Влияние сверхэкспрессии гена Са 2+-зависимой протеинкиназы VaCPK13 на устойчивость культур клеток винограда амурского Vitis amurensis Rupr. к абиотическим стрессам

Автор: Христенко В.С., Дубровина А.С., Алейнова О.А., Киселев К.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Сельскохозяйственные науки: Агрономия
Статья в выпуске: 2, 2015 года.
Бесплатный доступ
Полученные в результате исследования данные показали, что сверхэкспрессия гена VaCPK13 не влияла на устойчивость клеток V. amurensis к температурному, осмотическому и солевому стрессам. Полученные результаты свидетельствуют, что ген VaCPK13 не является сильным позитивным регулятором ответа растений на проанализированные абиотическиие стрессы.
Са 2+-зависимые протеинкиназы, абиотический стресс, клеточные культуры растений, cа 2+dependent protein kinase
Короткий адрес: https://sciup.org/14084129
IDR: 14084129 | УДК: 581.1
The influence of the gene overexpression Са 2+-dependent protein kinase VaCPK13 on the resistance of the Amur grape (Vitis amurensis Rupr.) cell cultures to the abiotic stress
The obtained as research result data showed that the VaCPK13gene overexpression did not influence on the stability of the V. amurensis cells to the temperature, osmotic and salt stress. The obtained results prove that the gene VaCPK13 is not a strong positive regulator of the plant response to the analyzed abiotic stress.
Текст научной статьи Влияние сверхэкспрессии гена Са 2+-зависимой протеинкиназы VaCPK13 на устойчивость культур клеток винограда амурского Vitis amurensis Rupr. к абиотическим стрессам
Понимание молекулярно-генетических механизмов ответа растений на неблагоприятные условия окружающей среды поможет понять, как растения справляются со стрессом. Приобретение растениями устойчивости к абиотическому стрессу нуждается в различных биохимических и физиологических изменениях, и большинство из них зависит от изменений в экспрессии генов. Исследования последних двух десятилетий показали, что различные стрессы влекут за собой сигнал-специфичные изменения уровня цитоплазматического Са2+, который функционирует как передатчик в модуляции разнообразных физиологических процессов, важных для адаптации к стрессам [4, 5].
Работа выполнена при поддержке Российского научного фонда (проект №14-14-00366).
Известно, что увеличение концентрации Ca2+ в цитозоли сигнализирует об изменениях в окружающей среде через связывание ионов Ca2+ с белками-сенсорами Ca2+, которые активируют дальнейшие события в сигнальной цепи. Одним из важнейших сенсорных белков в растительной клетке являются Ca2+-зависимые протеинкиназы (СDPK). CDPK играют важную роль в поддержании мембранного потенциала, регуляции углеводного и азотного обмена, устьичных движений и ответа клеток растений на абиотические и биотические стрессы [6]. Известно, что экспрессия генов и активность различных изоформ CDPK могут значительно возрастать в ответ на воздействие абиотических стрессов [7]. Кроме того, некоторые из идентифицированных белков-мишеней CDPK участвуют в защитном ответе клеток растений на абиотический стресс [8].
Особый интерес представляет изучение экспрессии генов CDPK в дикорастущих растениях с высоким уровнем устойчивости к стрессам, поэтому объектом нашего исследования был выбран виноград амурский Vitis amurensis Rupr., произрастающий на Дальнем Востоке России и обладающий высоким адаптивным потенциалом и устойчивостью к неблагоприятным условиям внешней среды. Ранее нами была изучена экспрессия генов CDPK под воздействием таких абиотических стрессовых факторов, как солевой стресс, водный дефицит, осмотический и температурные стрессы в дикорастущем V. amurensis. Согласно полученным данным, экспрессия гена VaCPK13 значительно увеличивалась при холодовом стрессе в растениях V. amurensis [9].
Цель работы . Изучение роли гена VaCPK13 в устойчивости дикорастущего винограда V. amurensis к абиотическим стрессам.
Материалы и методы исследования
Растительный материал и клеточные культуры. Каллусная культура V2 была получена в 2002 году из молодого стебля взрослого дикорастущего растения V. amurensis ( Vitaceae ), которое было собрано на Дальнем Востоке России (юг Приморского края) и определено в отделе ботаники Биолого-почвенного института ДВО РАН.
Для проведения экспериментов на культуре клеток винограда мы использовали W B/A агаризо-ванную питательную среду [10] с добавлением 0.5 мг/л 6-бензиламинопурина (БАП) и 2 мг/л α-нафтилуксусной (АНУ) кислоты, которую разливали в пробирки 150∙16 мм по 8 мл. Интервал субкультивирования составлял 30 дней в темноте при 23 ±1oС.
Получение трансгенных культур клеток винограда V. amurensis, сверхэкспресси-рующих ген VaCPK13, c помощью агробактериальной трансформации . Комплементарная (кДНК) гена VaCPK13 (номер доступа в ГенБанк KC488320) была амплифицирована после выделения РНК из листьев V. amurensis . Для амплификации полноразмерной кодирующей последовательности кДНК VaCPK13 использовали пару праймеров 5'ATG GGG AAC TGT TGC AGA T, 5'TTA CTC ATT CCC CAA GTT TAG. Полученный ОТ-ПЦР продукт VaCPK13 был выделен из геля при помощи набора Cleanup Mini kit (Евроген, Москва, Россия), клонирован в вектор pTZ57R/T согласно протоколу фирмы-производителя (Fermentas, Вильнюс, Литва). Затем, используя пару праймеров 5'GCT C GA GCT C AT GGG GAA CTG TTG CAG AT и 5'TCG A GG TAC C TT ACT CAT TCC CCA AGT TTA G, содержащих в своей последовательности сайты рестрикции для рестриктаз Sac I and Kpn I, мы ам-плифицировали полную последовательность VaCPK13. В качестве матрицы использовали предварительно полученную генетическую конструкцию pTZ57- VaCPK13 . Полученные ПЦР-продукты перенесли по рестрикционным сайтам Sac I and Kpn I в вектор pSAT1. В этой конструкции ген VaCPK13 находится под контролем двойного СаМV 35S промотора. После этого мы перенесли кассету, содержащую VaCPK13 , из вектора pSAT1 в вектор pZP-RCS2-nptII [11], используя сайт рестриктазы Pal AI (СибЭнзим, Новосибирск, Россия). Далее генетическая конструкция pZP-RCS2- VaCPK13-npt II была перенесена в агробактерии Agrobacterium tumefaciens GV3101::pMp90.
Суспензионную культуру клеток винограда V. amurensis V2 трансформировали полученными штаммами агробактерий по методике, описанной ранее [12]. После трансформации каллусы культивировали в течение 3 месяцев в присутствии 250 мг/л цефотаксима для подавления роста агробактерий. Отбор трансгенных клеток проводили на канамицине (Km) в концентрации 10–15 мг/л в течение 3–4 месяцев.
Воздействие абиотических стрессов на культуры клеток V. amurensis. Для анализа воздействия абиотических стрессовых факторов на устойчивость VaCPK13-трансгенных культур клеток мы исследовали влияние пониженной и повышенной температуры на рост трансгенных кле- точных культур (культивирование в течение 30 дней при температурах +16°C и +33°C) и влияние солевого стресса (культивирование в течение 30 дней при +23оС на питательных средах, содержащих NaCl в концентрации 60 и 120 мМ). NaCl добавляли в питательные среды после измерения pH. Также исследовали влияние осмотического стресса на рост трансгенных клеточных культур, сверх-экспрессирующих ген VaCPK13. Для этого эксперимент проводили при температуре +23оС, используя среду WB/A, содержащую маннитол в концентрации 200 и 300 мМ.
Выделение нуклеиновых кислот и получение комплементарной ДНК (кДНК) . Выделение РНК из культур клеток V.amurensis осуществляли на основе CTAB протокола по Bekesiova и др. [13]. кДНК была синтезирована, как описано ранее [12, 14].
Количественная оценка экспрессии гена VaCPK13 . Доказательство экспрессии экзогенного и эндогенного VaCPK13 , а также тотальной экспрессии VaCPK13 проводили с помощью ПЦР в реальном времени (ПЦР РВ). ПЦР РВ для гена был выполнен согласно методике, описанной Giulietti и др. [15]. Тотальную экспрессию гена VaCPK13 анализировали с помощью пары праймеров, подобранных к последовательности киназного домена гена VaCPK13 (табл.1). Эндогенную экспрессию VaCPK13 изучали, используя праймеры, один из которых был комплементарен концу последовательности киназного домена VaCPK13 , а другой к последовательности 3'-нетранскрибируемой области (3'UTR) VaCPK13 . Экспрессию дополнительной вставки гена VaCPK13 определяли, используя пару праймеров, подобранных к концу белок-кодирующей последовательности гена VaCPK13 и к последовательности CaMV 35S терминатора, находящегося в бинарной конструкции pZP-RCS2- npt II- VaCPK13 . Геноспецифичные пары праймеров представлены в таблице 1.
Праймеры для определения суммарной, эндогенной экспрессии и экспрессии трансгена VaCPK13
Таблица 1
кДНК амплифицировали с реактивом EvaGreen (Biotium, Хейвард, США). Для амплификации использовали амплификатор с функцией детекции результатов в реальном времени (ДНК Технология, Москва, Россия). Уровень экспрессии генов был определен по формуле 2-ΔΔCT. Полученное наивысшее значение экспрессии, накопленное отдельным образцом, было принято за единицу относительно количества мРНК. Гены VaActin1 (ГенБанк DQ517935) и VaGAPDH (ГенБанк GU585870) были использованы в качестве внутренних контролей для нормализации количества кДНК в каждой реакции ПЦР РВ. Данные ПЦР РВ были получены из 8 независимых реакций ПЦР РВ [16].
Статистический анализ полученных результатов. Результаты были обработаны при помощи программы Statistica, версия 10.0. Все данные представлены как среднее значение ± стандартная ошибка (СО). Полученные данные проверены по спаренному критерию Стьюдента. Уровень значимости в 0,05 был выбран как минимальное значение статистической разницы во всех экспериментах.
Результаты исследования. Трансгенные по гену VaCPK13 клеточные культуры КА-15-I, КА-15-II, КА-15-III и КА-15-IV были получены в результате независимых трансформаций суспензионной культуры V2 штаммом A. tumefaciens GV3101:pMp90, несущим векторную конструкцию pZP-RCS2- VaCPK13 - npt II. В конструкции ген VaCPK13 находится под контролем CaMV 35S промотора вируса мозаики цветной капусты [11]. Конструкция также несет ген устойчивости к Km ( npt II) под контролем
35S промотора. Сходным способом была получена контрольная клеточная линия КА-0, которая не содержала в конструкции ген VaCPK13 , а содержала только ген npt II .
Селекцию трансгенных клеточных агрегатов проводили в течение 3 месяцев на средах, содержащих 10–15 мг/л Km, и затем отобрали несколько устойчивых к Кm линий. В течение первого месяца трансформации мы отобрали быстрорастущие каллусы из отдельных маленьких агрегатов, которые обладали устойчивостью к Кm, и несколько Km-устойчивых независимых клональных линий KA-0, KA15-I, II, III, IV. Клеточная линия КА-0 идентична родительской клеточной культуре V2 по ростовым и морфологическим признакам. Это свидетельствует о том, что трансформация пустым вектором не вызвала значительных изменений в клеточных культурах. Клеточная линия КА-0 была использована далее как контрольная в ходе экспериментов. Четыре трансгенные клеточные линии KA15-I, II, III, и IV, активно экспрессирующие ген VaCPK13 , были получены в результате четырех независимых трансформаций. Данные ПЦР РВ показали, что трансген VaCPK13 активно экспрессируется во всех полученных клеточных линиях (рис.1, A), а экспрессия эндогенного гена VaCPK13 в трансгенных клеточных линиях КА15 не отличалась значительно от его экспрессии в контрольной клеточной линии КА-0 (рис.1, Б). Уровень тотальной экспрессии гена VaCPK13 увеличился в 1,6–4,8 раза по сравнению с контрольной клеточной линией КА-0 (рис.1, В).
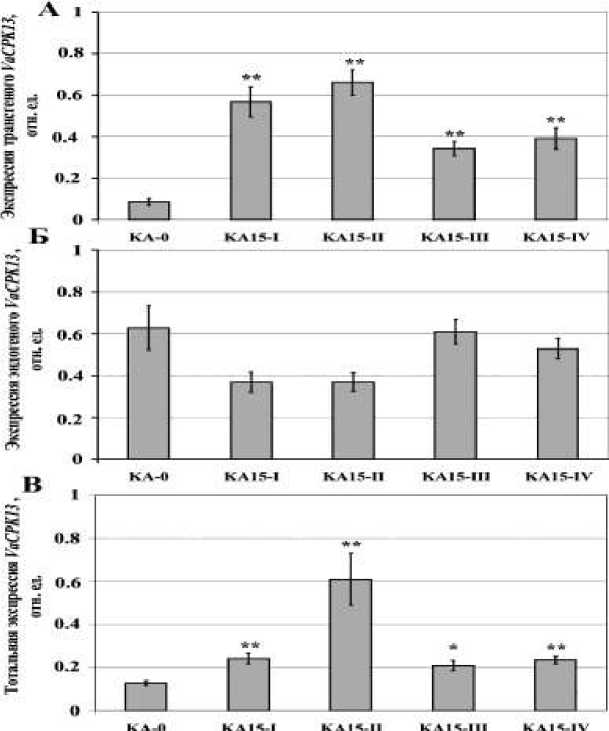
Рис.1. Экспрессия трансгена VaCPK13 (А), эндогенного VaCPK13 (Б), суммарная экспрессия эндогенного и экзогенного VaCPK13 (В) в клеточных линиях V. amurensis: КА-0 – контрольная клеточная культура, содержащая ген nptII (устойчивость к Кm); КА15-I, KA15-II, KA15-III и KA15-IV – трансгенные клеточные линии, сверхэкспрессирующие ген VaCPK13. Данные представлены как среднее значение ± СО; ** P < 0,01; * P < 0,05 по сравнению с уровнем флюоресценции для КА-0
Мы проанализировали эффект солевого, осмотического, теплового и холодового стрессов на рост четырёх VaCPK13- трансгенных клеточных линий V. amurensis . Сверхэкспрессия гена VaCPK13 незначительно увеличила устойчивость одной трансгенной клеточной линии V. amurensis (КА15-III) из четырёх к солевому стрессу (рис. 2, А).
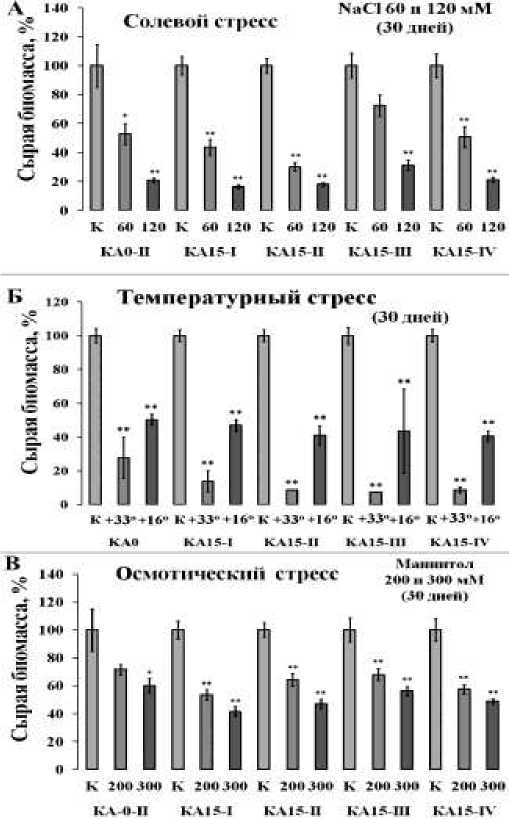
Рис.2. Накопление сырой биомассы клеточными линиями под влиянием солевого стресса, индуцированного 60 и 120 мМ NaCl при культивировании в течение 30 дней (А), теплового (+33о С) и холодового стрессов +16оС при культивировании в течение 30 дней (Б), осмотического стресса, индуцированного маннитолом 200 и 300 мМ в течение 30 дней (В): КА-0 – контрольная клеточная культура, содержащая ген nptII (устойчивость к Кm);
КА15-I, KA15-II, KA15-III и KA15-IV – трансгенные клеточные линии, сверхэкспрессирующие ген VaCPK13. Данные представлены как среднее значение ± СО; ** P < 0,01; * P < 0,05 по сравнению с накоплением биомассы клеточной линией КА-0
Устойчивость проанализированных трансгенных культур клеток к холодовому стрессу не отличалась от устойчивости контрольной клеточной линии КА-0 (рис. 2, Б). Сверхэкспрессия гена VaCPK13 ингибировала рост КА15-II и КА15-III трансгенных клеточных линий при действии теплового стресса (рис. 2, Б). Сверхэкспрессия гена VaCPK13 не вызывала значительных изменений роста клеточных линий при действии маннитола в концентрациях 200 и 300 мМ относительно роста контрольной клеточной линии КА-0 (рис. 2, В).
Заключение. В литературе присутствует информация о функциях и свойствах ближайших гомологов VaCPK13 у других растений (табл. 2). Известно, что ближайший гомолог VaCPK13 из Vitis vinifera, VvCPK10 активируется в период роста и развития растения [17]. Наибольшая экспрессия гена OsCPK3 в рисе наблюдалась в меристематических тканях [18]. Известно, что большое количество мРНК генов OsCPK3 и OsCPK16 образовывалось в каллусной культуре и в метёлках риса [19]. Также известно, что ген OsCPK16 является позитивным регулятором ответа риса на засуху и солевой стресс, а ген OsCPK3 – отрицательным регулятором при солевом воздействии [17]. Стоит отметить, что экспрессия гена PtCDPK14 из Populus trichocarpa увеличивалась при механическом воздействии в листьях и в корневых волосках [20].
Таблица 2
Сравнение выведенных аминокислотных последовательностей VaCPK13 (KC488320) c другими известными CDPK растений
|
Показатель |
I,% |
S,% |
|
VvCPK10 (VIT_08s0105g00390) |
99 |
99 |
|
AtCPK13 (NM115044) |
88 |
94 |
|
OsCPK3 (AP004366) |
80 |
87 |
|
OsCPK16 (AC108503) |
80 |
88 |
|
PtCDPK14 (POPTR_0016s12460) |
91 |
95 |
Примечание. I – аминокислотная идентичность; S – аминокислотная гомология.
В настоящем исследовании показано, что сверхэкспрессия гена VaCPK13 значительно не влияет на устойчивость клеток V. amurensis к солевому, тепловому, холодовому и осмотическому стрессам. Данные показывают, что ген VaCPK13 не является сильным позитивным регулятором ответа V. amurensis на проанализированные абиотические стрессы (холодовой, тепловой, осмотический и солевой стрессы) . Поскольку известно, что экспрессия гена VaCPK13 значительно возрастала при воздействии холодового стресса [14], можно предположить, что ген VaCPK13 не влияет на устойчивость растений напрямую как позитивный регулятор, но может участвовать в ответе винограда на холодовой стресс опосредованно либо в небольшой степени.
