Возможности повышения интенсивности отборов из перекрестноопыляющихся популяций

Автор: Дьяков А.Б., Бойко Ю.Г., Васильева Т.А.
Статья в выпуске: 2 (129), 2003 года.
Бесплатный доступ
Рассмотрение методов селекции и улучшающего семеноводства сортов подсолнечника и других перекрестников с позиций популяционной генетики и биологии популяций позволило сделать ряд важных выводов для практической селекции. При надежной идентификации продуктивных генотипов рекуррентная селекция может быть эффективной при использовании широкого диапазона интенсивностей отборов. Применявшиеся минимальные интенсивности отборов семеноводческой элиты практически не влияли на степень инбредности популяций и могли приводить к утрате только очень редких аллелей. При использовании же в селекции сортов подсолнечника однократного и, особенно, многократного индивидуального отборов приросты коэффициента инбридинга были выше ΔF=12,5 % на один цикл, была высока вероятность утраты даже аллелей, содержащихся в популяциях с высокой частотой. Такая высокая интенсивность отборов позволяет эффективно концентрировать желательные гены в генофонде популяции при условии высокой надежности как оценок семей потомков элит по результатам деляночных испытаний, так и идентификации генотипов по их фенотипам при отборах элитных растений. Кроме этого должны приниматься также другие меры минимизации инбредной депрессии и истощения генетической изменчивости в популяциях.
Короткий адрес: https://sciup.org/142150655
IDR: 142150655 | УДК: 633.854.78:631.52
Текст научной статьи Возможности повышения интенсивности отборов из перекрестноопыляющихся популяций
Независимо от используемого метода селекции ее основой является отбор наилучших особей и семей в качестве родителей следующего поколения. Поэтому селекционный прогресс в значительной, часто в решающей степени определяется результативностью отборов внутри популяций. Особенно важно оптимизировать на научной основе методику отборов на урожайность и другие количественные признаки при работе с пере-крестноопыляющимися популяциями. При этом необходимо учитывать основные факторы, определяющие величину селекционного сдвига при отборе.
Сдвиг при отборе (реакция на отбор) оценивается разницей между фенотипической средней величиной признака у потомства отобранной части популяции и средним значением тою же признака у исходной популяции. Зависимость величины селекционного сдвига 01 влияющих на нес факторов выражается формулой:
R- i-ap-h \ (I)
где К - селекционный сдвиг признака; i - селекционный дифференциал, т. с. выраженная в единицах стандартного отклонения разница между средними величинами признака исходной популяции и группы отобранных из нее растений или семей; оР - фенотипическое стандартное отклонение признака; Һ2- коэффициент наследуемости признака в узком смысле (реализуемый). Из формулы ( Г) следует, что результативность отборов на количественный признак тем выше, чем меньше процент отбираемых лучших растений или семей, чем выше изменчивость признака в исходной популяции и чем точнее определяется по фенотипам селекционная ценность генотипов. Эти требования определяют комплекс мер оптимизации селекционного процесса.
При селекции самоопыляющихся видов растений основным путем повышения вариабельное 1 и признаков за счел’ і енотипического полиморфизма является гибридизация. При рабо те же с перекрестноопыляющимися популяциями особенно важно после зевательными циклами отборов повышать концентрацию положи тельных генов, нс допуская при этом близкородственных скрещиваний, и обеспечивать высокую степень генетических рекомбинаций.
Повышать наследуемость признака, т.е. надежность идентификации желательных генотипов по их фенотипам особенно трудно при селекции на урожайность. По данным разных исследователей, коэффициент наследуемости этого признака особей популяций подсолнечника нс превышает h?-0J8 (Дьяков, 1991). При такой наследуемости средняя величина признака потомков отобранных особей незначительно отличается от показателей исходной популяции (Рокицкий, 1978, с. 310). Реально наследуемость признака при оіборах на урожайность cine меньше но той причине, что значительная доля наследственной изменчивоеги этого признака, обусловленная различиями генотипов но конкуре гр । оснособнос’ и и продолжительности вегетации, селекционно бесполезна (Дьяков, 1987). Озатко академик В.С. Нустовойт как в процессе селекции, гак и при улучшающем семеноводстве сирюв подсолнечника систематически повышал сборы масла за снег увеличения урожаев семян (ядер семянок), что свилетельствусч о принципиальной возможности эффективной идентификации желательных генотипов но их феночипам (Дьякон, 1987. 2003). Для исключения эффектов конкуренции, искажающих оценки генотипов, В.С. Пус-ювойі использовал в качестве фонового признака повышенную масличность семян (ядер семянок), характеризующую способность растений эффективно использовать ресурсы среды и за счет этого (формировать высокий урожай семян вследствие увеличения числа запасающих клеток без усиления жиронакопления в каждой клетке (Дьяков, 1988, 2003). По резулыаіам изучения нотомств В.С. Нустовойт выбраковывал позднеспелые семьи.
При испы таниях семей потомков можно в 2,7 раза уменьшить искажающие эф-фекчы межлеляночной конкуренции путем замены однорядных делянок квадратными, увеличивая при этом и число учетных растений с 22 до 32 (Дьяков, Хатнянский, Васильева, бойко, 1^96). Увеличение числа учтенных растений эффективно повышает наследуемое и. средней величины признака семей нолусибов, что показано в таблице I, данные коюрой вычислены но формуле (Falconer, 1960):
14),25(N-I)
һ ^ [ТоДЛД^Т) һ_ (2)
где һ hsl и Іг соответственно коэффициенты наследуемости семей нолусибов и отдельных расіений: N - число изученных растений семьи; 0,25 - коэффициент родства для нолусибов.
Полученные результаты (табл. 1) свидетельствуют о том, что при низкой насведу ем ости признака особей особенно важно увеличивать число изучаемых растений в каждой семье потом ков-нолусибов. Это позволяет в 4-7 раз повышать коэффициенты наследуемости средних величин признака семей. Даже если селекционно полезна лишь половина наследуемой доли дисперсии урожайности и коэффициент наследуемости признака особей реально составляет лишь ҺМ),!, вполне приемлемым увеличением числа изучаемых расіений каждой семьи можно повысить наследуемость средних величин признака ло h2hs! 0,7 и выше. Для этой цели лучше использовать предлагаемые квадратные дечянки, иены гания на. которых позволяют не только резко уменьшать искажения оценок урожайное г и конкуренцией, но и обеспечивать нужный уровень наследуемости признака при меньшем числе полевых повторностей.
Повышать вариабельность Ср признака в популяциях и коэффициент его наследуемости 11 сложно, а возможности воздействия на результативность селекции с помощью таких мер сравнительно ограничены. Интенсивность же отборов селекционер нланнрусі сам и их ужесточение не требует осуществления дополнительных мероприятий. При этом повышением интенсивности отборов, т. е. оставлением в качестве родителей следующею поколения самых лучших растений или семей, можно в большой степени увеличиваю селекционный дифференциал і и соответственно повышать величину селекционного сдвига признака R (Уильямс, 1968, с. 319-321). Повышать селекционный дифференциал особенно важно при низких коэффициентах наследуемости признака (Дубинин, Глембоцкии, 1967, с. 463).
Тайзица L Влияние числа изученных особей семьи нолусибов на соотношение между коэффициентами наследуемости и средней величины
1/ ри $ н а к а сем ьи (h hsj) и отд ельн ых рас тек ий (Һ )
|
Число учетных растений на делянке |
Число повторностей |
Всего растений |
1 Іаследуемосгь средних величин признака семей при наследуемости признака особей |
|||
|
ь2™о, 1 |
һЧ),2 |
ьЧи |
һЧ),4 |
|||
|
22 |
1 |
22 |
0.41 |
0,6! |
0,73 |
0,81 |
|
2^ |
3 |
44 |
0.57 |
0,75 |
0,83 |
0,89 |
|
2? |
3 |
66 |
0.66 |
0.81 |
0,88 |
0.92 |
|
32 |
] |
32 |
0.49 |
0,69 |
0,79 |
0.85 |
|
32 |
э |
64 |
0.65 |
0.81 |
0.88 |
0.92 |
|
32 |
3 |
96 |
0,73 |
0,86 |
0.91 |
0.94 |
Гюли из популяции отбирается большое число растений, семьи потомков отдельных уникальных генотипов нс могут существенно повысить среднюю величину признака потомств всех отобранных особей (Гуляев, Гужов, 1972, с. 341). Эксперименты Г.В. 1’у-ляева, АЛ. Гсрезкина и Л.И. Долгодворовой (1983) показали, что при отборах из пере-крестпоопыляющейся популяции ржи значимый селекционный сдвиг урожайности в потомстве достигался при отборах с последующим переопылением от 2 до 10 растений, а при отборе 25 или более особей урожайность переопыленных потомков существенно не отличалась от показателей исходной популяции.
13 результате свободного комбинирования гамет в каждом поколении перекрест-нооныляющейся популяции возникаю! редкие уникальные генотипы, вобравшие в себя большое количество генов, благоприятно влияющих на селекционные признаки. Сохранить в потомстве высокую концентрацию желательных генов можно только при отборе небольшого числа самых ценных генотипов и использовании близкородственных скрещиваний (Baker, Curnow, 1969). Наиболее эффективно сохраняется в популяции потомков высокая частота ценных генов при принудительном самоопылении уникальных родителей (Спрэг, 1957). Это позволяет закрепить такие аллели, т. е. перевести их в гомозиготное состояние. Однако при малом числе отбираемых особей происходит как дрейф генов со случайной утратой ценных аллелей других локусов, влияющих на селекционные признаки. так и переход в гомозиготное состояние вредных рецессивных аллелей, что приводит к фенотипическому проявлению нежелательных признаков и ин бред ной де прессті и роста и продуктивности растений. Интенсивное снижение гстерозиготности при самоопылении так быстро истощает генетическую изменчивость, что у селекционера не хватает времени для проведения эффективного отбора в пределах линии.
Под инбридингом понимается не только самоопыление, но и любые другие системы родственных скрещиваний, приводящие к повышению уровня гомозиготности (Ьрипс, Ноулз, 1972, с. 200). Ограничивать рассеивание ценных комбинаций генов в популяции и тем самым закреплять достигнутый уровень признаков, на которые ведется отбор, можно и более слабыми формами инбридирования. Издавна широко используется в селекции перскрсстноопыляющихся культур такая форма более слабого инбридинга как переопыление в пределах семей полусибов, т. с. потомков одного материнского растения и разных отцов (Macaulay, 192 8; Пустовойт, 1940; Эллиот, 1961. с. 313; Гуляев, Гужов,
1972, с. 234). К методам еще более слабого инбридинга относится скрещивание нескольких лучших по комплексу признаков растений со сходным чипом роста, отбираемых в каждом из ряда последовательных поколений (Harlan, 1950),
Чем больше скорость иибридированищ гем более аффективна концентрация ценных юнов в популяции при отборе, но тем раньше и в большей степени уменьшается ! снетнческнй полиморфизм и проявляется инбредная депрессия (Гуляев, Гужов. 1972, с. 234-245; Cornelius, I aylor, 1981; Бороевич, 1984, с. 116. 120). В связи с этим селекционеру приходится решать проблему, которую хорошо сформулировал Дж. Ф.Спрэг (1957): "Какая часи> популяции должна быть сохранена при каждом цикле отбора? Любой ответ па эго! вопрос является компромиссным. Чем меньше сохраненный образец, 'гем больший шаг вперед сделан при данном цикле отбора. Но при небольших размерах образца \шеличиваегея степень инбридинга, а поэтому быстро истощается генетическая изменчивость'. Увеличенное число отбираемых растений важно и для уменьшения случайной утраты аллелей вследствие дрейфа генов. Л. Меттлер и Г. Грегг (1972. с. 89) показали, ч то дрейф генов связан с ограничением возможности выбора партнера в малых субпопу-ляциях. что увеличивает родство особей, вступающих в скрещивания. С учетом эчих обстоятельств интенсивность отборов приходится поддерживать на уровне значительно ниже максимального (Уильямс, 1968. с. 33 Г) и важнейшей задачей при планировании программ рекуррентной селекции является определение оптимальной интенсивностп отборов. минимального числа родителей каждою очередного поколения (Rawlings. 1981). Бороевич. 1984, с. 120). Закон минимум отбираемых из популяции особей должен быть дос газонным для гою, чтобы не допустить инбредпой депрессии, а также сохранить вт ।ршюпуляционную генетическую изменчивость для дальнейших отборов, поскольку более важен общий успех, а не улучшения, достигнутые за один цикл отбора (Спрэг. 19S7; Уильямс, 1968, с. 337, Каминская, *985. с. 16).
Генетическое обоснование минимально допустимого числа отбираемых из популяций растений в качестве родителей следующего поколения важно не только при селек-пип, но !! при семеноводстве сортов нерекресгпоопыляюшихся культур, особенно если ведется улучшающее семеноводство. Н.Н. Дубинин и ЯЛ. Глембоцкий (1967, с. 247-250) подчеркивают, что эффективный контроль сложной, динамической генетической струю гуры сор гон-популяций подсолнечника, ржи, кукурузы и других перс крести и ко в часто оказывается делом значительно более чрудным. чем создание таких сортов. Однако до сих пор остается актуальным вывод Г.К Гуляева и В.Г. Шестакова (1970, с. 39): ‘ Многие нормы и правила в семеноводе те нс обоснованы теоретически1? При гаком обосновании, по мнению Г.В. Гуляева и ЮЛ. Гужова (1972. с. 342), - "очень важно экспериментально установить, какое число растений в отборах воспроизводит сорт у различных перекрест И О О Н Ы Л Я Ю Щ И X С Я КУ, I bl V р".
Представления селекционеров о минимальном числе отбираемых растений., достаточном для воспроизведения признаков сорта-популяции, очень различны Например, С.Бороевич ( 1984. с. 336) считает, ч то при семеноводстве растений-псрекрестников идентичность сорт можно сохранить только в том случае, если для производства элитных семян отбирать такое количество растений, которое позволит охватить почти все имеющиеся в популяции генотипы. Абсолютная нереальноеі ь такой задачи слелусз из важнейшею вывода популяционной генетики: "... какой бы большой ни была панмиктическая популяция, в ней не существует двух особей (за исключением идентичных близнецов), все гены которых были бы одинаковыми" (Ли, 1978, с. 360). Хотя разнообразие генотипов в свободнооныляющихся популяциях гак велико, GJ? Sprague (1939) установил, что средние пока газели десяти растений сорта кукурузы адекватно отражают его наследсг-6
венные особенности. Опыты Г.В. Гуляева и Л.И. Долгодворовой (1976) с сортами-популяциям и озимой и яровой ржи показали, что потомство переопыленных между собой 25-тн растений сорта полностью воспроизводит все его признаки. Также чисто 'эмпирическим путем в Швеции разработана методика первичного семсноводсчва сортов озимой ржи, при которой! тбирасіся 40-50 элитных растений, и по результатам испытаний. выбираются лучшие из их потомств на основе использования метода резервов (Гуляев. Гужов, 1972, с. 343-344). Обобщая экспериментальные данные, Г.В. Гуляев (1988) пришел к выводу, что не только у самоопыляющихся, но и у перекрестноопыляющихся культур сорт может воспроизводиться небольшим числом растений.
Принято сличать, что при сборе коллекций генетических ресурсов дикорастущих и культивируемых видов растений генофонд популяций почти полностью представляют семена, собранные с 50-100 растений (Hawkes, 1974). Такой размер выборки считается достаточным потому, что 50-100 особей с вероятностью 95 % сохраняю! все аллели, встречающиеся в популяции с частотой не менее 0,05 (Cristca, Gaspar, 1979), а нриросч гомозиготности при эффективной численности популяции 50 особей составляет 1 % на поколение (Франклин, 1983). Однако К. Yonezawa (1985) считает более эффективной стратегию поддержания и использования коллекций генетических ресурсов при отборах по 10 растений па популяцию. При таких выборках нс гарантируется с вероятностью 95 % сохранение аллелей, встречающихся в популяции с частотой 0,30 и меньше (Яблоков, 1987, с. 182).
13 программах рекуррентной селекции перекрестноопыляющихся растений, осуществляемых с целями как повышения урожайности селектируемых популяций, так и увеличения вероятности выделения из них особо ценных инородных линий, обычно рекомендуется в каждом цикле отбирать для формирования новой популяции по 10 генотипов (Sprague, Miller, Brimhall, 1952; Спрэг, 1957; Югенхеймер, 1979, с. 88; Каминская, 1985, с. 54). В качестве исходного материала для селекции родительских линий гибридов создаются также синтетические популяции из 6-8 инбредных линий (Хаджинов, Гусев. 1979; Югенхеймер, 1979, с. 315).
При таких числах основателей популяций не гарантируется с вероятностью 95 % сохранение в каждом цикле отбора аллелей, содержащихся в исходной популяции с частотой меньше 0,3-0,5, поэтому неизбежна эрозия генофонда. А поскольку основным преимуществом рекуррентной селекции является сохранение изменчивости популяций в каждом цикле отборов для обеспечения дальнейшего прогресса (Бриггс. Ноулз, 1972, с. 235), возникает вопрос, в какой мере при интенсивных отборах достигается эта цель и предотвращается инбредная депрессия.
При обсуждении проблемы сохранения наследственной изменчивости в популяциях и их реакции на отбор после ряда циклов рекуррентной селекции разные авторы а ши । изируют итоги многолетних опытов с популяциями кукурузы, дрозофилы и других объектов. Например, L.R. Reeder, A.R. Hallauer, K.R. Lamkey (1987) не обнаружили существенных изменений генотипической дисперсии и соотношения ее компонент после 6-ти циклов успешной рекуррентной селекции двух популяций кукурузы на урожайность. G.F. Sprague, Р.А. Miller, В. Brimhall (1952) установили, что при отборах на масличность зерна с самоопылением без скрещивания лучших линий к пятому поколению в линиях почти не осталось генетической изменчивости, а при чередовании отборов и скрещиваний изменчивость признака существенно увеличилась по сравнению с исходной популяцией. Особенно информативны результаты проведенного в штате Иллинойс длительного опыта, в котором изучалась реакция кукурузы на отборы по процентному содержанию масла и белка в зерне. Отборы были начаты в 1896 г., до 25-го поколения в каждом вари- анте в каждом цикле отбирали но 24, а затем - но 12 растений при сохранении интенсивности отборов во все годы 20% от числа оцененных особей каждой популяции, чему соответствует шипение і^1,4с7рһ. За 50 поколений отборов было достигнуто повышение масличное! и в 3,3 раза и се снижение в 4,7 раза, повышение содержания белка в 1,8 раза и ею снижение в 2/2 раза (Woodworth, Leng, Jugenhcimer, 1952). Анализ экспериментальных популяций в 1966 г. показал, что 65 поколений отборов на повышение содержания как масла, так и белка не только не привели к истощению наследственной изменчивости, но даже оказалось, ч то у высокомасличной популяции генотипическая дисперсия процента масла на 65 % больше, чем у высокобелковой, а генотипическая дисперсия процента белка у высокобелковой популяции на 60 % больше, чем у высокомасличной (Dudley, Lambert, 1969). За 76 поколений реакция па отбор достигла 20 генетических (аддитивных) стандартных отклонений в сторону увеличения как масличности, так и содержания белка, причем регрессия процента масла на число поколений оказалась практически прямолинейной, без признаков выхода се на плато (Dudley, 1977).
Анализируя результаты этого опыта, У. Уильямс (1968, с. 324) обращает внимание не только на непредсказуемо большие реакции на отбор в этих экспериментальных популяциях. Рще более важным он считает то, что при изменении в обратную сторону направлений отборов в субпопуляциях всех четырех популяций происходит сдвиг признаков в сторону исходных популяций. Он подчеркивает, что это противоречит представлениям классической гене!ики о том. что переход в результате длительных отборов в гомозиготное состояние все большего числа желательных генов не только должно дово-аи гь до нуля генетическую вариансу, но и отбор в обратном направлении не должен дос-лначь пределов показателей исходной популяции. Поэтому результаты описанного долгосрочно! о опыта с кукурузой и аналогичные опыты с популяциями других аутбред-ных видов заставляют признать существование иных источников значительной наследственной изменчивости (Уильямс, 1968, с. 324-325). На основе математического моделирования 10.М. Свирежев и В.П. Пассков (1982, с. 247) также заключили, что только признав существование более сложных механизмов наследственной изменчиво-сзи, можно объяснить способность популяции эффективно реагировать на изменения направлений отбора. Анализируя результаты опытов по отбору, Я.Р. Франклин (1983) пришел к выводу: “Маше понимание сил, лежащих в основе поддержания генетической дисперсии количественных признаков крайне поверхностно. Однако мы знаем, что разнообразии повсеместно и что большинство признаков с готовностью откликается на отбор".
В результате изучения закономерностей искусственного отбора в восьмидеся тые ; оды XX столетия были выявлены новые источники генетической изменчивости, в том числе выбывающий взрывную спонтанную мутабилыюсть механизм транспозиций. г. с. перемещения в пределах генома мобильных диспергированных генетических элементов (hcdoToff, 1984; Кайданов, 1988). Объектом, у которого впервые были открыты мобильные ысмсты, была кукуруза; на этом же объекте была разработана система выявления эффектов транспозиции ядерного и цитоплазматического геномов (Smith, 1985). Кроме подавления действия гена, в нуклеотидную последовательность которого встроился мобильный элемент, такие транспозиции усиливают кроссинговер. повышают частоту рекомбинаций (Greenblatt, 1981; Friedemann, Peterson, 1982). Установлено, что транспозиционная память обусловливает направленность внедрения мобильных элементов в определенные сайты генома, что придает спонтанному мутагенезу специфический характер, сопряженный с направлением отбора (Кайданов, 1996, с. 206, 212). По-видимому, генетическая изменчивость в течение 76 поколений отборов в популяциях кукурузы в значительной мере возобновлялась за счет транспозиций. Об этом свидетельствуют факты повышения реакции на отбор после 25 поколений отборов на и из кобел новость, отсутствие реакции между 50-м и 65-м поколением отборов на высокую масличноеи> и возобновление такой реакции в период оі 65-го до 76-го поколения, а также еще (юлес нс - кие смины Периодов отсутствия и проявления реакций при и^мснепни в обратную сторону направлений of боров (Woodworth, Leng, Jugenheimer, 1952, Dudley, w Lambert, 1969, Dudley. 1977). При сравнении одних и тех же глубокоинцухтированных родительских линий гибридов кукурузы, размножаемых селекционерами разных штатов США. было установлено, что спонтанные мутации усиливают варьирование признаков продуктивности в еще большей степени, чем изменчивость качества зерна (Fleming, 1971).
Медленное истощение генетической дисперсии при отборах на такие контролируемые большим числом генов признаки, как масличность, белковость, урожайность обусловлено не только мутационным процессом, но и постепенным высвобождением изменчивости при рекомбинациях сцепленных генов (Купцов, 1971, с. 235, Франклин, 1983). J.W Dudley (1977) показал, что при частоте генов 0,25 и 0,5 только для 50-200 локусов можно получить за счет рекомбинаций селекционный сдвиг в 20 стандартных отклонений.
Приведенные выше данные подтверждают мнение авторов, считающих преувеличенной опасность случайной утраты ценных аллелей, обеднения наследственной основы популяций, снижения реакций на отбор вследствие высокой интенсивности отборов, малого числа отбираемых особей как при селекции, так и при семеноводстве (Гуляет Шестаков, 1970, с. 37-38; Уильямсон, 1975, с. 115; Dudley, 1977; Франклин. 1983: Гуляев, 1988). В процессе отбора утрачиваются в основном нежелательные аллели и сохраняются лучшие гены в генном фонде, поскольку перевод в гомозиготное состояние желательных генов являеіся целью селекции. Поэтому А.И. Купцов (1971, с. 208-209) заключает, что снижение до определенного уровня гетерозиготности сортов перекрестноопыляющихся культур оправдано, причем оно не опасно и с точки зрения избежания инородной депрессии. поскольку іегерозис обеспечивается гетерозиготностью не всех генов., а лишь некоторых из них. Па примере ржи показано, что для закрепления ценных признаков можно в отдельных поколениях отбирать 2-5 растений (Гуляев, Долгодворова, 1976), а инбредная депрессия снимается при переопылении 10 растений (Гуляев, 1988). Для выращивания в производстве можно использовать как первое, так и последующие поколения синтетических сортов кукурузы, создаваемых из 4-8 инбредных линий (Sprague, Jenkins, 1943). Для создания таких стабильно урожайных, скрещивающихся внутри себя синтетических сортов-популяций оптимально использовать 5-6 инбредных линий (Уильямс, 1968, с. 339). } Следует учитывать, что хотя отбор малого числа особей не ограничивает селекционные сдвиги в следующих поколениях, это приводит к большим изменениям генофонда, что оказывает влияние на характер изменения комплекса признаков селектируемой понуля-| ции (Франклин, 1983).
I Таким образом, результаты многолетних опытов по отбору, а также практика рекуррентной селекции и семеноводства сортов перекрестноопыляющихся культур свидетельствует о том, что при определенных условиях отборы малого числа особей не обедняют наследственную изменчивость популяций, не вызывают инородную депрессию, но обеспечивают высокую результативность селекции за счет интенсификации отборов. Хотя многолетние эксперименты по отбору из популяций подсолнечника не проводились, нет оснований сомневаться в том, что выводы, полученные в опытах с кукурузой, рожью и другими перекрестнооплодотворяющимися видами, справедливы и для подсолнечника.
На основе таких данных и собственной практики селекции подсолнечника A.V. Vranceanu (1983) пришел к выводу: наиболее грудной и сложной проблемой является не увеличение генетической изменчивости. а разработка методов концентрации аллелей, контролирующих количественные признаки, для чего необходимо использовать широкий арсенал со-11 ремсн 11ы х методо1з.
Однако в публикациях но подсолнечнику многими авторами высказаны мнения, что индивидуальный отбор при селекции сортов приводит к снижению урожайности из-за инбредной депрессии (Жданов, 1946; Морозов, 1946, 1947, с. 246-247; Минкевич, Борковский, 1955, с. 310; Попова, Абрамова, 1968, с. 48; Анащенко, 1978а, 19786) и к полному исчерпанию наследственной изменчивости (Морозов, 1946; Анащенко, 1977, с. 7, 45; 1978а, 19786; Работа, 1985; Skoric, 1989). Полому для восстановления изменчивости рекомендуется испопьзование межсортовых скрещиваний в качестве основного метода селекции (Всцлавович, 1952; Жданов, 1952, 1957; Морозов, 1952; Прохоров, 1952). Некоторые авторы даже считают, что из-за близкородствен мости всех высокомасличных сортов работа с ними бесперспективна, что необходимо привлекать другой селекционный материал, использовать иные методы селекции (Бурлов, 1972, Анащенко, 1973, 1977, 1978а, 1980; Работа, 1985; Таволжанский, 2000, с. 276-277). При семеноводстве сортов подсолнечника для сохранения их наследственной основы и гетерозиготности рекомендуют испытывать не менее 300 потом ств элитных растений и объединять для высева в семенном питомнике резервы семян не менее 100-150 семей (Минкевич, Борковский, 195 5, с. 34 3: Хатнянский, Обыдало, 1991), что значительно превышает минимально допустимые объемы семеноводства, установленные экспериментально для других пере-крестноипыляющихся культур. К.И. Прохоров (1955) для размножения в семенном питомнике объединял семена около 50 лучших растений, но в процессе семеноводства сорта подсолнечника он обогащал его наследственную основу путем свободного межсортового переопыления (Прохоров, 1952). В.К. Морозов (1952) также закладывал питомники отбора семеноводческой элиты без пространственной изоляции от других сортов и селекционного материала подсолнечника и высевал в этих питомниках на отдельных делянках не менее 150 номеров, лучшие из которых использовал для семенного питомника.
Обсуждая проблемы сохранения наследственной основы популяций подсолнечника и их генотипического разнообразия, избежания инбредной депрессии при близкородственном разведении, ни один из цитированных авторов не обосновывает свои выводы и рекомендации вычислениями коэффициентов инбридинга и оценками вероятностей утраты аллелей при отборах малого числа элитных растений. Отсутствуют такие данные даже в обзорах по селекции и семеноводству подсолнечника (Морозов, 1977: Гундаев, 1972; Анащенко, 1977; Skoric, 1988, 1989; Skoric, Jancic, 1989). Подчеркивая важность и ого вопроса, А.А Созинов и 1О.П. Лаптев (1986, с. 90-91) утверждают, что улучшение сортов подсолнечника по урожайности и масличмости в процессе селекции и семеноводства было успешным только до тех пор, пока сам академик В.С. Пустовойт отбирал и интуитивно оценивал оптимальные с точки зрения популяционной генетики количества элитных растений. Они пишут: “Ученый все время шел на грани нежелательных отклонений. Если в популяцию включить очень мало исходных форм, она будет' терять жизнеспособность из-за близкородственного размножения, ... если исходных генотипов будет' много - трудно удержать селектируемый признак на нужном уровне”. При этом А А. Созинов и Ю.П. Лаптев также ограничились общими рассуждениями и не использовали обсуждаемые ими законы популяционной генетики для конкретной оценки коэффициентов инбридинга и вероятностей утраты аллелей при селекции и улучшающем семеноводстве сортов подсолнечника по методикам В.С. Пустовойта.
В то же время анализ метолик и результатов работы академика В.С. Пустовойча с позиций популяционной генетики важен не только для освоения наследия ученого, но и для обоснования перспектив и путей оптимизации селекции подсолнечника на современном этане. Значимость такого анализа особенно возросла после появления в литературе аыскд ^пышій о ;ом. чю и т ы высокой степени бли^кородсгвсиноон селсюпюнных сор юв подсолнечника они являючея туниками эволюции и работа с ними бесперспективна не только в направлении продолжения рекуррентной селекции, но и при использовании их в качестве исходного материала для родительских линий гибридов (Анащенко, 1977, 1978а, 16786; Рябо га, 1984).
При описании методик селекции и улучшающею семеноводства сортов подсолнечника В.С. Пустовойт' (1940, 1967) приводил подробные данные о количестве отбираемых родоначальных растений и семей их потомств. По этим данным можно вычислять коэффициенты инбридинга и оценивать вероятности утраты аллелей при отборах. Однако при таких вычислениях должны учитываться следующие обстоятельства. Важно иметь в виду, что выражение "оценка элитных растений ио потомству'; используемое даже в серьезных публикациях (например, Гундаев, 1971; Гуляев, Гужов, 1972, с. 343) не соответствует действительности. Буквальное понимание этого определения приводит к ошибочному представлению о сути индивидуального отбора в селекции растений, примером которого является описание его известными генетиками: "... сорта подсолнечника с высоким содержанием масла в семенах были получены путем индивидуального анализа отдельных растений, отбора лучших растений и скрещивания их между собой ' (Никоро и др., 1968. с 69). На самом же деле для использования в качестве родителей следующего поколения отбираются но результатам деля ночных испытаний не сами родоиачальные растения, а семьи иол>сибов, т. е. потомков одной матери и многих отцов (Купцов, 1971, с. 208). Поэтому оценка наследственных особенностей одной части семьи потомков но данным иены гания другой ее части является оценкой но полусибам. При этом на изменения параметров популяций влияет не только количество отбираемых семей, но и число отцовских растений каждой семьи полусибов, которое при панмиксии может превышать 1000 особей, поскольку отбираются корзинки, содержащие 1500-2000 семян (Пустовойт, 1967). Следовательно, при вычислениях коэффициентов инбридинга, оценках вероятностей утраты аллелей при отборах необходимо определить количество растений в нолевом посеве подсолнечника, пыльца которых опыляет цветки в корзинке одного материнского растения.
Гели бы в популяциях подсолнечника наблюдалась панмиксия, т. е. совершенно случайные скрещивания, при которых на рыльце любого цветка в корзинке с равной вероятностью попадала бы пыльца растений, растущих как рядом, 'гак и на противоположном конце ноля, то семьи полусибов получали бы с отцовскими гаметами предстали)ильную выборку генофонда всей популяции. В этом случае генофонд семьи потомков родоначального растения с большой вероятностью сохранял бы в основном все млели популяции, хотя частота некоторых из них была бы уменьшена вдвое. Однако в реальных популяциях панмиксия встречается очень редко, т. к. обычно скрещиваются особи, находящиеся поблизости друг от друга (Меттлер, Грегг, 1972, с. 89; Ли. 1978, с. 465).
S. Wright (1946) показал, что в случаях, когда дистанции рассеивания пыльцы и переноса семян меньше ареала популяции, скрещивания происходят случайно лишь в пределах панмиктических единиц, занимаемых группами растений (называемых соседствами) в непрерывной популяции. Вследствие ограниченных эффективных численностей Ме таких соседств в них наблюдается инбридинг в гем большей степени, чем меньше Ne, чго экспериментально установлено для разных видов растений (Gregorius, 1974, 1975; Coles, bower, 1976), в том числе для дикорастущего lldkmthus атшш (EHstrand. Torres. Levin, 1978 V Поэтому представление о том, что 4и... природные популяции в высокой степени гетерозиготны . . ." (Анащенко, 19786) нс соответствует действительности. Прямыми определениями установлено, что средняя гетерозигодность по !7-ти локусам у ряда перекрес гноопыляющихся видов растений равна лишь 7,8 % (Айала, 1984, с. 54).
Подразделенность популяций на панмиктические единицы приводит к тем большим генетическим последствиям, чем меньше численности панмиктических соседств (Ли, 1978, с. 478). Поэтому очень важно определить реальные дистанции потока генов в популяциях, их ограничения расстояниями разноса семян и пыльцы. Вследствие неспособности растений мигрировать, численность панмиктических соседств в их популяциях намного меньше, чем у животных. Так оценки эффективных численностей соседств у дрозофилы в разных местностях GHIA варьировали от Ne'3229 до Nc" 145780, а в популяциях разных видов растений - от Nc ~4 до Ne -282 (Грант, 1984, с. 32-3 3). Взаимное опыление у древесных пород охватывает особи в радиусе около 40 м, а у травянистых видов скрещиваются между собой растения, удаленные друг от друга не более чем на 2-4 м (Злобин. 1989, с. 6), до 99 % пыльцевых зерен рассеивается на расстоянии в 1-2 м от материнского цветка (Малецкий, 1995, с. 13 1). У вида Lkitrix ахреги (Composkud численность панмиктических соседств уменьшалась от Му2 191 до Nc:30 по мере увеличения гус тоты стояния растений на территории (Levin. Keister, 1969).
S. Wright (1946) разработал георию изоляции субпонуляций расстоянием для условий стабильного в течение ряда поколений формирования панмиктических соседств. При э том вследствие оі рапиченноі о рассеивания пыльны в популяциях перекрест ников скрещиваются родственные особи, в том числе выросшие из семян одного материнского растения (EHsirand, Torres, Levin, 1978; Малецкий, 1995, с. 131). Такие близкородственные скрещивания маловероятны в семеноводческих посевах сортов, т. к. при севе семена разных генотипов случайным образом распределяются по нолю (Френкель, Галун, 1982, с. 75). Однако результаты определений дистанций рассеивания пыльцы в посевах иере-кресгноопыляющихся культур необходимо учитывать при популяционно-генетическом анализе и оптимизации методик селекции и семеноводства популяций для оценки числа отцовских растений семей полусибов.
Рассеивание пыльцы в посевах перекресгноопыляющихся популяций культурных растений подчиняется тому же закону, который действует в естественных популяциях. Например, в посеве сафлора маркерный доминантный ген получали L2 % семян растений, растущих на расстоянии 0,6 м от донора пыльцы с этим маркером, и лишь единичные семена более удаленных растений вплоть до 34,6 м обладали маркерным геном (Deocar Pa.tiL 1976). В посевах рапса около 75 % всей пыльцы оседает в радиусе 6 м от растения-донора этой пыльны (Mcsquida, Renard, 1982). Па растениях, растущих от доноров пыльцы с генетическим маркером не далее L8 м в посевах лука репчатого и 40 м флокса, оказалось соответственно 99 и 97 % семян с маркерным геном от всего числа таких семян (Levin, Kerster, 1974). Оплодотворение семяпочек пыльцой с маркерным геном уменьшалось в 100 раз при расстоянии материнских растений от донора меченой пыльцы в посевах свеклы 6 м, кукурузы - 15 м, редиса- 48 м (Уильямс, 1968, с. 267).
Мы не нашли публикаций с данными о влиянии расстояния между донором и акцептором пыльцы в сплошном посеве подсолнечника на вероятность оплодотворения семяпочек этой пыльцой в условиях конкуренции с пыльной других растений. Много исследований было проведено лишь с целью обоснования максимального числа рядов материнской линии на участках гибридизации и требований к пространственной изоля- ции семеноводческих посевов подсолнечника. В частности, R.G. Robinson (1984) установил, что с увеличением расстояния до ряда опылителя от1 0,76 м до 30 м пропорционально снижалась завязываемость семян мужскостсрильных растений. 13.N. Patil с соавторами (1979) с помощью гена анюпиановой окраски установили, что пыльцой с таким геном оплодонюряюсь 7'1 и 72 % семяпочек растений, растущих на делянках ИһіО м на расстоянии 400 и 500 м от делянки донора маркерного гена. В опыте Л.И. Плотникова (1940) акцепторами пыльцы служили группы по 30-40 растений, в корзинках которых оставлялось по 150-200 кастрированных цветков; завязываемость семян при этом составили 3 5,6 и 35,7 % при расстояниях от источника пыльцы 400 и 5 00 м. Но наблюдениям же В.С. Пустовойта (1940), при расстояниях между питомниками 400-500 м за 25 лег было лишь три случая выщенления фуксиночных форм в посевах серосемянного подсолнечника. По данным Н.АЛибенко (1982), чужеродное перекрестное опыление материнских форм гибридов подсолнечника при изоляции от других посевов 1,2 и 2,5 км составило 18, II и 4 % соответственно.
На основе изложенных и подобных им данных нельзя определить число отцовских растений семьи полусибов-потомков элитного растения. В сплошном густом посеве подсолнечника, особенно если в корзинках много цветков, пчела перемешается на малые расстояния и переопыляет меньшее число растений, чем при расположении групп растений на больших расстояниях друг о г друга (Ellslrand, I ones, Levin, 1978). В сплошном посеве конкуренция между пыльцой разных особей также ограничивает возможности равиовероя гною переопыления всех рас гений одного поля, ч то используется в практике создания "пыльцевых барьеров’1 при работе с разными объектами (Югенхеймер, 1979, с. 408; Френкель, Галун, 1982. с. 81-82), в том числе и с подсолнечником (Рябота и др„ 1997). Рыльца мужскостсрильных растений или кастрированных цветков подсолнечника с большей всрояиюстыо воспринимают пыльцу дальних растений вследствие меньшей пыльцевой конкуренции и способности рыльца ожидать пыльцу 4-5 дней. хотя в норме оно завядает через I -2 дня (Плотников. 1940; Воскобойник, Бочкарев, Савченко. 1984).
С учетом вышеизложенно! о мы провели в 1997-1998 гг. опыты с целью оценки возможного числа отцовских растений каждой семьи полусибов при выращивании материнских растений в сплошном посеве подсолнечника на фонах обычной и ослабленной пыльцевой конкуренции. Опытные делянки располагались в пределах поля, засеянного различными сортами и гибридами подсолнечника, что создавало общий фон конкуренции между пыльцой разных генотипов Па делянках с растениями мужскостерильного простого гибрида Кубанский 48 этот фон был ослабленным, хотя и достаточным для обычной завязываемости семян в корзинках особей, в разной степени удаленных от ряда растений линии ВК 57L используемых в качестве опылителей для растений гибрида, своя пыльца которого стерильна. Имеющийся у линии ВК 571 юн Rf восстановления фертильности служил маркером для оценки вероятности оплодотворения семяпочек пыльцой этой линии в зав нс и мости от расстояния между отцовскими и материнскими растениями. С этой целью учитывали процент особей с восстановленной фергильностью пыльцы среди потомков этих матерей. Результаты представлены на графике (рис. 1).
Полученные результаты свидетельствуют1 о том, что наиболее велика вероятность оплодотворения меченой пыльцой семяпочек растений, растущих рядом с донором гена RL По мере увеличения расстояния от этих доноров до материнских растений в диапазоне от 0,7 до 4,2 м круто снижается доля их потомков, отцами которых оказались растения-доноры генетически маркированной пыльцы. Начиная с 7-го ряда материнских растений, т. с. с 4,9 м от ряда отцовских, доля потомков с восстановленной фертильностью пыльцы мало меняется но мере удаления от опылителя, оставаясь на уровне 10 %.
Этот уровень обусловлен в основном наличием гена Rf в общем фоне пыльцы на поле, засеянном сортами и гибридами подсолнечника, а частично тем, что с увеличением расстояния вероятность скрещивания резко снижается, но не достигает нуля: всегда остается малая вероятность скрещивания между особями, удаленными друг от друга (Солбриг, Солбриг. 1982. с. 169). В данном опыте (рис. 1) около половины семян в корзинках муж-скофсртильной мал ери. выросших на расстоянии 0.7 м от ряда особей донора гема Rf, завязались вследствие оплодотворения не имеющей этого гена фоновой пыльцой, т. к. * только у 52,2 % потомков оказалась восстановленной фертильность пыльцы. Обусловлено эго конкуренцией с фоновой пыльцой, поскольку в этих же условиях при выращивании между двумя рядами донора гена Rf ВК 571 одного ряда мужскостерильных растений гибрида Кубанский 48 среди его потомков оказалось 85,9 % особей с восстановленной фертильностью пыльцы.
^ 60
^"
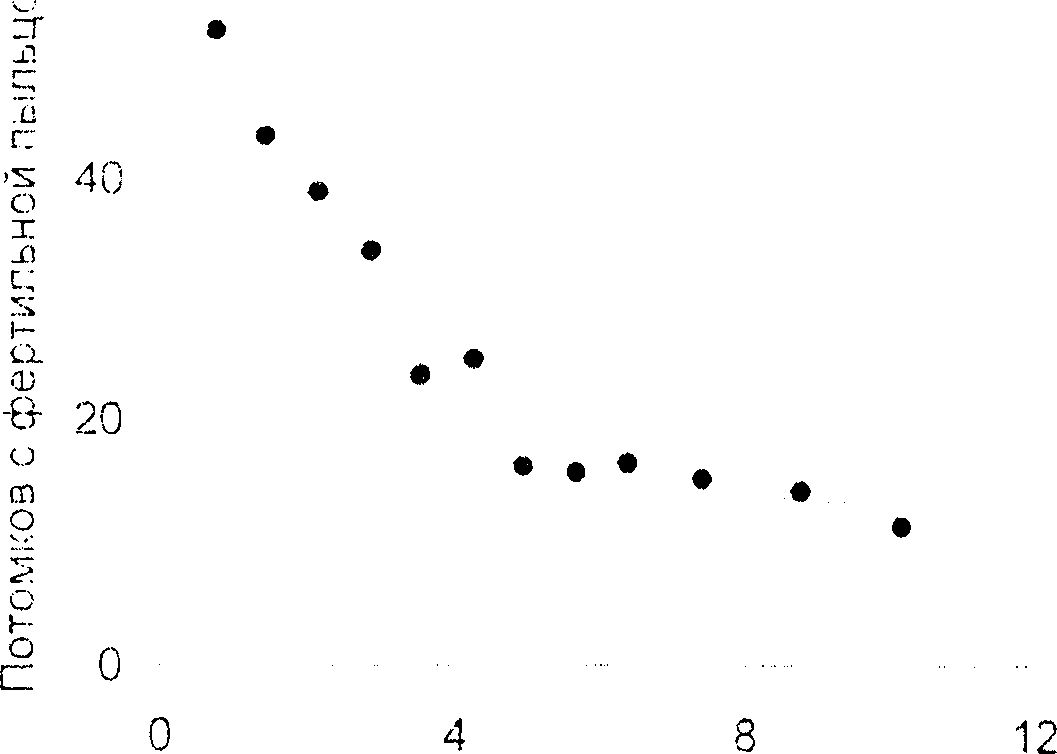
Расстояние от доноров гена Rf. м
Рисунок L Процент растений подсолнечника с фертильной пыльцой в зависимости от расстояния их мужскостерильных матерей (гибрид Кубанский 48) от донора гена Rfвосстановления (фертильности пыльцы (линия ВК 571).
Конкуренция собственной пыльцы снижает степень перекрестного оплодотворения в посевах разных видов культурных растений, в том числе масличных: у сафлора оно варьирует в таких условиях от 5 до 90 %, у подсолнечника - от 20 до 75 (Френкель. Галун, 1982. с. 40). Поэтому надо было оценить возможное число отцовских растений одной семьи полусибов в описанных выше условиях при обычной конкуренции собственной пыльцы в посеве сорта-популяции подсолнечника. Для этого использовали в качестве опылителя донор доминантного гена Т антоциановой окраски линию Сл 2349 из генетической коллекции ВНИИМК, а в качестве акцепторов генетически маркированной пыль- цы растений сорта подсолнечника Лидер. Результаты учета пропета сеянцев с антоциановой окраской среди потомков растений сорта Лидер, росших на разных расе гоя киях от растений линии ('л 2349, представлены на графике (рис. 2).
о 30*
ГО
о
X 20*
го о• со_ о_ • о 10
о•
О
0 2 4 68
Расстояние от доноров гена Т, м
Рисунок 2. Процент растений подсолнечника с антоциановой окраской в зависимости от расстояния их неокрашенных матерей (сорт Лидер) от донора антоциановой окраски (линия (’л 2349).
Вероятность оплодотворения меченой геном Т пыльцой семяпочек растений, имеющих собственную пыльцу, оказалась в среднем на 38 % ниже, чем вероятноеіь оплодотворения семяпочек мужскостерильных рас гений пыльцой с геном Rf. Пели же ряд растений copra Лидер располагался между рядами линии Сл 2349. то доля потомков, генетически меченых антоцианом, возросла только до 43,3 %, т. е. оказалась вдвое меньше процента мужскофертильных растений среди потомков растений гибрида Кубанский 48, растущих между рядами линии ВК 57 I. Эти факты свидетельствуют о том, что конкуренция собственной пыльцы в значительной мере ограничивает степень перекрестного оплодотворения в посевах подсолнечника. Однако как при обычной (рис. 2), так и при ослабленной (см. рис. I) конкуренции пыльцы обнаружено, что в корзинках подсолнечника основная часть семян завязывается в результате оплодотворения пыльцой соседних отцовских растений, растущих не далее 4,2 м от материнского растения.
Исследованиями с различными энтомофильными и анемофильными видами растений в США и других странах установлен одинаковый характер рассеяния пыльцы и вероятности оплодотворения ею по мере удаления от донора этой пыльцы. Эта зависимость описывается лсптокуртической функцией распределения, которая отличается о г нормального распределения положительным эксцессом, т. е. вокруг источника пыльцы наблюдается более высокая ее концентрация, чем можно ожидать в соответствии нор- мальным распределением (Уильямс, 1968, с. 268; Солбриг, Солбриг, 1982, с. 169-170; Френкель, Галун. 1982, с. 72; Грант, 1984, с, 30-32; Малецкий, 1995, с. 131). На этом основании S. Wright (1946) ввел понятие панмиктической единицы, г. е. части непрерывной популяции, в пределах которой происходят случайные скрещивания. Панмиктической единицей считается окружность с радиусом репродуктивной активности 2су, где а - среднее квадратическое отклонение расстояний, на которые переносится пыльца. S. Wright (1946) предложил также понятие “соседство”. В данном случае это совокупность отцов семьи потомков материнского растения, находящегося в центре окружности с радиусом 2ст. Поскольку с меньшей вероятностью пыльца переносится и с цветков растений, растущих вне этой окружности, в число особей соседства входят не 100, а 86,5 % всех отцов семьи полусибов. С учетом этого, по данным наших опытов (см. рис. 1, рис. 2), можно ориентировочно принять 4,2 м величиной радиуса репродуктивной активности в посеве подсолнечника. В этом случае площадь панмиктической единицы S= лг2г55м?. Поскольку в питомниках огбора обычно выращивают подсолнечник при густоте 2 раст./м2, величина соседства равна 110 растениям, поэтому количество отцов семьи потомков одного материнского растения не меньше этого числа. Часть семян элитной корзинки используется для анализов и высевается в питомниках испытания потомств. Поэтому можно принять, что у тех растений семьи полусибов, которые становятся родителями следующего поколения, число отцов не менее 60 особей той популяции, в которой было выращено материнское растение. В качестве минимального числа отцов семьи полусибов можно считать 4 растения, которые росли в посеве рядом с материнским растением.
Используя эти оценки числа отцов семьи полусибов подсолнечника, можно вычислять эффективные численности популяций, коэффициенты инбридинга, вероятности утраты редких аллелей при разных интенсивностях отбора, используя для этого общепринятые формулы (Никоро и др., 1968, с. 115; Ли, 1978, с. 498, 500; Рокицкий, 1978. с. 324; Солбриг, Солбриг, 1982, с. 181) и таблицу Л.А. Животовского (Яблоков, 1987, с. 182). Такие вычисления нами произведены применительно к интенсивностям отборов при обычных методах селекции и семеноводства сортов подсолнечника: использования одной семьи полусибов в качестве родоначальника сорта (Пустовойт, 1940; Жданов. 1964), объединения семян не менее 100 семей потомков элитных растений для высева в семенном питомнике при семеноводстве сорта (Хатнянский, Обыдало, 1991). Кроме этого оценены сдвиги параметров популяций для тех случаев, когда в каждом цикле рекуррентной селекции рекомендуется создавать популяции из 10 номеров, и когда для закрепления признака при отборах используются парные скрещивания под изоляторами (Пустовойт Г.В.. Хатнянский, 1980). Результаты вычислений представлены в таблице 2.
Вычисления показали, что при увеличении в 15 раз с 4 до 60-ти числа отцовских растений Nm одной семьи полусибов увеличивает эффективную численность популяции Nc только на 22,8 %. При этом на 18,6 % уменьшается коэффициент инбридинга, но остается очень низкой вероятность утраты редких аллелей. Даже при 60-ти отцах отобранной одной семьи полусибов прирост коэффициента инбридинга составил ДҒ-12,7 %, тогда как при идеальной панмиксии его величина была бы еще меньше АҒ=Ч2,5 % (Дубинин Глембоцкий, 1967, с. 390; Ли, 1978, с. 318).
Полученные результаты (табл. 2) свидетельствуют также о том, что опасность обеднения наследственной основы и снижения гетерозиготности сорта подсолнечника в процессе его семеноводства (Хатнянский, Обыдало, 1991) сильно преувеличена. При объединении 100 семей потомков элитных растений сохраняются с 95 %-ной вероятностью даже редкие аллели с частотой 1,5-2,2 %, а коэффициент инбридинга возрастает не более чем на АҒ" 0.1 6 %, что намного меньше повышения гетерозиготности популяции в каждом поколении за счет спонтанных мутаций (Гершензон, 1979, с. 215; Богоявленский и др., 1984, с. 256; Яблоков, 1987, с. 160; Глотов, 1988). В еще большей степени повышает уровень гетерозиготности сорта-популяции и восстанавливает в нем частоты редких аллелей так называемая миграция; в данном случае это случайные переопыления с растениями других сортов и падалицы подсолнечника. Насколько большим может быть влияние этого процесса можно судить по такому факту: в результате случайного опыления растений сорта Первенец пыльцой низкоолеиновых генотипов содержание олеиновой кислоты в его масле снижалось с 75-78 до 48-55% (Харченко, 1984). Даже при низких скоростях миграции до одного иммигранта на популяцию за 1-4 поколения предотвращается снижение генетической изменчивости и гетерозиготности (Меттлер, Грегг, 1972, с. 192; Франклин, 1983).
Таблица 2. Эффективные численности популяций (N^, увеличение инбредности (ДҒ,%) и минимальные частоты аллелей (Ғ^, сохраняющиеся в популяциях с вероятностью 0,95 и 0,99 при отборах разного числа родоначальных растений (N^ и числа отцов (N^ семей полусибов
|
Число отби-рае-мых расте НИЙ, Nf |
Число доноров пыльцы |
Эффективная численность популяции, Ne |
Увеличение инбредности. ДР, % |
Частоты аллелей РП)|П, сохраняющиеся при данных Nc с вероятностями |
||
|
на одну мать Nm |
Всего NB1-Nf |
0,95 |
0.99 |
|||
|
100 |
4 |
400 |
320 |
0.156 |
0,022 |
0.028 |
|
100 |
60 |
6000 |
393 |
0,127 |
0,015 |
0,022 |
|
1 |
4 |
4 |
3,2 |
15.6 |
>0,50 |
>0,50 |
|
1 |
60 |
60 |
3,93 |
12,7 |
>0,50 |
>0,50 |
|
J0 |
4 |
40 |
32 |
1.56 |
0,148 |
0.185 |
|
10 |
60 |
600 |
39,3 |
1,27 |
0,125 |
0,155 |
|
6 |
4 |
24 |
19 |
2,6 |
0,205 |
0,26 |
|
5 |
60 |
300 |
20 |
2,5 |
0,20 |
0,25 |
|
1 |
1 |
1 |
2 |
25 |
>0,50 |
>0,50 |
Даже при значительно более высокой интенсивности отборов, когда в качестве родителей следующего поколения используется 10 семей полусибов, а эффективная численность создаваемой при этом популяции достигает Ме=39 (табл. 2), величины приростов инбридинга оказываются близкими к уровню спонтанного мутационного процесса. При этом не гарантируется с вероятностью 95 % сохранение аллелей, присутствующих в популяции с частотой менее 12,5 %. Однако если при утрате таких аллелей ухудшаются хозяйственные признаки популяции, выбраковки при отборах 1 % нежелательных биотипов достаточно, чтобы избежать таких потерь (Меттлер, Грегг, 1972, с. 192).
Учитывая сложность генетической структуры сортов-популяций перекрести и ков, С. Бороевич (1984, с. 336) считает, что идентичность таких сортов при семеноводстве и... можно сохранить только в том случае, если для производства элитных семян будут отбирать такое количество растений, которое позволит охватить почти все имеющиеся генотипы”. Решить эту задачу невозможно, отбирая не только 10, но и 100 семей полусибов, поскольку “в процессе существования сорта-перекрестноопылителя в нем возникает бесчисленное количество гибридных комбинаций...” (Пустовойт, 1940). В таких популя- ниях нет постоянных юно типов, а есть постоянный генофонд, способный формирова ть бес пределы юс число возможных генотипов (Бриггс, Ноулз, 1972, с. 166). При воспроизводстве гене тичсской структуры популяций перекрсстников надо сохранять нс генотипы, а частоты генов в генофонде. а для этою рекомендуется отбирать в 2-3 раза меньшее чис.:ю растений, чем требуется для сохранения состава популяций самоопылителей (ScossirolL 1986).
Л.В. Анащенко (1978а) считает, что при использовании методов академика В.С. Пустовойта 30 лет отборов из популяций одного происхождения соответствуют 3-4 годам самоопыления, т. е. в итоге величины коэффициентов инбридинга должны были подня ться до 1 87,5 или F-93,8 %. При улучшающем семеноводстве сортов за 30 лет было проведено 15 поколений отбора, но и за этот период до такого высокого уровня инородность сортов не может подняться при приростах коэффициента инбридинга на одно поколение не только AF-~0J27 % при отборах по 100 семей, но и ЛҒ"1,27 % при отборах но И) семей в каждом поколении (см. табл. 2).
В значительно большей степени изменяются параметры при селекции сортов методом индивидуального отбора, при котором каждый сорт создается на основе семьи потомков одного родоначального растения. Этим методом были созданы, например, такие известные сорта, как Круглик Л-41 и Круглик 1846 (Пустовойт, 1940), Маяк (Жданов, 1964), ВНИИМК 1813 и ВНИИМК 1646 (Зайцев, 2003), Саратовский 169 (Купцов, 1971, с. 209-210). Переопыление потомков-полусибов одною материнского родоначального рас Юния приводиі к увеличению коэффициента инбридинга как минимум на АГ’ 12,7 % и велика вероятность утрат аллелей, содержащихся в исходной популяции с частотой более 50 °о ( табл. 2). При резком увеличении численности семьи в следующем поколении гомо-зиюзгнация практически прекращается, по степень инбридинга и частоты аллелей в генофонде сохраняются на достигнутом уровне (Сол бриг, Солбриг, 1982, с. 182). Однако продолжение отборов растений с определенными признаками способствует дальнейшему нарас танию родс твенно близких форм в сорте подсолнечника (Плачек, 1936).
В еще большей степени изменялись такие параметры популяций при использовании многократною индивидуального отбора до введения в схему селекции сортов подсолнечника питомника направленного переопыления. По описанию В.С. Пустовойта (1940), из сорта Круглик 7-15-163 была выделена семья № 3, из которой было отобрано родоначалыюе растение сорта Круглик А-41, а из него - элитное растение - родоначальное для сорта Круглик 1846, из этого сорта получена семья № 253 8, из которой также индивидуальным отбором был выведен сорт ВНИИМК 3519, а затем из него таким же методом - высокоурожайная семья № 4078. Если бы во время этой серии отборов не действовали факторы, смягчающие негативные эффекты близкородственною разведения, то такой шестикратный индивидуальный отбор довел бы в итоге коэффициент инбридинга до Ғ 50,9 %, что превышает уровень этого коэффициента.., достигаемый при однократном самоопылении (Рокицкий, 1978, с. 338).
В.К. Морозов (1946) выступал против использования при селекции подсолнечника мноюкратною индивидуального отбора из близкого по происхождению материала, считая, что такое близкородственное размножение ведет к уменьшению наследственной изменчивости и к инбредной депрессии. Однако Е.Н. Синская (1946) ко и статор о вал а не только отсутствие депрессии у сортов В.С. Пустовойта, но и отмечала даже во многих случаях большую мощность их растений по сравнению с исходными низкомасличными сортами. Так же характеризует А.И. Купцов (1971, с. 209-210) растения сорта Саратовский 169, созданного также из одной семьи. Анализируя подобные факты, он пишет: "Многократный и непрерывный семейственный отбор давал нам прекрасные сорта ржи, свеклы, подсолнечника и ряда других растений без всяких следов депрессии. Следовательно, приближение к юмозигогпости при непрерывном контроле со стороны селекционера за продуктивносзью отбираемых рас гений и качеством их продукции вполне осуществимо без депрессии из-за относительной гомозиготностгС (Купцов, 1971. с. 222).
^гог вычкд под. тверд; ла г ! с ?; гем. что такие сорта потсолнечника, -оотцмалс таким методом, как Круглик А-41, Круглик 1846. (арагонский 169 и другие находились десятилетиями в сортовом районировании и выращивались на огромных площадях.
В.С. Пустовойт (1945) наблюдал проявление депрессии во втором и последующих поколениях семей при селекции подсолнечника методом многократного индивидуального отбора. Однако при этом жизненность растений была снижена в слабой степени, а у части изучаемых семей инбредная депрессия не обнаруживалась. Отбор таких семей обеспечивал высокий уровень урожайности создаваемых сортов подсолнечника. По мнению Л.И. Гундаева (1971), такой отбор повышает уровень гетерозиса популяции, г. к. при испытаниях семей выявляются лучшие комбинации от скрещивания родоначальных растений с общим тестером - исходным родительским сортом. Па основе анализа отнята селекционной практики и данных экспериментальной генетики А.И. Купцов (1971, с. 208-209) пришел к выводу об ошибочности представления ортодоксальных дарвинистов о полной зависимости проявления гетерозиса от уровня общей гетерозигоз пости, а следовательно нет и тесной связи между величиной коэффициента инбридинга и степенью । депрессии. Он показал, что гетерозис обусловлен гегерозигоікостью лишь в некоторых
І локусах, поэтому при отборах семей полусибов из популяций перекрести икон можно добиваться гомозиготности по многим селекционным признакам без заметной депрессии жизнеспособности и плодовитости.
Важной предпосылкой проявления гетерозиса у семей полусибов является сложность их генетической структуры за счет разнообразия гамет разных отцов. Если же при парных скрещиваниях у всех потомков одной матери также и одно отцовское рас тение, то при скрещивании особей таких семей полных сибов инбредность возрастает на АЕ~25 % (см. табл. 2), поэтому в следующих поколениях наблюдается инбредная депрессия но элемен там продуктивности (Пустовойт Г.В., Хатнянский, 1980).
При селекции сортов подсолнечника методом индивидуального отбора велика вероятность утраты аллелей, имевшихся в исходной популяции (см. табл. 2). Поскольку при увеличении числа отцов семьи потомков родоначального растения с 4 до 60 эффективная численность популяции возрастает лишь с Ne-3,2 до Ne-~-3,93, в потомстве одного материнского растения даже при увеличенном числе доноров пыльцы могут нс сохраняться с вероятностью 95 % не только редкие аллели, но и содержавшиеся в исходной популяции с частотами 50 % и выше. При использовании многократного индивидушнлю-( го отбора переходить в гомозиготное состояние могут аллели многих локусов.
По мнению В.К. Морозова (1946, 1947, с. 182-183), А.В. Анашенко (1978а. 19786), АЛ. Рябо гы (1985), десятилетия использования метода индивидуального отбора 1 из близкородственных популяций подсолнечника привело к такому уровню гомозиготности сортов-популяций, который не позволяет вести селекцию на урожайность и адаптивность, обусловливает необходимость коренного изменения методики селекции и привлечение нового исходного материала. В то же время А.И. Гундаев (1971) и I). Skoric (1988) считают, что разработанные В.С. Нустовойтом методики селекции и улучшающего семеноводства сортов подсолнечника обеспечивают получение постепенных изменений в популяции при одновременном сохранении генетической изменчивости, необходимой для проведения дальнейших отборов. На основе популяционного анализа результатов работы и методики В.С. Пустовойта Е.Н. Синская (1948, с. 197) пришла к выводу, что дли тельный непрерывный отбор в одном направлении из одной экотипической популяции в определенных экологических условиях персеграиваез конституцию организмов, ускоряя за счет этою изменчивость в желательном направлении и темны селекции.
В.С Нустовойт (1940) снизал важным принципом своей работая проведение отборов селекционной зли гы из лучшею селекционного сорта последнего выпуска, мало использовал гибридизацию с материалом иною происхождения (Нустовойт; 1952). В то же время Л. А. Жданов (1946) считал необходимым свободное переопыление сортов различною происхождения для повышения наследственной изменчивости исходного материала. Однако А.И. Гундаев (1964) установил, что гетерозиготность сорта-популяции подсолнечника настолько велика, что предвари тельное усложнение генотипической структуры сорта пузом скрещивания его с другими сор гам и не только бес полезна, но и затрудняет последующий индивидуальный отбор, замедляет работу. К такому же выводу пришли R.H. Moll и C.W. Stuber (1971) при проведении индивидуального отбора полных сибов из сортов кукурузы. Анализ генетической структуры популяций подсолнечника нутом самоопыления растений показал, что даже после многолетнего непрерывною индивидуального отбора сорт Саратовский 169 и другие сорта оказались в высокой степени гетерозиготными (Плачек, 1936). Отмечая, чзо сорз' Саратовский 169 и после последующих десятилетий непрерывного отбора оставался гетерозиготным, А.И. Купцов (1971, с. 208-210) пишет, что при селекции сортов перекрестноопылителей отбор должен закреплять в семьях гомозиютность по признакам экономическою значения при сохранении гетеро тютносз и по локусам, определяющим гетерозис.
С целью подтверждения своего вывода о непригодности индивидуального отбора для селекции сортов подсолнечника на повышение урожайности и адаптивности В.К. Мо- < рогов (1947, с. 184) ссылается на результаты проводившихся в США опытов, в которых *• была установлена высокая результативность початкорядного отбора на изменение содержания масла и белка в зерне кукурузы, но неэффективность этого метода при селекции на урожайность. Однако крупнейшие специалисты США, работавшие с кукурузой, Дж.Ф. Спрэг (195 7) и Р.У. Югенхеймер (1973, с. 69. 71) писали, что метод початкорядного отбора на урожайность не дал положительных результатов не вследствие неудовлетворительности его с генетической точки зрения, а по причине непригодности методики испы таний нотомств для точной оценки урожайности. После того как Лонквист усовершенствовал этот метод га счет проведения испытаний потомств в 4-кратной повторности и использования 4-рядных делянок, его применение позволило эффективно повыгнать урожайность сортов-популяций кукурузы (Lonnquist, 1964; Webel, Lonnquist, 1967: Compton, Comstock, 1976; Compton, Bahadur, 1977).
Конечной целью и критерием успеха при селекции подсолнечника В.С. Пусто-лойт (1946, 1952, 1967) считал выведение сортов, дающих наивысший сбор масла с гектара. У создаваемых им сортов последовательно повышалась масличность семян и семянок, но пому было широко распространено представление, будто используемая при угом методика обеспечивала увеличение сборов масла только за счеі' повышения маслич-носги, но была неэффективна при селекции на урожайность. Однако детальные исследо- j вания показали, чзо причиной повышения сборов масла явилось только увеличение урожаев собственно семян (ядер семянок), т. к. при этом и их масличность возрастает из-за снижения процента белка с ростом урожая, а урожайность семянок менялась мало вследствие уменьшения сборов лузги но мере снижения лузжистости (Нустовойт, Дьяков, 1971). Поэтому высказывания о непригодности разработанной В.С. Пустовойтом методики для выведения высокоурожайных сортов являются следствием ошибочных пред- ставлений о характере происходящих в растениях подсол ночника изменений в результате селекции.
В.С. Пустовойт (1964, 1967) представлял селекцию сортов подсолнечника как циклический процесс. Каждый из таких циклов состоит из процессов рекомбинаций во время іырсэнылиння луіших геноншов. отбора элтиных растений, испытания их но томств и отбора лучших семей для переопыления в начале следующего селекпионноіо цикла (Пустовойт, 1946, 1952). Поэтому отбор селекционной элиты производился преимущественно на участках размножения лучшего селекционного сорта последнего выпуска (Пустовойт, 1940) при минимальном использовании иного материала для гибридизации (Пустовойт; 1940, 1952). Такой подход обеспечивал в каждом цикле повышение частот желательных генов в популяции. В.С. Пустовойт (1946) писал об этом: "... у этой новой линии ... созда ются но вые во зможнос ти ... как в отношении большей частоты возникновения прогрессивных гибридных комбинаций по сравнению с сортом, из которого зга линия была выделена, так и в отношении таких гибридных комбинаций с повышенными и в количественном и в качественном отношениях положительными признаками, которые в исход ном с орте не могли быть выявлены ” (подчеркнуто В.С'. Пусто войтом).
Весь этот комплекс особенностей методики В.С. Пустовойта в полной мере соответствует принципам рекуррентной селекции растений. Поэтому многие авторы справедливо считают не только эту методику селекции, но и метод улучшающего семеноводства сортов подсолнечника оригинальными вариантами простого периодического отбора, т. е. рекуррентной селекции (Гундаев, 1971; Турбин, 1971; Гуляев, Гужов, 1972, с. 23 8; Каминская, 1985, с. 56-57).
Рекуррентная селекция ведется на основе лучших сортов или синтетиков, в генофонде которых повторяющимися циклами рекомбинаций и отборов повышается концентрация желательных генов, имевшихся в исходной популяции. 'Такую методику А.С. Серебровский (1969, с. 147-148) характеризует как селекцию “в себе" в противоположность селекции с использованием гибридизации, которая может быть комбинационной или трансгрессивной (Akerman, Mackey, 1948). Поэтому нельзя считать предложенную А.В. Анащенко (1977, 1978а) методику селекции сортов подсолнечника на основе гибридизации с образцами мировой коллекции ВИР вариантом рекуррентной селекции, т. к. такие скрещивания приведу! к рассеиванию уникальных концентраций положи тельных аллелей. 'Такое предложение могло возникнуть только вследствие непонимания сути рекуррентной селекции. Об этом свидетельствует ошибочная характеристика метода В.С. Пустовойта, который А.В. Анащенко (1978а) определил как "‘вариант периодического рекуррентного отбора". По его представлению такой отбор является периодическим потому, что методика не менялась многие десятки лет, а рекуррентным потому, что улучшению подвергаются популяции одного происхождения. Это не только не отражает суть рекуррентной селекции, но и грамматически не верно, т. к. термин ‘"периодический отбор" является неудачным переводом английского словосочетания ‘"recurrent selection" (Турбин. 1971; Каминская, 1985, с. 3).
Приблизительно одинаковые темпы повышения сборов масла при реализации В.С. Пустовойтом программ селекции и улучшающего семеноводства достигались при различных интенсивностях отбора и степени воздействия на параметры популяции. При селекции была выше интенсивность отбора, но более длительные циклы. При семеноводстве - предельно короткие циклы отбора при низкой его интенсивности. По приростам коэффициентов инбридинга на один цикл эти методики различались не менее чем в 100
раз (см. габл. 2). В связи с этим важно выяснись допустимые диапазоны уровней ДҒ и их ош имальпые значения в разных программах рекуррентной селекции.
В ж и во гноводстве испол ьзуется умеренный инбридинг, коэффициент которого не превышает Г 10 %, хотя в некоторых стадах доходит до Г з25 % и выше (Брюбейкер, 1966, с. 55). При рекуррентной селекции кукурузы считается, что при достаточно полном перекрестном опылении растений в предыдущей популяции очередной синтетик следует формировать из 1(1 линий, что обеспечивает максимально допустимую скорость инбридинга Аг 2,8 % на цикл (Sprague. Miller, Brimhall, 1952; Спрэг, 1957). При формировании популяций подсолнечника из семей полусибов близкие приросты в каждом цикле коэффициентов инбридинга AF-2,5 % и ЛҒ-2,6 % можно обеспечивать при переопылениях 5 или 6 семей при 60 или 4 отцах каждой семьи соответственно (табл. 2).
Даже после 6-кратного индивидуального отбора с приростами в каждом цикле коэффицента инбридинга не менее АҒ^12,5 % В.С. Пустовойт (1940) выделял высокоурожайные семьи без признаков депрессии. Однако с 1945 г. он начал писать о наблюдаемых им фактах инбредной депрессии и о мерах, которые он вынужден был принимать для избежания негативных последствий близкородственных скрещиваний (Пусто войт, 1945. 1946).
Возможность разработки таких мер становится понятной, если учесть, что приведенные выше оценки инбридинга и вероятностей утраты редких аллелей вычисляются но формулам, отражающим сдвиги параметров идеальных панмиктических популяций без учет дефектов мутаций, миграций, отбора, неполного перекрестного оплодотворения. Вследствие небольшой частоты спонтанных мутаций в малых популяциях их влияние на степень гетерозиттносзи и генные частоты значительно меньше по сравнению с отбором, величиной эффективной численности популяции, степенью панмиксии (Меттлер. Грегі , 1972, с. 134,191). Основным механизмом миграций (потока генов) при селекции и $ семеноводстве подсолнечника является опыление корзинок селектируемых популяций пыльцой других сортов или семей полусибов. Чем интенсивней миграции, тем эффективней восстанавливается гетсрозиготность популяций, но снижается эффективность отборов. При невысокой скорости миграций особенно велики возможности ограничения гомозиготизации популяций отбором в пользу гетерозигот (Никоро и др., 1968, с. 137; Свирежсв, Пассков, 1982, с. 185). При численности популяции 50 особей достаточно 1 %-ного отбора для предотвращения случайного закрепления аллелей (Меттлер, Грегг, 1972, с. 189,192). На примере популяций Melianthus annuus M.S. Elistrand, A.M. Torres и I).A. Levin (1978) показали, что чем меньше гетсрозиготность проростков, тем в большей мерс естест венным отбором уменьшаются коэффициенты инбридинга взрослых растений но сравнению с величинами Ғ у проростков, причем отрицательные величины разницы достигаю! ЛҒ 46 %. Выбраковка слаборазвитых растений и низкоурожайных семей потомков также поддерживает гетсрозиготность популяций подсолнечника при их селекции и семеноводстве. За счет этого отбор обеспечивает также накопление в популяции ншов самонссовместимости, которые являются важным фактором поддержания высокого * уровня генетической гетерозиготности (Френкель, Галун, 1982. с. 237), В создаваемых из инородных линий синтетических популяциях подсолнечника из-за низкой частоты генов самонссовместимости велика вероятность самооплодотворения (Robinson, 1980). Поэтому при рекуррентной селекции на основе таких синтетиков в них трудно обеспечивать перекрестное онлодоторсние особей во всех комбинациях, что не только снижает эффективность рекомбинаций, но и увеличивает коэффициенты инбридинга в соответствии с формулой (Nei. Syakudo, 1958):
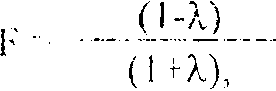
i.ic ? - чистота перекреишиго он.юдотворения. Этот фактор особенно сильно влияеі на юмффицен! инбридинга при небольших значениях численности кяі). ыцш; (Корее гышевский, 1976).
Для рекуррентной селекции особенно важны процессы рекомбинации по двум причинам. Во-первых. они противодействуют истощению запаса генетической изменчивости популяции, т.к. даже при небольшом числе гетерозиготных локусов обусловливают возможность образования огромного множества генотипических вариантов. Во-вторых, в процессе мейитической рекомбинации возникают уникальные генные комплексы, что обеспечивает возможность рекуррентным отбором концентрировать ценные гены в іено-фонде. Как и в исследованиях с кукурузой, в программе рекуррентной селекции подсолнечника также установлены высокие реакции на отбор и одновременно не только сохранение наследственной изменчивости в популяции, но даже удвоение коэффициента наследуемости (Mishra, Chaubey, 1987).
Первой мерой, принятой с целью избежания отрицательных последствий родственного разведения при селекции подсолнечника, был отказ от многократного индивидуального отбора и его 'замена однократным после переопыления между растениями лучших ногомств в каждом цикле (Нустовойт, 1946). Однако при этом еще использовалась утвержденная в 1939 г. схема индивидуального отбора, в которой отсутствовало такое звено как питомник направленного переопыления. Это звено было введено позже путем двух важных изменений методики закладки пространственно удаленных изолято-, ров (Нустовойт, 1952). Во-первых, число рядов кукурузы между делянками, засеянными ? семьями потомков селекционной элиты подсолнечника, было сокращено с 3-5 до одного ряда, что резко усиливает взаимное переопыление семей (см. рис. L рис. 2), увеличивает обмен генами вследствие миграций. Во-вторых, в отличие от прежних пространственных изоляторов в питомники направленного переопыления высевали семьи иолусибов не одновременно с их испытаниями в питомнике оценки ногомств, а после такшх испытаний, по результатам которых отбирались и переопылялись между собой только самые лучшие семьи. Это позволило до минимума снизить отрицательное влияние интенсивных миграций генов на эффект ивность отборов.
Принятие описанных мер для нейтрализации отрицательных проявлений родственного размножения при использовании в селекции подсолнечника высоких интенсивностей отборов позволило еще многие годы эффективно применять индивидуальный отбор для увеличения сборов масла. В то же время следует признать ведущую роль отбо-; ра в поддержании гетерозиготности популяций и наследственной изменчивости в них. Это доказывается высокой результативностью применения интенсивного многократного индивидуального отбора до введения в схему селекции питомника направленного । переопыления. Следует особо подчеркнуть, что высокая эффективность проводимых В.С. Нустовойтом отборов была возможна только при надежных оценках наследст венных достоинств изучаемых генотипов. Методику таких оценок В.С. Нустовойт (1940) разрабатывал 15 лет и лишь на этой основе были обеспечены успехи в селекции и улучшающем семеноводстве сортов подсолнечника. Принципиально важно то, что разработанные им методы были надежными нс только при деляночных испытаниях семей потомков элит. В.С. Нустовойт (1940) особенно подчеркивал важность точной оценки генотипов отдельных растений по их фенотипам, т. к. возникающие в каждом поколении перекрестно- опыляюшейс я популяции уникальные генотипы не воспроизводя гея в последующих поколениях.
Таким образом, рассмотрение методов селекции и улучшающего семеноводства сортов подсолнечника и других пере крестников с позиции популяционной генетики и биологии популяций позволяет сделать следующие выводы. При надежной идентификации продуктивных генотипов рекуррентная селекция может быть эффективной при использовании широкого диапазона интенсивностей отборов. Применявшиеся минимальные интенсивности отборов семеноводческой элиты практически не влияли на степень инородности популяций и могли приводить к утрате только очень редких аллелей. При использовании же в селекции сортов подсолнечника однократного и. особенно, многократного индивидуального отборов приросты коэффициента инбридинга были выше АР 12,5 % на один цикл, была высока вероятность утраты даже аллелей, содержащихся в популяциях с высокой частотой. Такая высокая интенсивность отборов позволяет эффективно концентрировать желательные гены в генофонде популяции при условии высокой надежности как оценок семей потомков элит по результатам деляночных испытаний, так и идентификации генотипов по их фенотипам при отборах элитных растений. Кроме этого, должны приниматься эакже другие меры минимизации инородной депрессии и него-шепия генетической изменчивост и в популяциях.
