Влияние внеклеточных везикул, происходящих из овариальных фолликулов различного размера, на созревание ооцитов и их развитие in vitro до стадии бластоцисты у коров (Bos taurus taurus)
")
Автор: Сингина Г.Н., Шедова Е.Н., Узбеков Р.Э., Узбекова С.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Репродуктивные технологии
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Внеклеточные везикулы, выделенные из фолликулярной жидкости (follicular fluid-derived extracellular vesicles, ffEVs) яичников, вовлечены in vivo в регуляцию оогенеза, что делает их перспективным объектом для оптимизации протоколов созревания ооцитов in vitro (in vitro maturation, IVM) и, как следствие, повышения эффективности получения эмбрионов in vitro (IVP). В настоящей работе мы впервые установили, что внеклеточные везикулы (extracellular vesicles, EVs) из фолликулярной жидкости (ФЖ) овариальных фолликулов малого (3-6 мм) и большого (более 6 мм) размеров равным образом положительно влияют на способность ооцитов развиваться до стадии бластоцисты после экстракорпорального оплодотворения. Цель представленной работы заключалась в изучении влияния внеклеточных везикул из фолликулярной жидкости различного фолликулярного происхождения в среде созревания коровьих ооцитов на их способность к эмбриональному развитию в условиях in vitro. Опыты проводили в ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста в 2024 году. ФЖ собирали аспирацией из коровьих фолликулов диаметром 3-6 мм (Small Follicles, SF) и более 6 мм (Large Follicles, LF) и последовательно центрифугировали при 300, 2000 и 12000 g в течение 15 мин, чтобы освободить ее соответственно от клеточного дебриса, апоптотических телец и крупных микровезикул. EVs из очищенной ФЖ выделяли посредством ультрацентрифугирования при 100 000 g. В результате из жидкости SF и LF были получены фракции, обогащенные EVs и содержащие соответственно 46,8 и 39,2 мкг тотального белка/мл ФЖ. Образцы проанализировали с использованием трансмиссионной электронной микроскопии, которая подтвердила, что в выделенных препаратах присутствуют EVs, соответствующие по размерам экзосомам. Ооциты коров созревали in vitro в контрольной среде (ТС-199, содержащей бычий сывороточный альбумин — 3 мг/мл, пируват натрия — 0,5 мМ и эпидермальный фактор роста — 100 нг/мл) или в среде IVM, дополненной ffEVs из ФЖ малых (ffEVs-SF) и больших (ffEVs-LF) фолликулов. Везикулярный белок добавляли в среду IVM в физиологической концентрации (на 1 мл среды количество EVs, выделенное из 1 мл ФЖ). Через 24 ч созревания ооциты подвергали экстракорпоральному оплодотворению (ЭКО) и культивированию для эмбрионального развития. На 3-и сут после оплодотворения изучали морфологию раздробившихся оплодотворенных ооцитов, на 7-е сут культивирования определяли число эмбрионов, развившихся до стадии бластоцисты (Бл). В полученных Бл определяли общее число ядер и долю ядер с признаками апоптоза (цитологический анализ). Доля созревших ооцитов была сходной в экспериментальных группах и варьировала от 81 до 84 %. Присутствие ffEVs в среде IVM не изменяло долю раздробившихся ооцитов после оплодотворения in vitro, которая составила 77,0±2,3, 81,3±2,0 и 83,3±2,5 % соответственно для контроля, ffEVs-SF и ffEVs-LF. Тем не менее было выявлено положительное влияние ffEVs как из малых, так и больших фолликулов на развитие созревших ооцитов до стадии Бл. При культивировании ооцитов в контрольной среде выход Бл составлял 24,0±3,0 %. Введение ffEVs-SF и ffEVs-LF в среду IVM повышало этот показатель соответственно до 37,4±0,7 (p < 0,01) и 41,7±1,0 % (p < 0,001). Было обнаружено положительное влияние везикул обоих видов на качество полученных Бл. В сравнении с контролем в группах ffEVs-SF (p < 0,01) и ffEVs-LF (p < 0,001) наблюдалось увеличение общего числа ядер в Бл, а также снижение доли ядер в Бл с признаками апоптоза (p < 0,001). Таким образом, использование ffEVs из ФЖ как малых, так и больших овариальных фолликулов в процедуре IVM повышает компетенцию коровьих ооцитов к развитию до стадии Бл после ЭКО, а также улучшает качество полученных бластоцист. Следовательно, ffEVs могут быть использованы для повышения эффективности получения эмбрионов крупного рогатого скота in vitro.
Внеклеточные везикулы фолликулярного происхождения, in vitro созревание, эмбриональное развитие, апоптоз, крупный рогатый скот
Короткий адрес: https://sciup.org/142247693
IDR: 142247693 | УДК: 636.2:591.39 | DOI: 10.15389/agrobiology.2026.2.350rus
Effect of extracellular vesicles from ovarian follicles of various sizes on bovine oocyte maturation and in vitro devel-opment to blastocyst stage
Follicular fluid-derived extracellular vesicles (ffEVs) are involved in vivo in the regulation of oogenesis, making them a promising tool for optimizing in vitro oocyte maturation (IVM) protocols and, consequently, for increasing the efficiency of in vitro embryo production (IVP). In the present study, we demonstrate for the first time that extracellular vesicles (EVs) derived from the follicular fluid (FF) of small (3-6 mm) and large (> 6 mm) ovarian follicles have similarly positive effects on the ability of oocytes to develop to the blastocyst stage after in vitro fertilization (IVF). The aim of this work was to investigate the effect of ffEVs of different follicular origin, when added to the maturation medium of bovine oocytes, on their competence to in vitro embryo development. The experiments were performed at the Laboratory of Embryo Technologies of the Ernst Federal Research Center for Animal Husbandry. FF was collected by aspiration from bovine follicles measuring 3-6 mm (small follicles, SF) and more than 6 mm (large follicles, LF), and sequentially centrifuged at 300 g, 2,000 g, and 12,000 g for 15 min to remove cellular debris, apoptotic bodies and large microvesicles, respectively. Small EVs from the clarified FF were isolated by ultracentrifugation at 100,000 g. As a result, ffEV-enriched fractions were obtained from SF and LF, containing 46.8 μg and 39.2 μg of total protein per ml of FF, respectively. The samples were analyzed using transmission electron microscopy, which confirmed that obtained preparations contain ffEVs, corresponding by size to exosomes. Bovine oocytes were matured in vitro either in control medium (TC-199 containing 3 mg/mL of bovine serum albumin, 0.5 mM sodium pyruvate and 100 ng/mL of epidermal growth factor) or in IVM medium supplemented with ffEVs from small (ffEVs-SF) or large (ffEVs-LF) follicles. EV-enriched preparations were added to IVM at a physiological concentration (a preparation of ffEVs isolated from 1 ml of FF to 1mL of culture medium). After 24 h of IVM, the oocytes were subjected to IVF and subsequent in vitro embryo development. On day 3 after the IVF, the morphology of cleaved fertilized oocytes was assessed; and on day 7 of culture, the number of embryos that had developed to the blastocyst stage was determined. In the obtained blastocysts, the total number of nuclei and the proportion of nuclei showing signs of apoptosis were assessed by cytological analysis. The rates of oocyte maturation were similar among the experimental groups, ranging from 81 % to 84 %. The presence of ffEVs in the IVM medium did not alter the cleavage rate after IVF, which were 77.0±2.3 %, 81.3±2.0 %, and 83.3±2.5 % in control, ffEVs-SF and ffEVs-LF groups, respectively. Nevertheless, a positive effect of ffEVs derived from FF of both small and large follicles was observed on the proportions of mature oocytes developing to blastocyst stage. In the control IVM medium, the blastocyst rate was 24.0±3.0%. Supplementation of IVM medium with ffEVs-SF and ffEVs-LF increased blastocyst rates to 37.4±0.7% (p < 0.01) and 41.7±1.0 % (p < 0.001), respectively. A positive effect of both types of ffEVs on blastocyst quality was observed. Compared with the control group, the ffEVs‑SF (p < 0.01) and ffEVs-LF (p < 0.001) groups showed an increased number of nuclei per blastocyst and a decreased proportion of nuclei exhibiting signs of apoptosis (p < 0.001). Thus, the use of ffEVs from small and large ovarian follicles in the IVM procedure increased the competence of bovine oocytes to develop to blastocyst stage after IVF and improved blastocyst quality. Therefore, ffEVs preparations may be used to improve the efficiency of in vitro embryo production in cattle.
Текст научной статьи Влияние внеклеточных везикул, происходящих из овариальных фолликулов различного размера, на созревание ооцитов и их развитие in vitro до стадии бластоцисты у коров (Bos taurus taurus)
Производство эмбрионов в условиях in vitro (IVP) — биотехнологический подход в современном скотоводстве, позволяющий воспроизводить большее количество потомства от высокоценных животных (1, 2). Статистические данные последних лет (3-5) показывают рост коммерческой привлекательности этого подхода и повышение мирового производства IVP-
* Работа выполнена при финансовой поддержке Российского научного фонда (проект ¹ 24-16-00261).
эмбрионов с 992 тыс. в 2017 году до 1876 тыс. в 2023 году. Одновременно возросший спрос на эмбрионы высокого качества определяет актуальность исследований, направленных на дальнейшее совершенствование отдельных этапов IVP (3). В частности, внимание уделяется поиску факторов, улучшающих качество созревающих in vitro ооцитов и повышающих их способность развиваться вне организма (6).
У млекопитающих естественной средой созревания ооцитов служит жидкость антральных фолликулов. Она содержит различные молекулы, которые опосредуют аутокринные и паракринные связи между фолликулярными клетками и созревающим ооцитом, играя решающую роль в оогенезе (7, 8). В этих коммуникациях участвуют внеклеточные везикулы (extracellular vesicles, EVs), включающие экзосомы и микропузырьки, которые секретируются клетками фолликула и способны влиять на клеточный ответ и биологическую активность посредством переноса разнообразных регуляторных молекул — РНК, белков и липидов (9, 10). Показано, что EVs фолликулярной жидкости (follicular fluid-derived extracellular vesicles, ffEVs) играют важную роль в регуляции функций клеток кумулюса, передаче сигналов от соматических клеток к ооциту (11) и, в конечном счете, в контроле развития ооцитов (12). Это позволяет рассматривать ffEVs в качестве перспективного компонента для оптимизации сред IVM для повышения качества ооцитов млекопитающих, включая крупный рогатый скот (КРС) (13, 14).
На сегодняшний день показано, что ffEVs в условиях in vitro могут существенно влиять на изменения транскриптома, протеома, экспрессии генов в ооцитах и окружающих их клетках кумулюса (КК) (14-17), а также на потенциал ооцитов к дальнейшему эмбриональному развитию (14, 1820). У КРС в среде in vitro созревания (in vitro maturation, IVM) ооцит-ку-мулюсных комплексов ffEVs усиливали экспансию КК (21) и их пролиферацию (22). Кроме того, обнаружен позитивный эффект ffEVs на коровьи ооциты, которые созревали в условиях воздействия стрессовых факторов, таких как тепловой шок и окислительный стресс, либо после созревания in vitro замораживались. То есть при воздействии везикул повышалась стрессо-и криоустойчивость ооцитов (15, 23, 24).
Хотя описанные выше результаты убедительно демонстрируют положительное влияние ffEVs на ооциты в системе IVM, остается неясным, сохраняется ли одинаковый эффект при использовании внеклеточных везикул, выделенных из фолликулов различного размера. Этот вопрос требует дальнейшего изучения, поскольку размер фолликула связан с его физиологическим статусом и качеством фолликулярной среды (25), что потенциально может определять состав и биологическую активность EVs (26, 27).
В настоящей работе мы впервые установили, что внеклеточные везикулы из фолликулярной жидкости (ФЖ) овариальных фолликулов малого (3-6 мм) и большого (более 6 мм) размеров равным образом положительно влияют на способность ооцитов развиваться до стадии бластоцисты после экстракорпорального оплодотворения.
Цель представленной работы заключалась в изучении влияния внеклеточных везикул из фолликулярной жидкости различного фолликулярного происхождения в среде созревания коровьих ооцитов на их способность к эмбриональному развитию в условиях in vitro.
Методика. Опыты проводили в ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста в 2024 году. Во всех экспериментах, за исключением отдельно отмеченных случаев, использовали реагенты фирмы «Sigma-Aldrich» (США).
Яичники коров (Bos taurus taurus) неизвестной породной принадлеж- ности, отобранные post mortem на мясокомбинате, транспортировали в лабораторию на льду. После доставки их освобождали от прилегающих соединительных и жировых тканей, а затем многократно промывали в охлажденном до +4 °С физиологическом растворе (ФЗ), содержащем пенициллин (100 МЕ/мл) и стрептомицин (50 мкг/мл), чтобы удалить компоненты крови и микробную контаминацию. Фолликулярную жидкость (ФЖ) собирали аспирацией из фолликулов диаметром 3-6 мм (small follicles, SF) и более 6 мм (large follicles, LF) и центрифугировали (центрифуга 5424 R, «Eppendorf AG», Германия) при 300 g в течение 15 мин.
На следующем этапе для освобождения от апоптотических телец и крупных микровезикул размером 200-1000 нм ФЖ центрифугировали (центрифуга 5424 R, «Eppendorf AG», Германия) по 15 мин сначала при 2000 g, а затем при 12000 g. После каждого этапа очищения полученный супернатант собирали и переносили в новые пробирки. EVs выделяли из очищенной ФЖ ультрацентрифугированием (центрифуга CS 150 NX, «Hitachi», Япония) в течение 90 мин при 100 000 g с последующим разведением супернатанта фосфатно-солевым буфером (ФСБ) и повторным ультрацентрифугированием в том же режиме.
Осадок, полученный после ультрацентрифугирования, ресуспенди-ровали в 100 мкл ФСБ и хранили до использования при - 80 ° С, предварительно отобрав от полученной суспензии две аликвоты объемом по 5 мкл. Одну аликвоту использовали для определения количества EVs по концентрации белка: анализ проводили на приборе Qubit 4 Fluorometer с использованием набора Qubit Protein Assay Kit («Thermo Fisher Scientific, Inc.», США) и белкового стандарта Qubit с концентрацией от 0,125 до 5 мг/мл. Из второй аликвоты для установления присутствия EVs готовили цитологический препарат и проводили его ультраструктурный анализ с помощью трансмиссионной электронной микроскопии (ТЭМ, микроскоп JEOL 1011, «Jeol, Ltd.», Япония), как описано ранее (28).
Для функционального эксперимента выделенные post mortem яичники коров доставляли с мясокомбината в лабораторию в теплом ФЗ (не ниже 28 ° С) и препарировали, как описано выше, используя физиологический раствор, предварительно подогретый до 37 ° С. Ооциты в составе ооцит-кумулюсных комплексов (ОКК) выделяли из яичников механическим способом — рассекали стенки видимых фолликулов лезвием, после чего промывали и проводили селекцию извлеченных ОКК, как было описано ранее (29). Незрелые ОКК с соответствующими морфологическими характеристиками созревали in vitro в среде ТС-199, содержащей бычий сывороточный альбумин (БСА, 3 мг/мл), пируват натрия (0,5 мМ), эпидермальный фактора роста (100 нг/мл), а также антибиотик гентамицин (50 мкг/мл) в течение 24 ч. В опытных группах в среде IVM дополнительно присутствовали EVs выделенные из жидкости фолликулов диаметром 3-6 мм (ffEVs-SF) и диаметром более 6 мм (ffEVs-LF). В обоих случаях везикулы использовали в физиологической концентрации: на 1 мл добавляли количество EVs, выделенное из 1 мл ФЖ. ОКК инкубировали группами по 25-30 ОКК в 4-лу-ночных планшетах (ООО «Биомедикал», Россия) в каплях среды объемом 500 мкл и в условиях инкубатора MCO-18AIC («Sanyo», Япония) при 38,5 ° С и 5 % СО 2 в атмосфере.
Во втором эксперименте по завершении периодов созревания (24 ч IVM) ооциты подвергали процедуре экстракорпорального оплодотворения с целью получения эмбрионов in vitro. Экстракорпоральное оплодотворение проводили в среде BO-IVF («IVF Bioscience», Великобритания). Бычью сперму размораживали и подвергали процедуре swim-up c использованием 352
среды Sperm-TALP (30). Полученную фракцию активных сперматозоидов вносили в среду ЭКО с предварительно перенесенными туда ОКК до конечной концентрации 1,5½106 сперматозоидов/мл. Продолжительность совместной инкубации гамет составляла 15-16 ч. Затем ооциты освобождали от клеток кумулюса и налипших сперматозоидов и оценивали морфологически. Выявляли наличие у ооцитов полярных телец (ПТ) и определяли процент созревания (долю ооцитов с ПТ от общего числа ооцитов). Изолированные ооциты переносили в среду BO-IVC («IVF Bioscience», Великобритания) и инкубировали в ней для эмбрионального развития. Культивирование происходило при температуре 38,5 ° С и в присутствие 5 % СО 2 , 5 % О 2 и 90 % N 2 . Через 3 сут культивирования in vitro (IVC) подсчитывали число раздробившихся ооцитов, на 7-е сут инкубации определяли число ооцитов, развившихся до стадии бластоцисты.
Для оценки качества полученных бластоцист готовили и анализировали их цитологические препараты, используя протокол, описанный ниже. Эмбрионы промывали в ФСБ, содержащем 0,2 % БСА (ФСБ-БСА), и фиксировали 4 % раствором параформальдегида в ФСБ в течение 1 ч при комнатной температуре. После промывания в ФСБ-БСА образцы подвергали процедуре пермеабилизации в 0,5 % растворе Triton Х-100 в течение 30 мин. Детекцию апоптотических ядер в эмбрионах проводили методом TUNEL с использованием набора In Situ Cell Death Detection Kit, fluorescein («Roche Diagnostics», Швейцария) согласно инструкции компании-производителя. Затем эмбрионы обрабатывали в течение 20 мин раствором DAPI (1 мкг/мл) для окрашивания ядер и переносили на предметное стекло с последующим заключением в среду Vectashield («Vector Laboratories», Великобритания).
Микрофотографирование и анализ препаратов выполняли под флуоресцентным микроскопом Axio Imager M2 («Carl Zeiss AG», Германия), оснащенным фильтром для TUNEL (возбуждение при X = 445-470 нм) и DAPI (возбуждение при X = 365 нм). Использовали цифровую камеру Axiocam 506 и программу Zen pro («Carl Zeiss», Германия). Определяли общее число ядер в эмбрионах и число TUNEL-позитивных ядер. Степень апоптоза оценивали по доле TUNEL-позитивных ядер от общего числа ядер в бластоцистах.
Статистическую обработку данных проводили при помощи лицензионного программного пакета SigmaStat 2.03.0 («Systat Software, Inc.», США). Число независимых опытов — 3. Результаты выражали как средние значения ( M) и стандартные ошибки средних (±SEM). Для оценки статистической значимости различий применяли однофакторный дисперсионный анализ (ANOVA) с поправкой Тьюки. Различия считали достоверными при p < 0,05.
Результаты. Метод выделения EVs из фолликулярной жидкости коров основывался на дифференциальном ступенчатом центрифугировании и ультрацентрифугировании при 100 000 g, что позволяет эффективно получать фракцию EVs, представленных преимущественно экзосомами диаметром 30-150 нм. Первоначально эта методика была предложена для изоляции EVs из жидкости яйцевода (31), а в настоящем исследовании применена в несколько модифицированном виде (28). В результате из жидкости средних (SF) и больших (LF) фолликулов были получены фракции, обогащенные EVs и содержащие соответственно 46,8 и 39,2 мкг тотального белка/мл ФЖ. Полученные препараты были проанализированы с использованием ТЭМ, которая подтвердила присутствие EVs типа экзосом в обоих образцах, что послужило основанием для их дальнейшего применения в среде созревания ооцитов.
На рисунке 1 представлены типичные EVs, полученные из фолликулярной жидкости яичников коров. Отмечалось наличие как отдельных везикул, так и их агрегатов.
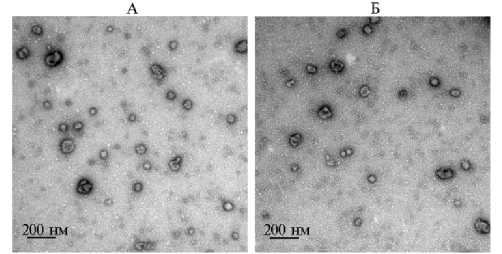
Рис. 1. Изображения внеклеточных везикул из фолликулярной жидкости малых (ffEVs-SF) (А) и больших (ffEVs-LF) (Б) антральных фолликулов коров ( Bos taurus taurus ) неизвестной породной принадлежности, полученные с помощью трансмиссионной электронной микроскопии (увеличение 100 000½, микроскоп JEOL 1011, «Jeol, Ltd.», Япония) (University of Tours, 2024 год).
Через 24 ч культивирования коровьих ооцитов in vitro доля MII ооцитов в контроле, оцениваемая по наличию по крайней мере одного полярного тельца на момент окончания процедуры оплодотворения, составила 81,6±0,84 % (рис. 2, А). Присутствие везикул как из малых (ffEVs-SF), так и больших фолликулов (ffEVs-LF) не изменяло значение показателя, что свидетельствует об отсутствии их влияния на этот процесс.
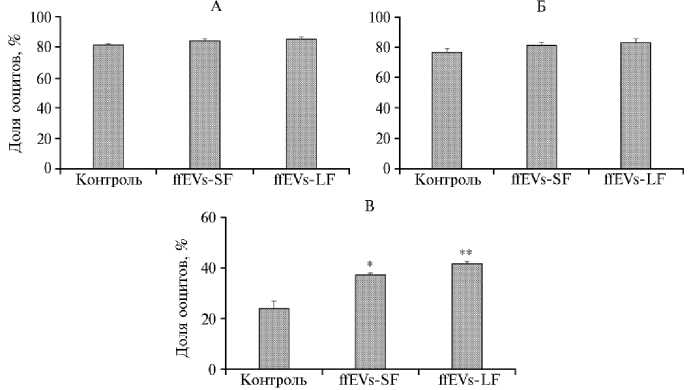
Рис. 2. Доля ядерного созревания ооцитов (А) , их дробления (Б) и развития до стадии бластоцисты (В) после in vitro созревания в контроле и под влиянием ffEVs из фолликулярной жидкости фолликулов диаметром от 3 до 6 мм (ffEVs-SF) и более 6 мм (ffEVs-LF) и последующего экстракорпорального оплодотворения ( n = 3, M ±SEM; лаборатория эмбриональных технологий ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024 год) .
* и ** Различия статистически значимы по сравнению с контролем соответственно при р < 0,01 и р < 0,001.
Для оценки компетенции зрелых ооцитов к эмбриональному развитию их подвергали экстракорпоральному оплодотворению. Доля дробления ооцитов в контроле составила 77,0±2,2 % (см. рис. 2, Б). Введение ffEVs в среду in vitro созревания существенно не меняло это значение: для ffEVs-SF и ffEVs-LF показатель составил соответственно 81,3±2,0 и 83,3±2,3 %. Тем не менее обнаружено влияние ffEVs как из LF, так и SF в период IVM коровьих ооцитов на их дальнейшее развитие до стадии бластоцисты (см.
рис. 2, В, рис. 3, а-в). В случае созревания ОКК в контрольной среде выход бластоцист составил 24,0±3,0 %. Введение ffEVs-SF и ffEVs-LF в среду IVM в физиологической концентрации повышало показатель до 37,4±0,7 (p < 0,01) и 41,7±1,0 % (p < 0,001).
Контроль ffEVs-SF ffEVs-LF
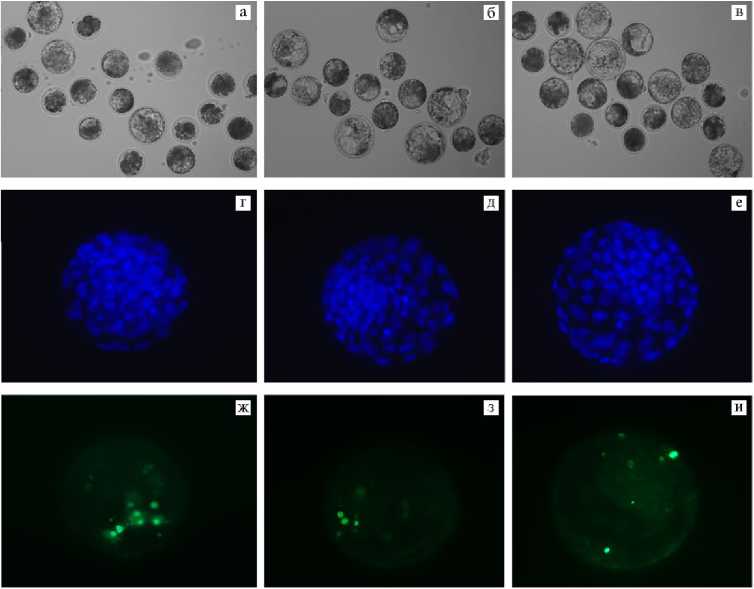
Рис. 3. Микрофотографии эмбрионов коров ( Bos taurus taurus ) неизвестной породной принадлежности, полученных вследствие экстракорпорального оплодотворения ооцитов, созревших в контроле и при воздействии внеклеточных везикул из жидкости овариальных фолликулов малого (ffEVs-SF) и большого (ffEVs-LF) размеров: а-в — эмбрионы, развившиеся до стадии бластоцисты (увеличение 100½, микроскоп Eclipse Ti-U, «Nikon», Япония); г-е — окрашивание ядер в бластоцисте с помощью DAPI (синий цвет; цитологический препарат), ж-и — окрашивание апоптотических ядер в бластоцисте методом TUNEL (зеленый цвет; цитологический препарат) (увеличение 400½, микроскоп Axio Imager M2, «Carl Zeiss», Германия).
Оценка качества бластоцист, полученных вследствие экстракорпорального оплодотворения коровьих ооцитов, созревших в контроле и при воздействии внеклеточных везикул из жидкости овариальных фолликулов малого (ffEVs-SF) и большого (ffEVs-LF) размеров ( n = 3; лаборатория эмбриональных технологий ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024 год)
|
Группа |
Общее число бластоцист, шт. |
Число ядер в бластоцисте, M ±SEM |
|
|
всего, шт. |
с признаками апоптоза, % |
||
|
Контроль |
16 |
79,5±1,4 |
6,6±0,3 |
|
ffEVs-SF |
28 |
84,5±0,8* |
4,4±0,2** |
|
ffEVs-LF |
31 |
87,7±1,1** |
3,8±0,1** |
* и ** Различия статистически значимы по сравнению с контролем соответственно при р < 0,01 и р < 0,001.
Охарактеризовано качество эмбрионов (табл.), полученных из ооцитов, созревших при воздействии ffEVs из фолликулов различного диаметра по общему числу ядер (см. рис. 3, г-е) и числу ядер с признаками апоптоза (см. рис. 3, ж-и). Цитологический анализ показал, что в контрольной группе в бластоцистах в целом число ядер соответствовало стадии развития. Добавка ffEVs-SF приводила к увеличению этого показателя (p < 0,01), равно как и добавка ffEVs-LF (p < 0,001). Также присутствие ffEVs в среде IVM ооцитов снижало долю ядер в бластоцистах с признаками апоптоза (p < 0,001). При этом значимых различий внутри опытных групп по обоим показателям обнаружено не было.
Несмотря на то, что ffEVs принимают участие в регуляции гамето-и эмбриогенеза у разных видов млекопитающих (11, 12), работ, посвященных их применению для оптимизации условий развития ооцитов и эмбрионов in vitro, немного. У животных, включая КРС особого внимания заслуживают присутствующие в фолликулярной жидкости малые внеклеточные везикулы — экзосомы, которые в основном секретируются клетками гранулезы, теки и кумулюса (32). В ряде исследований по использованию этих везикул в системе культивирования ооцитов было продемонстрировано их участие в регуляции экспрессии критически важных для качества яйцеклеток генов (17), а также выявлено положительное влияние ffEVs на способность яйцеклеток к последующему эмбриональному развитию (15).
В настоящем исследовании ffEVs добавляли в среду IVM, чтобы повысить полноценность яйцеклеток и, как следствие, эффективность получения IVP эмбрионов у КРС. При этом вносимые внеклеточные везикулы выделяли из фолликулярной жидкости овариальных фолликулов разного размера. Мы впервые обнаружили, что присутствие ffEVs не только из малых (диаметр 3-6 мм), как это было продемонстрировано ранее (13, 33, 34), но и из больших (диаметр более 6 мм) овариальных фолликулов положительно влияет на способность ооцитов развиваться до стадии бластоцисты после экстракорпорального оплодотворения (см. рис. 2, В), а также повышает качество полученных Бл (см. табл.). Эти результаты свидетельствуют о положительном влиянии обоих видов ffEVs в условиях in vitro на цитоплазматическое созревание ооцитов коров, несмотря на то, что от размера фолликула может зависеть состав и биологическая активность ffEVs (26, 27).
Таким образом, воздействие внеклеточных везикул из фолликулярной жидкости яичников (ffEVs) на коровьи ооциты во время их созревания in vitro способствует повышению компетенции к эмбриональному развитию in vitro после экстракорпорального оплодотворения. Это проявляется в повышении выхода бластоцист на 7-е сут культивирования оплодотворенных ооцитов, увеличении общего числа ядер в бластоцистах и снижении доли ядер с признаками апоптоза. Эффект наблюдается как при использовании ffEVs из овариальных фолликулов малого (диаметр 3-6 мм), так и большого (диаметр более 6 мм) размеров. Следовательно, внеклеточные везикулы, выделенные из жидкости таких фолликулов, могут быть использованы для оптимизации условий культивирования ооцитов крупного рогатого скота, созревающих in vitro.
ФГБНУ Федеральный исследовательский центр животноводства — БИЖ им. академика Л.К. Эрнста, 142132 Россия, Московская обл., г.о. Подольск, пос. Дубровицы, 60,
