Влияние пептида HLDF6 на пролиферативную активность спленоцитов в культуре клеток на фоне введения агонистов опиатных рецепторов

Автор: Сысоева Г.М., Даниленко Е.Д., Масычева В.И., Самуков В.В., Костанян И.А.
Журнал: Сибирский журнал клинической и экспериментальной медицины @cardiotomsk
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 4-1 т.24, 2009 года.
Бесплатный доступ
В культуре клеток исследовали влияние биологически активного пептида HLDF6 на пролиферативную активность спленоцитов мышей линии C57BL/6 на фоне введения селективных агонистов опиатных рецепторов мю-, дельта- и каппа-типа (DAGO, DADLE, динорфин (1-13)). Показано, что агонисты DAGO, DADLE и динорфин (1-13) обладали способностью усиливать пролиферативную активность лимфоцитов при их прямом взаимодействии с клетками. Интенсивность стимуляции определялась типом агониста и, соответственно, опиатного рецептора. Пептид HLDF6 повышал спонтанный и конкан- авалин-стимулированный пролиферативный ответ клеток при добавлении в среду на начальном этапе культивирования. Данные, касающиеся совместного использования HLDF6 и опиатных агонистов, свидетельствуют о взаимодействии пептида и эндогенной опиатной системы.
Пептид hldf6, агонисты опиатных рецепторов, пролиферативная активность, спленоциты, мыши
Короткий адрес: https://sciup.org/14919144
IDR: 14919144 | УДК: 612.014.3+616.8-091.81
Effect of HLDF6 peptide on splenocyte proliferation in vitro in the setting of opiate receptor agonists
Effect of bioactive HLDF6 peptide on proliferative activity of C57Bl/6 mice splenocytes in the setting of selective opiate receptors agonists was studied in cell culture. Specific agonists of m-, d- и k-receptors DAGO, DADLE and Dynorphin (1-13) were used. Opiate agonists were shown to be capable of enhancing proliferative response in mouse splenocytes during their direct interaction with cells. Data concerning combined administration of HLDF6 and opiate agonists point to the interaction between the peptide and the endogenous opiate system.
Текст научной статьи Влияние пептида HLDF6 на пролиферативную активность спленоцитов в культуре клеток на фоне введения агонистов опиатных рецепторов
Как известно, формирование синдрома наркотической зависимости сопряжено с изменением иммунного статуса организма [1-3]. Есть данные о способности ряда иммуномодуляторов снижать тяжесть абстинентного синдрома при лечении наркомании [1-4]. Это обусловлено тем, что между нейроэндокринной и иммунной системами существует тесная двусторонняя связь [1, 5-8], механизмы реализации которой привлекают пристальное внимание исследователей.
Важную роль в процессе коммуникации между нервной и иммунной системами играют опиоидные пептиды и специфические опиатные рецепторы. Многочисленными исследованиями установлено, что эндогенные опиоидные пептиды и синтетические агонисты опиатных рецепторов могут модулировать функции лимфоцитов, NK-клеток и макрофагов через присутствующие на их поверхности опиатные рецепторы трех основных типов: р, б и к [9-14].
В лаборатории белков гормональной регуляции Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова из культуральной среды клеток человеческого промиелоци-тарного лейкоза HL-60 был выделен белок HLDF с молекулярной массой 8,2 кДа, который вызывал дифференцировку клеток по гранулоцитарному пути. В структуре пептида был обнаружен шестичленный фрагмент HLDF6 (TGENHR), который воспроизводил способность полноразмерного фактора индуцировать дифференцировку и ингибировать пролиферацию клеток HL-60 [15].
Ранее было показано, что введение HLDF6 подавляло проявление основных симптомов абстинентного синдрома у мышей [16], нормализовало показатели ноцицептивных реакций у потомства морфинто- лерантных животных [17]. В процессе исследования влияния пептида на функцию опиатной системы было установлено, что HLDF6 отменял анальгетический эффект агонистов р- и к-рецепторов [18], усиливал адгезивные свойства и функциональную активность перитонеальных макрофагов, ослаблял эффекты селективных агонистов опиатных рецепторов [19]. Эти данные свидетельствует о взаимодействии пептида и эндогенной опиатной системы ЦНС и клеток иммунной системы, хотя вопрос об особенностях реализации этой взаимосвязи, в частности, на иммунокомпетентных клетках изучен пока недостаточно.
Целью данной работы являлось исследование влияния пептида HLDF6 на пролиферативную активность лимфоцитов селезенки и его способности модулировать иммунорегуляторные эффекты селективных лигандов р, б и к -опиатных рецепторов.
МАТЕРИАЛ И МЕТОДЫ
В качестве рецептор-специфических агонистов опиатных рецепторов были выбраны агонист μ-ре-цепторов DAGO (Tyr-D-Ala-Gly-(N-Me)Phe-Gly-ol), агонист δ-рецепторов DADLE (Tyr-D-Ala-Gly-Phe-D-Leu) и агонист к -рецепторов динорфин (1-13) (Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-Pro-Lys-Leu-Lys), полученные методом химического синтеза по специально разработанным методикам. Препараты DAGO и DADLE синтезировали по схемам (3 + 2). Синтез C-концевого сегмента DAGO проводили по специально разработанной трехстадийной схеме из 2-гидроксиэтиламида фенилаланина. Динорфин (1-13) синтезировали твердофазным методом на сти-рол-дивинилбензольной смоле Ванга с использованием Nsc-аминокислот. Пептид HLDF6 (Thr-Gly-Glu -Asn-His-Arg) был синтезирован методом активированных эфиров в растворе по авторской методике.
Спонтанную и индуцированную пролиферацию спленоцитов мышей линии C57Bl/6 оценивали после инкубации in vitro с пептидом HLDF6 и/или опиатными агонистами с помощью МТТ-теста по уровню восстановления клетками 3-(4,5-диметилти-азолил-2)-2,5-дифенил тетразолиум бромида (МТТ) [20]. Клеточную суспензию спленоцитов мышей готовили с использованием полной среды RPMI-1640, содержащей 10% эмбриональной сыворотки телят (10 мМ Hepes, 2 мМ L-глутамина, 50 мкг/мл гентамицина). Суспензию клеток распределяли по лункам 96-луночных круглодонных планшет по 100 мкл на лунку. Для индукции пролиферации использовали митоген – конканавалин А (ConA, ICN Biomedicals Inc) в концентрации 10 мкг/мл. Пептид добавляли в инкубационную среду непосредственно перед началом культивирования в концентрациях 0,01100 мкг/мл индивидуально либо в сочетании с опиатными агонистами (0,01-0,1 мкг/мл). Планшеты культивировали при температуре 37°С и 85-90% влажности в течение 72 ч. По окончании периода куль-55
тивирования во все лунки добавляли 20 мкл рабочего раствора МТТ с концентрацией 5 мг/мл, инкубировали в течение трех часов, после чего из лунок удаляли супернатант и вносили 200 мкл/лунку растворителя кристаллов красителя (формазана) – диметилсульфоксид (ДМСО). Планшеты оставляли при комнатной температуре на 30 минут, в течение которых происходило полное растворение кристаллов формазана. Оптическую плотность содержимого лунок замеряли с помощью спектрофотометра для микропланшетов при длине волны 540 нм.
Данные экспериментов подвергали статистической обработке с использованием пакета программ Statgrapfics 5.0 (Statistical Graphics Corp., USA). В качестве критерия статистической значимости обнаруженных межгрупповых различий был выбран t-критерий Стьюдента. Для подтверждения статистической значимости различий считали достаточным значение достигнутого уровня значимости (вероятности ошибочно принять отсутствие различий за их наличие) P<0,05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В ходе исследования было обнаружено, что внесение рецептор-специфических агонистов опиатных рецепторов в культуральную среду, как правило, приводило к повышению пролиферативной активности спленоцитов мышей. Однако интенсивность эффекта зависела как от типа агониста, так и от его дозы.
Повышение спонтанной пролиферативной активности клеток в результате воздействия на начальном этапе культивирования агониста д -рецепторов
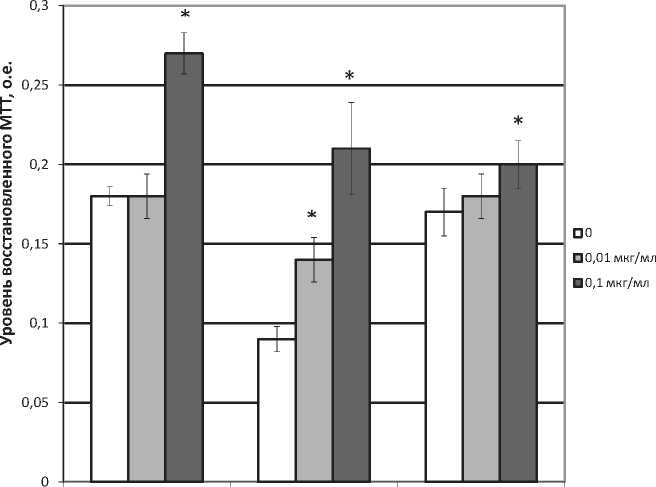
DAGO Динорфин (1-13) DADLE
Рис. 1. Влияние агонистов опиатных рецепторов μ-, δ- и κ-типа DAGO, DADLE и динорфина (1-13) на спонтанную пролиферативную активность спленоцитов мышей.
* – различия статистически значимы по отношению к контролю (физиологический раствор), р≤0,05
DAGO наблюдалось в дозе 0,1 мкг/мл: показатели МТТ-теста возрастали по отношению к контрольному уровню на 50% (рис. 1). Инкубация спленоцитов с агонистом к -рецепторов динорфином (1-13) также стимулировала клеточную пролиферацию, причем особенностью воздействия данного агониста являлось то, что эффект носил дозозависимый характер и обнаруживался уже в минимальной использованной дозе. Повышение клеточной пролиферации составляло 55 и 133% при внесении препарата в концентрациях 0,01 и 0,1 мкг/мл, соответственно.
Наименьшей способностью стимулировать пролиферативную активность лимфоцитов обладал агонист δ-рецепторов DADLE, который в дозе 0,1 мкг/мл повышал показатели МТТ-теста лишь на 19%. Описанные различия в активности опиатных агонистов разных типов рецепторов сохранялись и на клетках, обработанных Т-клеточным митогеном конканава-лином А.
Анализ имеющихся литературных данных подтвердил тот факт, что селективные агонисты опиатных рецепторов разных типов способны модулировать пролиферативный ответ лимфоцитов, причем есть сведения как о стимулирующем, так и об ингибирующем эффектах этих препаратов. Так, установлено, что селективные агонисты д -, б- и к -рецепторов (DAGO, DPDPE, U-50488) являются потенциальными стимуляторами митоген-индуцированной пролиферации Т-лимфоцитов мышей в системе in vitro, которая отменялась селективными антагонистами соответствующих рецепторов [21, 22]. В работах [23, 24] приводятся данные о стимулирующем влиянии Динорфина на митогениндуцированную пролиферацию спленоцитов крыс, пролиферативный ответ спленоцитов мышей и продукцию ими IL-2 и IL-1 β .
С другой стороны, показано, что агонист δ-рецепторов (лей-энкефа-лин) в зависимости от используемой дозы может негативно регулировать митогениндуцированную пролиферацию, стимулируя клетки-супрессоры или клетки-хелперы иммунного ответа [25, 26]. Есть данные и о зависимости направленности эффекта селективных агонистов μ- и δ- рецепторов DAGO и DADLE от состава клеточной фракции [27].
Пептид HLDF6, подобно агонистам опиатных рецепторов, усиливал пролиферативный процесс в культуре клеток селезенки мышей. Показатели пролиферативной активности спленоцитов после инкубации с пептидом в концентрациях от 0,01 до 10 мкг/мл возрастали в 2,4-5,1 раза (показатель спонтан- ной активности) или 2,2-3 раза в условиях воздействия митогена (табл.). Добавление HLDF6 в дозе 100 мкг/мл не приводило к дальнейшему усилению эффекта, более того, отмечена тенденция к некоторому снижению показателей МТТ в этих лунках по сравнению с показателями, зарегистрированными после инкубации клеток с пептидом в дозе 10 мкг/мл.
Культивирование спленоцитов в среде, содержащей DAGO и HLDF6, приводило как к усилению, так и к ингибированию способности клеток восстанавливать МТТ по сравнению с показателями, зарегистрированными в лунках с добавлением одного DAGO. При этом направленность эффекта зависела от концентрации агониста и интенсивности вызванной им реакции (рис. 2). Так, добавление пептида в среду, содержащую DAGO в концентрации 0,01 мкг/мл, которая не вызывала статистически значимого изменения спонтанной пролиферации спленоцитов, приводило к значительному повышению показателей МТТ-теста, превышавшему контрольный уровень в
1,5-1,8 раза. В том случае, когда результатом внесения DAGO (0,1 мкг/мл) являлось значительное повышение уровня пролиферативной активности клеток, пептид ослаблял эффект опиатного агониста. Те же закономерности были обнаружены при оценке последствий совместного применения агониста μ-ре-цепторов и HLDF6 в культуре клеток, индуцированных внесением в среду конканавалина А, где пептид в концентрациях 0,01 и 0,1 мкг/мл ослаблял стимулирующий эффект DAGO (0,1 мкг/мл) на пролиферацию митоген-индуцированных спленоцитов. Наиболее выраженный эффект был отмечен при использовании препаратов DAGO и HLDF6 в концентрациях 0,1 и 1 мкг/мл, соответственно, то есть при десятикратном избытке пептида.
Усиление под действием пептида HLDF6 спон- танной пролиферации спленоцитов в условиях слабо выраженного эффекта опиатного агониста наблюдалось и в экспериментах с использованием аго- ниста к-рецепторов динорфина (рис. 3). Отличи-
Таблица
тельной особенностью этих экспериментов являлось отсутствие
Пролиферативная активность спленоцитов мышей С57ВL/6 при культивировании в среде с добавлением пептида HLDF6
|
Препарат |
Концентрация, мкг/мл |
Уровень восстановленного МТТ, о.е. |
|
|
спонтанный |
митогениндуцированный |
||
|
Физраствор |
– |
0,155±0,189 |
0,335±0,043 |
|
HLDF6 |
0,01 |
0,352±0,014* |
0,734±0,030* |
|
0,1 |
0,564±0,015* |
0,884±0,031* |
|
|
1,0 |
0,732±0,028* |
1,030±0,031* |
|
|
10,0 |
0,767±0,015* |
1,017±0,011* |
|
|
100,0 |
0,630±0,027* |
0,947±0,039* |
|
* – различие статистически значимо по отношению к контролю (физиологический раствор), р<0,05.
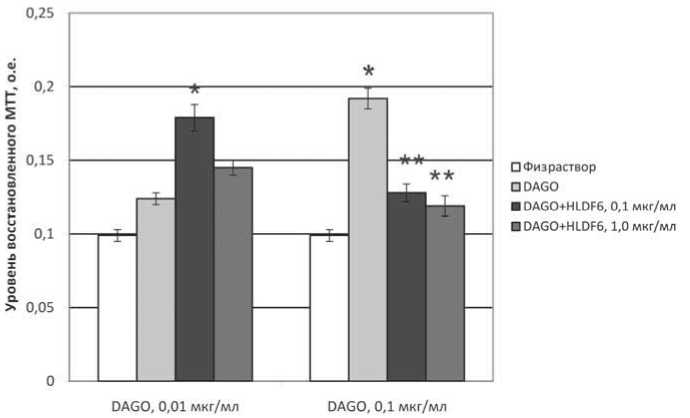
Рис. 2. Показатели спонтанной пролиферативной активности спленоцитов мышей при культивировании с агонистом опиатных μ-рецепторов DAGO и пептидом HLDF6.
Различия статистически значимы (р<0,05): * – по отношению к контролю (физиологический раствор); ** – по отношению к показателям в лунках, содержащих DAGO в соответствующей концентрации
статистически значимого ингибирующего эффекта пептида в условиях выраженной активации спонтанной пролиферативной активности клеток, вызванной динорфином. Кроме того, не было зарегистрировано сколько-либо значимого влияния HLDF6 на показатели митогениндуци-рованной пролиферации клеток на фоне добавления динорфина.
Внесение HLDF6 в среду культивирования, содержащую DADLE, в соотношении 1:1 и 10:1 не оказывало существенного влияния на показатели спонтанной и индуцированной пролиферативной активности, за исключением незначительного усиления эффекта агониста в концентрации 0,01 мкг/мл (на 21% по отношению к показателям в лунках с добавлением одного DADLE) (рис. 4).
Таким образом, установлено, что пептид HLDF6 обладал способностью модулировать эффекты агонистов разных типов опиатных рецепторов, изменяя как спонтанную, так и митоген-индуцированную активность лимфоцитов селезенки. Направленность эффекта пептида зависела от типа агониста и интенсивности вызванной им реакции.
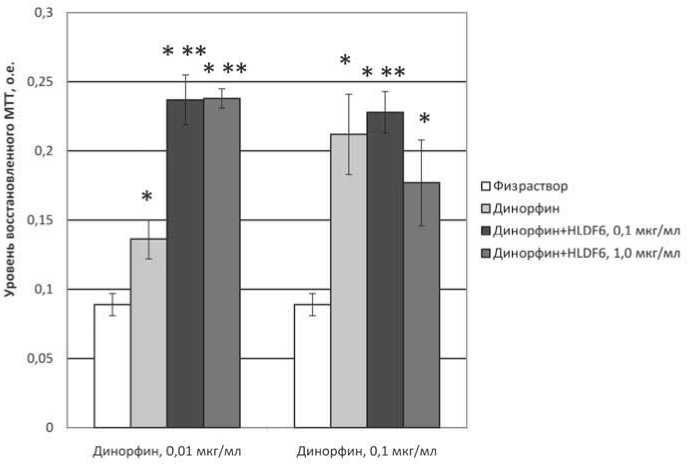
Рис. 3. Показатели спонтанной пролиферативной активности спленоцитов мышей при культивировании с агонистом опиатных κ-рецепторов динорфином (1-13) и пептидом HLDF6. Различия статистически значимы (р<0,05): * – по отношению к контролю (физиологический раствор); ** – по отношению к показателям в лунках, содержащих динорфин в соответствующей концентрации
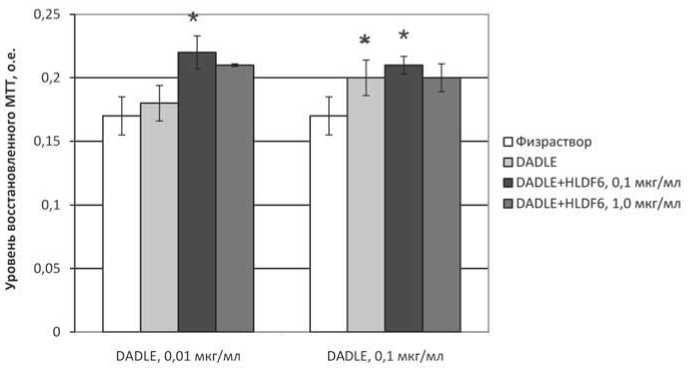
Рис. 4. Показатели спонтанной пролиферативной активности спленоцитов мышей при культивировании с агонистом опиатных δ-рецепторов DADLE и пептидом HLDF6. Различия статистически значимы (р<0,05): * – по отношению к контролю (физиологический раствор); ** – по отношению к лункам, содержащим DADLE в соответствующей концентрации
мом ингибирующего эффекта HLDF6 могут быть конформационные изменения структуры ионных каналов, в частности кальциевых, которые подвержены регуляции опиатными пептидами [15]. В связи с этим не исключено, что эффект HLDF6 на функциональное состояние спленоцитов может быть обусловлен деблокированием Ca2+-каналов, вызванным опиатами, и активацией Ca2+-кальмодулин-опосредованных путей передачи клеточного сигнала.
Таким образом, показано, что агонисты р-, б- и к -опиатных рецепторов (DAGO, DADLE, динорфин (1-13)) обладают способностью модулировать пролиферативную активность спленоцитов при их прямом взаимодействии с клетками. Направленность и интенсивность эффекта определялись типом агониста и, соответственно, опиатного рецептора. Пептид HLDF6 повышал пролиферативную активность спленоци-тов при добавлении его в инкубационную среду на начальном этапе культивирования. Данные, касающиеся совместного использования HLDF6 и опиатных агонистов, свидетельствуют о взаимодействии пептида и эндогенной опиатной системы. Направление эффекта пептида зависело от типа опиатного агониста и используемой дозы.
Механизмы модуляторного эффекта HLDF6 на спленоциты пока неясны. Известно, что связывание рецептор-специфических агонистов с опиатными рецепторами разных типов на поверхности клетки приводит к снижению активности аденилатциклазы и уровня цАМФ в клетке [28]. Добавление HLDF6 к активно пролиферирующим клеткам HL-60 вызывало аналогичные изменения (уменьшение синтеза цАМФ и фосфорилирования белков клетки) [29]. Однако подобная схема, если она верна и в отношении лимфоцитов, может объяснить только потенцирующий эффект HLDF6.
С другой стороны, как было установлено ранее, HLDF6 вызывает модификацию физико-химических свойств клеточной мембраны, повышая ее текучесть [15]. Следовательно, возможным механиз-
Список литературы Влияние пептида HLDF6 на пролиферативную активность спленоцитов в культуре клеток на фоне введения агонистов опиатных рецепторов
- Захарова Л.А., Василенко А.М. Опиоидэргическая система в сочетанной регуляции боли и иммунитета//Известия РАН. Сер. Биол. -2001. -№3. -С. 339-352.
- Зозуля А.А., Пшеничкин С.Ф. Опиоидные рецепторы и их лиганды//Итоги науки и техники/ВИНИТИ. Сер. Иммунология. -М. -1990. -Т. 25. -С. 48-120.
- Herz A. Role of immune processes in peripheral opioid analgesia//Adv. Exp. Med. Biol. -1995. -Vol. 373. -P. 193-199.
- McCarthy L., Wetzel M., Sliker J.K. et. al Opioids, opioid receptors, and the immune response//Drug Alcohol. Depend. -2001. -Vol. 62. -№2. -P. 111-123.
- Коростелев С.А., Исакова К.Л., Щеголев А.И., Утешев Б.С. Влияние эндогенных опиоидных пептидов и их синтетических аналогов на T-клеточный иммунитет//Эксперим. и клин. фармакология. -1994. -Т. 57. -№1. -С. 57-60.
- Чейдо М.А., Идова Г.В. Влияние опиоидных пептидов на процессы иммуномодуляции//Рос. физиол. журн. им. И.М. Сеченова. -1998. -Т. 84. -№4. -С. 385-390.
- Roy S., Loh H.H. Effects of opioids on the immune system//Neurochem.Res. -1996. -Vol. 21. -P. 1306-1375.
- Stefano G.B., Scharrer B., Smith E.M. et. al Opioid and opiate immunoregulatory processes//Crit. Rev. Immunol. -1996. -Vol. 16. -P. 109-144.
- Гейн С.В. Эффект динорфина А (1-13) на пролиферативный ответ лимфоцитов и изменение Th1/Th2 цитокинового профиля//Докл. РАН. -2009. -Т. 424. -№4. -С. 563-566.
- Гейн С.В., Симоненко Т.А., Черешнев В.А. Влияние μ-эндорфина и селективного агониста µ-опиатных рецепторов DAGO на пролиферативную активность лимфоцитов//Докл. РАН. -2003. -Т. 391. -№1. -С. 128-130.
- Alicea C., Belkowski S., Eisenstein T.K. et al. Inhibition of primary murine macrophage cytokine production in vitro following treatment with κ-opioid agonist U50,488H//J. Neuroimmunol. -1996. -Vol. 64. -P. 83-90.
- Band L., Pert A., Williams W. et al. Central µ-opioid receptors mediate suppression of natural killer cell activity in vitro//Prog. Neuroendocrin. Immunol. -1992. -Vol. 5. -P. 95-101.
- Sharp B.M. Multiple opioid receptors on immune cells modulate intracellular signaling//Brain Behav. Immun. -2006. -Vol. 20. №1. -P. 9-14.
- Taub D.D., Eisenstein T.K., Geller E.B. et al. Immuno-modulatory activity of mu-and kappa-selective opioid agonists//Proc. Nat. Acad. Sci. USA. 1991. -Vol. 88. -P. 360-364.
- Костанян И.А., Астапова М.В., Новолотская Е.В., Лепихова Т.Н., Драницына С.М., Телегин Г.В., Родионов И.Л., Байдакова Л.К., Золотарев Ю.А., Молотковская И.М., Липкин В.М. Биологически активный фрагмент фактора дифференцировки клеток линии HL-60. Идентификация и свойства//Биоорган. хим. -2000. -Т. 26. -№7. -С. 505-511.
- Литвинова С.В., Калюжный А.Л., Аристов В.В. и др. Подавление пептидом HLDF6 симптомов абстинентного синдрома на экспериментальной модели опийной наркомании//Бюл. эксперим. биологии и медицины -2004. -Т. 137. -№2. -С. 511-515.
- Литвинова С.В., Костанян И.А., Аристов В.В. и др. Применение пептида для коррекции патологии эндогенной опиоидной системы у потомства морфинтолерантных животных//Бюлл. эксперим. биологии и медицины -2003. -Т. 135. -№2. -С. 155-158.
- Федосова Л.К., Даниленко Е.Д., Масычева В.И., Костанян И.А., Самуков В.В. Оценка анальгетических свойств фрагмента пептидного фактора дифференцировки клеточной линии HL-60//Новые информационные технологии в медицине, биологии, фармакологии и экологии: 10-я юбилейная междунар. конф. и дискус. науч. клуб: -Запорожье, 2002. -С. 365-366.
- Даниленко Е.Д., Фадина В.А., Масычева В.И. и др. Модуляция пептидом HLDF6 функциональной активности макрофагов на фоне введения агонистов опиатных рецепторов//Мед. иммунология. -2005. -Т. 7. -№1. -С. 77-84.
- Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays//J. Immunol. Methods. -1983. -Vol. 65. -№1-2. -P. 55-63.
- Kowalski J. Immunomodulatory action of class mu-, and kappa-opioid receptor agonists in mice//Neuropeptides. -1998. -Vol. 32. -№4. -P. 301-306.
- Гейн С.В., Бабаева Т.А. Бетаэндорфин и селективные агонисты опиатных рецепторов в регуляции пролиферативного ответа лимфоцитов. Роль блокады опиатных рецепторов//Вестник Пермского университета. -2007. -№5. -С. 190-193.
- Ni X., Lin B.C., Song C.Y. et al. Dynorphin A enhances mitogen-induced proliferative response and interleukin-2 production of rat splenocytes//Neuropeptides. -1999. -Vol. 33. -№2. -P. 137-143.
- Bidlack J.M. Detection and function of opioid receptors on cells from the immune system//Clin. Diagnostic Lab.Immunol. -2000. -Vol. 7. -№5. -P. 719-723.
- Sizemore R.C., Dienglewicz R.L., Pecunia E. et al. Modulation of concanavalin A-induced, antigen-nonspecific regulatory cell activity by leu-enkephalin and related peptides//Clin. Immunol. Immunopathol. -1991. -Vol. 60. -№2. -P. 310-318.
- Lin J., Lu G., Jiang G. In vitro influence of enkephalins on the proliferative response of mouse and rat splenic lymphocytes to phytohemagglutinin//Proc. Chin. Acad. Med. Sci. Pecin Union Med. Coll. -1990. -Vol. 5. -№3. -P. 145-148.
- Гейн С.В., Баева Т.А., Гейн О.Н., Черешнев В.А. Роль моноцитов в реализации эффектов β-эндорфина и селективных агонистов µ-и δ-опиатных рецепторов на пролиферативную активность лимфоцитов//Физиология человека. -2006. -Т. 32. -№3. -С. 111-116.
- Keith D.E., Anton B., Murray S.R. et al. Mu-opioid receptor internalization: opiate drugs have differential effects on a conserved endocytic mechanism in vitro and in the mammalian brain//Molecular Pharmacology. -1998. -Vol. 53. -P. 377-378.
- Жохов С.С., Костанян И.А., Гибанова Н.В. и др. Сходство и различие действия пептидов TGENHR и TQVEHR на промиелоциты линии HL-60 и клетки Пуркинье червя мозжечка крыc//Докл. РАН -2004. -Т. 394. -№5. -С. 696-669.
