Влияние пролонгированного гипокинетического стресса на уровень тревожности и соотношение между липопероксидацией и карбонилированием белков в структурах головного мозга

Автор: Стрельников И.В., Цейликман О.Б., Цейликман В.Э., Манухина Е.Б., Дауни Ф.
Журнал: Человек. Спорт. Медицина @hsm-susu
Рубрика: Проблемы здравоохранения
Статья в выпуске: 3 т.13, 2013 года.
Бесплатный доступ
Увеличение продолжительности гипокинетического стресса до трех дней отменило анксиолитическое действие однодневного гипокинетического стресса и усугубило нарушения регуляции в системе гипоталамус - гипофиз - кора надпочечников. Это сопровождалось усилением перекисного окисления липидов в коре головного мозга.
Стресс, глюкокортикоиды, тревожность, перекисное окисление липидов
Короткий адрес: https://sciup.org/147153156
IDR: 147153156 | УДК: 576.35
The effect of prolonged hypokenetic stress on the anxiety level and the ratio of lipid peroxidation and carbonylation of proteins in brain structures
Extending the period of hypokinetic stress up to three days has brought to the calming action of a day-long hypokinetic stress and aggravated regulation disturbances in the hypophysis - pituitary - adrenocortical axis system. It’s followed by the increase of lipid peroxidation in the cerebral cortex.
Текст научной статьи Влияние пролонгированного гипокинетического стресса на уровень тревожности и соотношение между липопероксидацией и карбонилированием белков в структурах головного мозга
Введение. Стрессорные нарушения поведения в значительной степени опосредованы глюкокортикостероидами (ГКС), вследствие их способности вызывать угнетение функциональной активности нейронов префронтальной коры, гиппокампа, гипоталамуса, амигдалы и других структур ЦНС, что приводит к расстройству эмоциональной сферы и когнитивных функций [5]. При стрессе поддержание в течение длительного времени повышенного уровня ГКС обусловлено нарушениями регуляции гипоталамо-гипофизарно-адреналовой системы (ГГАС) по механизму длительной петли отрицательной обратной связи [4].
В условиях гипокинетического стресса наблюдается развитие поведенческих расстройств, изменение уровня свободно-радикального окисления в различных отделах ЦНС и увеличение уровня ГКС. Однако не понятно – сопровождается ли повышение содержания глюкокортикоидов при гипокинезии нарушениями регуляции ГГАС по механизму «длинной петли отрицательной обратной связи»? Также неизвестно как нарушения регуляции ГГАС соотносятся с изменениями свободнорадикального окисления в различных структурах головного мозга.
Методика. Исследование выполнено на лабораторных крысах массой 230–250 г обоего пола. Использовались беспородные животные. Гипокинетический стресс моделировали путём помещения животных на 1 сутки (ГК1) или на 3 суток (ГК3) в клетки-пеналы. Для изучения регуляции в ГГАС использовали дексаметазоновый тест, который проводился после завершения гипокинезии по схеме, учитывающей особенности циркадной ритмики ГГАС у лабораторных крыс [4]. В качестве оценочного показателя функционирования механизмов обратной связи использовали изменения уровня кортикостерона в плазме крови до (исходный уровень) и через 6 часов после введения дексаметазона (KRKA, Словения; внутрибрюшинно в дозе 5 мкг/кг). Поведенческие реакции животных изучались с использованием теста «приподнятый крестообразный лабиринт» [7]. Выделение отделов головного мозга крыс производилось в соответствии с рекомендациями, изложенными в работе [8].
Уровень кортикостерона определялся флюорометрическим методом Ю.Г. Балашова [1]. Содержание продуктов перекисного окисления липидов (ПОЛ) оценивали спектрофотометрически в липидном экстракте исследуемых тканей по методике И.А. Волчегорского и соавт. [6]. Окислительную модификацию белков оценивали по уровню образования динитрофенилгидразонов по методу Е.Е. Дубининой и соавт. [3].
Для обработки результатов исследований использовали пакет прикладных программ Statistica 6.0 for Windows. Статистически значимые различия между несколькими группами определялись с помощью критерия Краскелла – Уолиса (Kruskal-Wallis). Для определения статистически значимых различий между двумя сравниваемыми группами использовали критерии Манна – Уитни (U), Вальда – Вольфовица (WW). Различия считали значимыми при р ≤ 0,05. Статистические взаимосвязи изучали при помощи непараметрического корреляционного анализа, выполняя расчёт коэффициентов корреляции рангов по Спирмену (Rs) и Кен-деллу (rk).
Результаты и обсуждение. Установлено, что у интактных животных введение дексаметазона привело к снижению циркулирующего кортикостерона на 51 % по сравнению с исходным уровнем при ГК1 – только на 16 %, а при ГК3 введение дексаметазона не сопровождалось снижением уровня кортикостерона (см. рисунок). При этом содержание кортикостерона в группе «ГК3 + дексаметазон» статистически значимо повышено по сравнению с группой «ГК1+дексаметазон» и группой «дексаметазон».
Анксиолитическое действие односуточной гипокинезии проявлялось в увеличении времени пребывания в светлых рукавах «крестообразного лабиринта» с 98,72 ± 3,49 до 187 ± 5,27 с (Р = 0,036U). Наблюдаемые изменения поведенческой активности имели транзиторный характер и отсутствовали после завершения ГК3. Однодневная и трёхдневная гипокинезия характеризовались одинаковым по выраженности увеличением содержания кортикостерона крови (см. рисунок). Прирост содержания кортикостерона на базальном уровне может быть связан с нарушением регуляции по механизму «длиной петли отрицательной обратной связи», которое проявляется в снижении чувствительности гипоталамических центров типа PVN (паравентри-кулярное ядро) к глюкокортикоид-зависимому торможению продукции кортиколиберина. Через 120 ч после завершения ГК1 в коре головного мозга наблюдалось угнетение ПОЛ. Это проявлялось в снижении содержания изопропанол-растворимых диеновых конъюгатов и в уменьшении содержания Шиффовых оснований (см. таблицу). После завершения ГК3, напротив, наблюдалось усиление ПОЛ. Это проявлялось в увеличении содержания изопропанол-растворимых, а также гептан-раст-воримых кетодиенов и сопряжённых триенов по сравнению с контролем. Важно отметить, что содержание изопропанол-растворимых кетодиенов и сопряжённых триенов при ГК3 имело более высокое значение чем при ГК1. Пролонгирование гипокинетического стресса до трех суток привело к снижению содержания карбонилированных белков в гиппокампе (с 2,762 ± 0,193 мкмоль/г белка до 1,38 ± 0,35 мкмоль/г белка Р = 0,021U), а также в среднем мозге (с 8,254 ± 0,293 мкмоль/г белка до 4,248 ± 0,235 мкмоль/г белка Р = 0,014U). Кроме того, в среднем мозге на фоне повышенного содержания изопропанол-растворимых диеновых конъюгатов (с 0,364 ± 0,024 до 0,464 ± 0,025 у.е.о. Р = 0,014U) наблюдалось уменьшение содержания гептанофильных диеновых конъюгатов (с 0,817 ± ± 0,064 до 0,65 ± 0,035 у.е.о. Р = 0,014U), а также кетодиенов и сопряжённых триенов (с 0,624 ± 0,037 до 0,264 ± 0,025 у.е.о. Р = 0,014U). В связи с этим уместно обратить внимание на наличие положительной корреляции между содержанием изопро-панол-растворимых диеновых конъюгатов и временем пребывания в тёмных рукавах «крестообразного лабиринта» (Rs= 0,9; P = 0,037).
Полученные результаты свидетельствуют о том, что увеличение продолжительности гипокинезии до трёх суток проявляется в более выраженном нарушении регуляции ГГАС по механизму
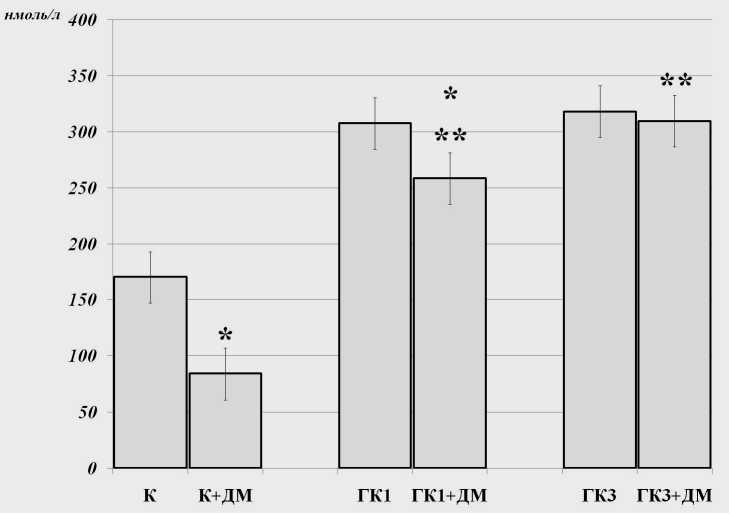
Содержание кортикостерона в плазме крови при гипокинезии после введения дексаметазона (дексаметазоновый тест): К - контроль (интактные животные); К + ДМ -группа «контроль» через 6 ч после введения дексаметазона; ГК (1 или 3) - гипокинезия; ГК + Дм - группа «гипокинезия» через 6 ч после введения дексаметазона; * - статистически значимое снижение содержания кортикостерона по сравнению с исходным уровнем; ** - межгрупповые различия
Проблемы здравоохранения
Содержание продуктов липопероксидации в коре головного мозга при введении глюкокортикоидного препарата после завершения гипокинезии
|
Показатель |
Диеновые конъюгаты (изопропанольная фаза) |
Кетодиены и сопряжённые триены (изопропанольная фаза) |
Шиффовы основания (изопропанольная фаза) |
|
1 Контроль (n = 5) |
0,319 ± 0,008 |
0,179 ± 0,004 |
0,059 ± 0,014 |
|
2 ГК1 (n = 6) |
0,285 ± 0,01 Р 1,2 = 0,036 |
0,17 ± 0,008 |
0,044 ± 0,005 Р 1,2 = 0,044 |
|
3 ГК3 (n = 6) |
0,36 ± 0,033 |
0,216 ± 0,010 Р 1,3 = 0,028 Р 2,3 = 0,016 |
0,05 ± 0,011 |
«длинной петли» отрицательной обратной связи. Это свидетельствует об увеличении порога гипоталамической чувствительности к ингибирующему влиянию глюкокортикоидов. Данный эффект гипокинетического стресса можно рассматривать как проявление гиперадаптоза, с позиций концепции В.М. Дильмана [2], или «аллостатической нагрузки», с точки зрения B. McEwen [9]. Увеличение продолжительности гипокинезии купировало свойственное односуточной гипокинезии анксиолитическое действие. Важно отметить, что это сопровождалось изменениями уровня свободнорадикального окисления в различных отделах головного мозга.
Выводы
-
1. Односуточная и трёхсуточная гипокинезии характеризуются увеличением концентрации кортикостерона в плазме крови на базальном уровне, а также после проведения дополнительного «дек-саметазонового теста». Введение дексаметазона после трёхсуточной гипокинезии вызвало более значительный прирост содержания кортикостерона по сравнению с односуточной.
-
2. В коре головного мозга при односуточной гипокинезии по сравнению с контролем снижено содержание изопропанол-растворимых молекулярных продуктов ПОЛ, а при трёхсуточной – повышено. После трёхсуточной гипокинезии по сравнению с контролем снижено содержание геп-тан-растворимых молекулярных продуктов при одновременном увеличении изопропанол-раство-римых продуктов ПОЛ в среднем мозге.
-
3. Пролонгирование гипокинетического стресса привело к снижению содержания карбонилиро-ванных белков в гиппокампе и в среднем мозге.
Работа выполнена в рамках реализации государственного задания на оказание услуг на 2012 и на плановый период 2013 и 2014 годов в части проведения научно-исследовательских работ № 4.4022.2011.
Список литературы Влияние пролонгированного гипокинетического стресса на уровень тревожности и соотношение между липопероксидацией и карбонилированием белков в структурах головного мозга
- Балашов, Ю.Г. Флюорометрический микрометод определения кортикостероидов: сравнение с другими методами/Ю.Г. Балашов//Физиол. журн. СССР. -1990. -№ 12. -С. 280-283.
- Дильман, В.М. Четыре модели медицины/B.М. Дильман. -М.: Медицина, 1987. -288 с.
- Окислительная модификация белков сыворотки крови человека, метод её определения/Е.Е. Дубинина, С. О. Бурмистров, Д.А. Ходов и др.//Вопр. мед. химии. -1995. -№ 41. -С. 24-26.
- Подвигина, Т.Т. Закономерности реагирования гипофизарно-адренокортикальной системы на повторные стрессорные повреждения/Т.Т. Подвигина//Успехи физиол. наук. -1998. -Т. 29, № 1. -С. 29.
- Филаретов, А.А. Принципы и механизмы регуляции гипофизарно-адренокортикальной системы/А.А. Филаретов. -Л.: Наука, 1987. -164 с.
- Экспериментальное моделирование и лабораторная оценка адаптивных реакций организма/И.А. Волчегорский [и др.]. -Челябинск: Изд-во ЧГПУ, 2000. -167 с.
- Belzung, C. Measuring normal and pathological anxiety-like behaviour in mice: a review/C. Belzung, G. Griebel//Behav. Brain. Res. -2001. -Nov 1;125(1-2). -Р. 141-149.
- Glowinski, J. Regional studies of catecholamines in the rat brain. The diposition of norepinephrine, dopamine and DOPA in varriosus regions of the brain/J. Glowinski, L.L. Iversen//J. Neurochem. -1966. -Vol. 13. -P. 655-669.
- McEwen, B.S. Stress and the individual: mechanisms leading to disease/B.S. McEwen, E. Stellar//Arch. Intern. Med. -1993. -Vol. 153. -P. 2093-2101.
- Rodgers, R.J. Influence of social isolation, gender, strain, and prior novelty on plus-maze behaviour in mice./R.J. Rodgers, J.C. Cole//Physiol. Behav. -1993. -Vol. 54(4). -Р. 729-736.
