиновых рецепторов, которые могут быть локализованы на различных структурах респираторного тракта – в том числе непосредственно на гладкомышечных клетках, чувствительных окончаниях С-волокон, дыхательном эпителии, тучных клетках.
* Работа выполнена при поддержке Программы стратегического развития ПетрГУ в рамках реализации комплекса мероприятий по развитию научно-исследовательской деятельности на 2012–2016 гг.
ADENOSINE AND HISTAMINE LOW CONCENTRATION INFLUENCE ON MUSCLE CONTRACTION IN TRACHEA AND BRONCHI
Список литературы Влияние низких концентраций аденозина и гистамина на сокращения мышцы трахеи и бронхов
- Гусев Н. Б. Молекулярные механизмы мышечного сокращения//Соросовский Образовательный Журнал. 2000. T 6. № 8. С. 24-32.
- Anvari F., Sharma A. K., Fernandez L. G. Tissue-derived proinflammatory effect of adenosine A2B-receptor in lung ischemia-reperfusion injury//Thorac Cardiovasc Surg. 2010. Vol. 140. № 4. P. 871-877.
- Auchampach J. A., Gross G. J. Adenosine A1 receptors, KATP channels, and ischemic preconditioning in dogs//Am. J. Physiol. 2005. P. 264.
- Baraldi P. G., Cacciari B., Merighi S. et al. A(3) adenosine receptor ligands: history and perspectives//Med. Res. Rev. 2000. P. 20.
- Begueret H., Berger P., Vernejoux J. M., Dubuisson L., Marthan R., Tunon-De-Lara J. M. Inflammation of bronchial smooth muscle in allergic asthma//Thorax. 2007. Vol. 62. № 1. P. 8-15.
- Linden A., Eltzschig Y. K. Role of pulmonary adenosine during hypoxia: extracellular generation, signaling and metabolism by surface adenosine deaminase/CD26//Expert Opinion on Biological Therapy. 2007. Vol. 7. № 9. P. 1437-1447.
- Undem B., Kollarik M. The Role of Vagal Afferent Nerves in Chronic Obstructive Pulmonary Disease//The Proceedings of the American Thoracic Society. 2005. Vol. 2. P. 355-360.
- Varani K., Caramori G., Vincenzi F., Adcock I. Alteration of adenosine receptors in patients with chronic obstructive pulmonary disease//Am. J. Respir. Crit. Care Med. 2006. Vol. 173. P. 398-406.
- Vass G., Horvath I. Adenosine and adenosine receptors in the pathomechanism and treatment of respiratory diseases//Curr. Med. Chem. 2008. Vol. 15. № 9. P. 917-922.
- Winchilli V. M., Elswick R. K. The multivariante assessment of distributions//J. Royal. Stat. 2007. Vol. 7. № 1. P. 444-460.
- Zhong H., Belardinelli L., Maa T., Zeng D. Synergy between A2B adenosine receptors and hypoxia in activating human lung fibroblasts//Am. J. Respir. Cell. Mol. Biol. 2005. Vol. 32. P. 2-8.
- Zhou Y., Schneider D. J., Blackburn M. R. Adenosine signaling and the regulation of chronic lung disease//Pharmacol. Ther. 2009. Vol. 123. № 1. P. 105-116.
