Возрастная изменчивость одонтологических признаков рыжей полевки (Clethrionomys glareolus Sсhreber)
")
Автор: Емельянова Алла Александровна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 1, 2005 года.
Бесплатный доступ
При проведении фенетических исследований природных популяций важно исключать признаки, подверженные возрастной изменчивости. Изучалось наличие возрастной составляющей в полиморфической изменчивости рисунка жевательной поверхности моляров европейской рыжей полевки, являющейся представителем корнезубых полевок. Обнаружено, что большинство вариаций, отражающих особенности формы вершин углов и сторон эмалевых призм, а также количество замкнутых дентиновых полей, тесно связано с процессами формирования и стирания коронки зуба.
Короткий адрес: https://sciup.org/146116031
IDR: 146116031 | УДК: 59.08
Age variation of the teeth characters of the bank vole (Clethrionomys glareolus Sсhreber)
On the course of the phenetics research in the natural populations it is crucial to exclude characters influenced by variability due to the age. The presence of the age component in the polymorphic variability of chewing surface of molars of the root-toothed Bank Vole has been studied. It has been found, that the majority of variations connected to the morphology of angles and sides of the enamel loops as well as the number of closed dentin fields are intimately related to the process of teeth formation and wearing.
Текст научной статьи Возрастная изменчивость одонтологических признаков рыжей полевки (Clethrionomys glareolus Sсhreber)
С начала 70-х гг. XX в. в популяционно-морфологических исследованиях интенсивно развивается новый методологический подход к изучению изменчивости признаков – фенетический, базирующийся на известном факте фенотипического проявления генотипа и концепции фена как признака-маркера генотипа [1;8;9]. Фенетика популяций возникла на основе понятия «эпигенетический полиморфизм» (полиморфическая изменчивость) – это прерывистые, обычно неметрические вариации в выражении наследственных признаков. Фенетические исследования позволяют решить ряд популяционных задач, таких, как обнаружение границ между популяциями и их группами, установление уровня сходства и иерархии популяционных группировок, выявление закономерностей географической изменчивости, выявление действия естественного отбора и мн. др. [11;12].
У млекопитающих в качестве фенов часто используются дискретные вариации структурных элементов зубной системы, изучение которых представляет особый интерес, так как форма жевательной поверхности моляров, с одной стороны, находится под генным контролем, с другой – является признаком пищевой адаптации [2;15;17].
Для того чтобы признаки позволяли легче выявлять общие черты популяционных особенностей, следует исключить те из них, которые связаны с полом и возрастом. Об отсутствии полового диморфизма по набору фенов зубной системы и распределению частот их встречаемости неоднократно упоминалось в литературе. Изучению же возрастной изменчивости рисунка жевательной поверхности моляров полевок обычно не уделяется должного внимания в фенетических исследованиях, в то время как Н.Н. Воронцов указывает на значительное влияние на строение жевательной поверхности особей питания и манеры обработки кормов [2].
На изменения формы жевательной поверхности особенно влияет наличие или отсутствие корней зубов. Известно, что для некорнезубых полевок характерен постоянный рост зубов, обеспечивающийся функционированием в течение всей жизни эмалевого органа. Из-за отсутствия корней зубы у этих полевок всю жизнь имеют призматическое строение, и стирание жевательной поверхности мало влияет на её рисунок. У корнезубых полевок, к которым относится и рыжая полевка, рисунок жевательной поверхности меняется с возрастом, что позволяет использовать морфологию зубов для определения их возраста [3;6;10;13;14;16].
С целью проверки наличия возрастной составляющей в полиморфической изменчивости одонтологических признаков европейской рыжей полевки были исследованы серии зверьков (195 особей), отловленных летом и осенью 1998 г. в Кали- нинском районе Тверской области. Зверьки распределялись на пять наиболее распространенных в зоологических исследованиях возрастных групп: аdultus (Ad) – взрослые, перезимовавшие полевки; subadultus3 и abulescens (Sb3 – Ab) – способные к размножению сеголетки, частично достигшие размеров взрослых; subadultus2 (Sb2) – молодые неполовозрелые зверьки, ведущие самостоятельный образ жизни; subadultus1 (Sb1) – совместно живущий молодняк; subadultus0 (Sb0) – выходящий из нор, но живущий вместе с самкой молодняк [3].
При изучении строения коренных зубов рыжей полевки за основу были взяты признаки и их вариации (фены), предложенные Н.И. Лариной и И.В. Ереминой [5], а также морфотипы по И.В. Ереминой [3]. Выделение фенов проводилось по рисункам, выполненным рисовальным аппаратом РА-6, установленным на микроскопе при постоянном увеличении. Зарисовывались правые и левые моляры верхней и нижней челюсти. Всего в работе использовано 1170 рисунков, с помощью которых было выявлено 296 вариаций 109 признаков элементов жевательной поверхности моляров. Оценка достоверности различий возрастных групп по частотам вариантов признака проводилась методом Х2, констатация различий между выборками происходила при уровне значимости р≤0,05 [7].
Ниже приводятся наиболее яркие примеры возрастной изменчивости ряда признаков зубной системы рыжей полевки.
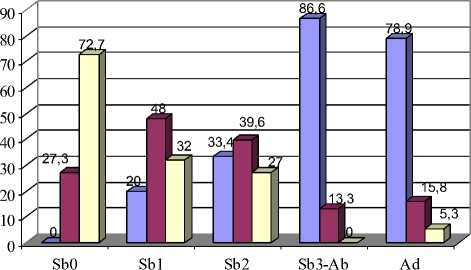
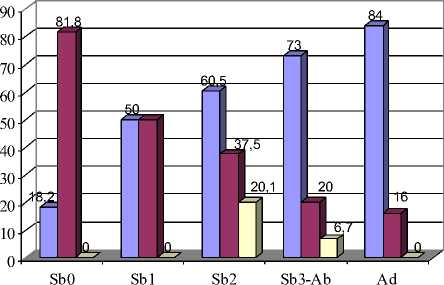
Анализ особенностей строения первого верхнечелюстного моляра (М1) у полевок разного возраста показал, что признак, отражающий форму передней стороны непарной петли, подвержен возрастной изменчивости (рис. 1; 5,А). Изучая встречаемость вариаций данного признака, можно отметить постепенность изменения их доли от возрастной группы Sb0 к группе Ad. При этом равномерно округлая, выпуклая передняя сторона (вариация 1\1) встречается преимущественно в старших возрастных группах – у полувзрослых и перезимовавших зверьков (86,6 и 78,9 % соответственно). Плоская форма (вариация 1\2) характерна для молодых полевок. Особенно велик процент этого фена в возрастных группах Sb 1 и Sb 2 – 48,0 и 39,6 %. Частота встречаемости передней стороны непарной петли с выемкой или желобком (1\3) максимальна в возрастной группе Sb0 – 72,7 %. Различия по частоте встречаемости данных фенов между зверьками из старших и младших возрастных групп достоверны (р≤0,05; 0,01).
Рассматривая возрастную изменчивость признаков характера замкнутости зубных полей (13, 14, 15 и 16-й), можно отметить явную тенденцию к слиянию дентиновых полей у сеголеток возрастной группы Sb0 и у перезимовавших полевок. В целом у молодых полевок по мере взросления наблюдается увеличение доли моляров с замкнутыми пространствами (р≤0,05; 0,01; 0,001) (рис. 1; 5,Б).
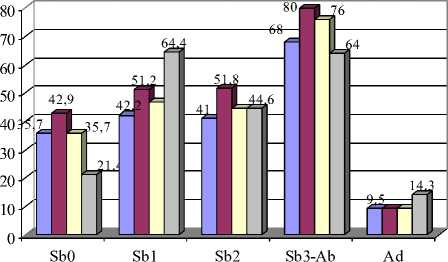
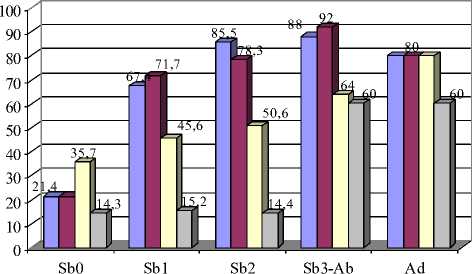
Вариации 18\2 и 19\1 второго верхнечелюстного моляра (М2) соответствуют симметричной форме протокона, только признак 18 означает, что вершины выступающих углов закругленные, а признак 19 – заостренные (рис. 2). В обоих случаях симметричная передняя непарная петля оказалась более характерна для молодых полевок. Закругленная вершина внутреннего выступающего угла (19\2) минимальна в возрастной группе Sb0 (33,3 %) и максимальна у перезимовавших зверьков (90 %) Кроме этого, частота встречаемости срезанной вершины внутреннего выступающего угла протокона (19\3) у взрослых полевок достоверно меньше по сравнению с другими возрастными группами (р≤0,05; 0,01) (рис. 6,А).
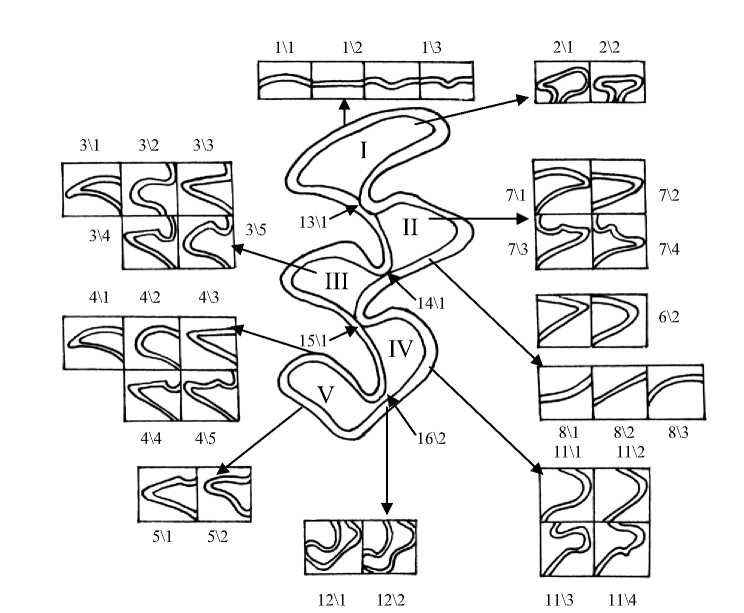
Рис. 1. Жевательная поверхность М1 и некоторые вариации её элементов Здесь и далее приводятся рисунки моляров правой стороны тела:
I – передняя непарная петля (передний воротничок); II – V – основные треугольники (протокон, гипокон, паракон, метакон)
17\1 17\2 17\3 17\4
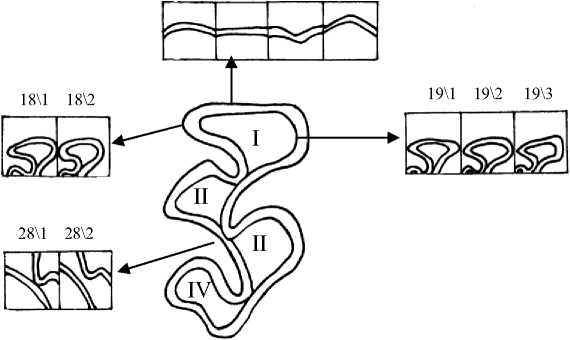
Рис. 2. Жевательная поверхность М2 и некоторые вариации её элементов I – передняя непарная петля (протокон); II – IV основные треугольники (гипокон, паракон, метакон)
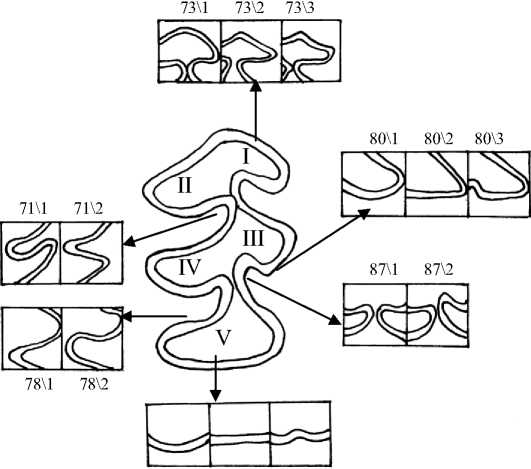
83\1 83\2 83\3
Рис. 3. Жевательная поверхность М 2 и некоторые вариации её элементов I – параконид; II – метаконид; III – протоконид; IV – энтоконид; V – гипоконид (задняя непарная петля)
88\1 88\2 88\3
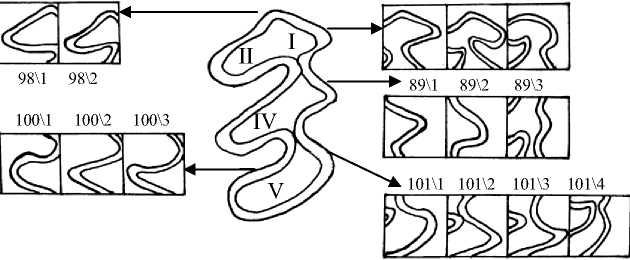
Рис. 4. Жевательная поверхность М3 и некоторые вариации её элементов I – IV – основные треугольники (параконид, метаконид, протоконид, энтоконид);
V – гипоконид (задняя непарная петля)
Возрастные изменения формы основных треугольников М2, а также формы их передних сторон сходны с таковыми признаками основных треугольников М1 и заключаются в следующем: выпуклая, округлая и округлая со срезанной вершиной форма основных треугольников в основном обнаружена в возрастных группах Sb3 и Ad (например, вариации 3\1, 3\2 и 3\5 соответственно); процент треугольной и срезанной формы велик у сеголеток, особенно в возрастной группе Sb 1 (вариации 3\3, 3\4) (рис. 1).
Различия встречаемости таких вариаций формы передней стороны метакона М2, как округлая, срезанная и вогнутая сторона (21\2, 21\3, 21\4), в разных возрастных группах подтверждаются статистически (р≤0,05; 0,01; 0,001) (рис. 6,Б).
Среди элементов жевательной поверхности третьего верхнечелюстного моляра (М3) возрастной изменчивости подвержены: форма основных треугольников, их передних и задних сторон, выступающих углов.
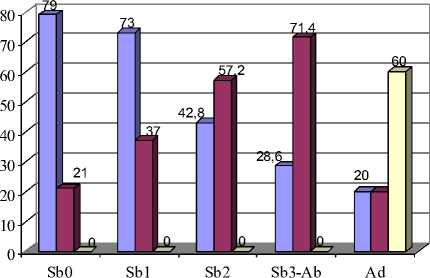
Характер возрастных изменений признаков, описывающих форму основных треугольников и число замкнутых пространств М 1 , М 2 (для М 2 исключение – признаки характера замкнутости зубных полей) и М 3 , также сходен с рассмотренной ранее возрастной изменчивостью подобных признаков на примере моляров верхней челюсти. На рис. 7,А представлена возрастная изменчивость встречаемости выпуклой, плоской и срезанной формы задней стороны протоконида М2 (79\1, 79\2, 79\3 соответственно) (р≤0,05).
Исследуя фенетические комплексы М 2 и М 3 у рыжих полевок разного возраста, обратим внимание на ряд признаков, выделенных для жевательной поверхности этих моляров и ранее нами не рассматриваемых.
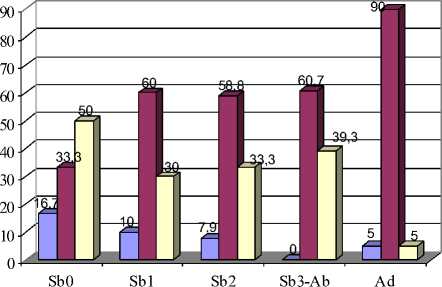
Признаки 69-72 второго нижнечелюстного и 92-95 третьего нижнечелюстного моляров характеризуют ориентацию вершин внутренних входящих углов относительно продольной оси зуба, и все они изменяются с возрастом (рис. 3). У полувзрослых и взрослых зверьков значительно чаще, чем у молодых полевок, вершины входящих углов направлены под углом к продольной оси зуба, что подтверждается статистически (р≤0,05; 0,01; 0,001) (рис. 7,Б).
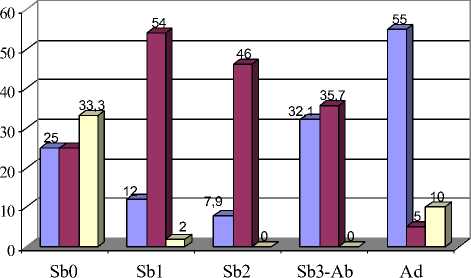
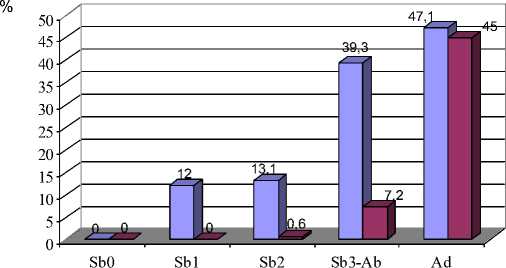
Признаки взаиморасположения пар входящих углов у М 2 и М 3 (86, 87, 110, 111) также обнаруживают возрастную изменчивость, наиболее заметную у третьего нижнечелюстного моляра и заключающуюся в увеличении частоты встречаемости супротивного положения первой и второй пар входящих углов у зверьков старшего возраста (Sb 3 и Ad) (р≤0,05, 0,01, 0,001) (рис. 8, Б).
Признаки 84-85 и 107-109 отражают число изолированных дентиновых полей. Интерес представляет факт, что для М2 (84 и 85) отмечено постепенное нарастание процента зубов с разделением дентиновых полей от возрастной группы Sb0 к группе Ad. Это отличается от зафиксированных ранее для большинства моляров возрастных изменений подобных признаков: от возрастной группы Sb 0 к группе Sb 3 наблюдается направленная изменчивость в сторону увеличения доли зубов с разделением зубных полей, затем данная тенденция сменяется увеличением встречаемости моляров со слиянием дентиновых полей в возрастной группе Ad.
Приведем предположительную схему возрастных изменений признаков характера замкнутости зубных полей. У сеголеток, относящихся к возрастной группе Sb 0 , зачастую дентиновые поля образуют единое пространство в связи с только начавшимся процессом формирования призм моляров. В онтогенезе происходит утолщение слоя эмали и дифференциация жевательной поверхности с четким обособлением репаративного дентина от первичного. При этом происходит изменение рисунка жевательной поверхности, заключающееся в углублении входящих углов, что влечет за собой увеличение количества замкнутых полей. Максимальная доля вариаций разделения зубных пространств найдена у сеголеток группы Sb2 и
А
1\1
1\2
1\3
Возрастные группы
Возрастные группы
Б
13\1
14\1
15\1
16\1
Возрастные группы
Рис. 6. Возрастная изменчивость частот встречаемости фенов М2:
А – форма вершины внутреннего выступающего угла протокона; Б – форма передней стороны метакона
19\1
19\2
19\3
Рис. 5 Возрастная изменчивость частот встречаемости фенов М1:
А – форма передней стороны непарной петли; Б – признаки разделения 1-4 дентиновых полей
А %
Возрастные группы
Б
21\2
21\3
21\4
Возрастные группы
А
□ 79\1
□ 79\2
□ 79\3
Возратные группы
Б
□ 69\1
□ 70\1
□ 71\1
□ 72\1
Возрастные группы
Рис. 8. Возрастная изменчивость частот встречаемости фенов М3:
А – форма вершины параконида; Б- встречаемость супротивного расположения 1-2 пар входящих углов
Рис. 7. Возрастная изменчивость частот встречаемости фенов М2:
А – форма задней стороны протоконида; Б – ориентация вершин входящих углов относительно продольной оси зуба
А
□ 88\1
□ 88\2
□ 88\3
Возрастные группы
Б
□ 110\1
□ 111\1
полувзрослых рыжих полевок, моляры которых имеют вполне сформированную коронку, а процесс образования корней находится на начальной стадии (в отловленных нами сериях зверькам из возрастной группы Sb 3 -Ab – 3-4 месяца). У перезимовавших полевок в возрасте 9 месяцев имеются вполне развитые корни, длина которых около 60 % общей высоты зуба [3]. На этом этапе коронка ещё сохраняет призматическое строение. Дальнейшее удлинение корней приводит к тому, что межкорневой свод начинает примыкать к основанию коронки, одновременно призмы коронки стачиваются. В результате этих процессов дентиновые поля вторично сливаются, а выходящие углы и стороны основных треугольников и непарных петель приобретают округлую форму. При этом у наиболее интенсивно изнашивающихся задних коренных зубов [6] наружные выступающие углы снашиваются вплоть до полного их отсутствия (вариации 63-64\3, 88-89\3) (рис. 4; 8,А).
Отступления от данной схемы возрастных изменений рисунка жевательной поверхности, отмечаемые для некоторых моляров, могут быть связаны с разной нагрузкой, испытываемой коренными зубами и их частями при обработке пищи, или с особенностями строения конкретного моляра. Косвенным доказательством этого предположения может служить наблюдение, что вышеуказанные возрастные изменения признаков характера замкнутости зубных полей наиболее ярко выражены и раньше происходят (уже в возрастной группе Sb 3 начинается уменьшение доли вариаций разделения дентиновых пространств) у моляров, испытывающих наибольшую нагрузку, которыми являются М3 и М1 [3;6]. При совершении жевательных движений у М2 максимальная нагрузка падает на ближайшие к М3 основные треугольники – паракон и метакон, выраженное вторичное слияние дентиновых полей которых найдено у полувзрослых особей. Для других двух признаков характера замкнутости зубных полей М2 не обнаружено такой резкой возрастной изменчивости: встречаемость случаев разделения дентиновых полей с возрастом изменяется незначительно, и в группе перезимовавших полевок лишь на 2-5 % ниже, чем у полувзрослых зверьков.
Отметим, что статистически значимые различия возрастных групп по признакам, относящимся к элементам рисунка жевательной поверхности М3 и М 1 , изменчивость которых явилась основанием для выделения морфотипов этих коренных зубов И.В. Ереминой [3] (форма наружной и внутренней стороны талонуса и пара-конидного отдела), нельзя в полной мере отнести на счет возрастной изменчивости, так как распределение частот встречаемости вариаций данных признаков в возрастных группах хаотично и не образует определенной тенденции. Это свидетельствует о других причинах преобразований рисунка жевательной поверхности.
В литературных источниках указывается, что по каждому признаку парако-нидного отдела М 1 выделяются фены, контролируемые одним геном. Анализ признаков зубной системы на черепах полевок строго фиксированного возраста позволил в пределах инбредных линий обнаружить некоторые особенности формы медиальной и латеральной сторон жевательной поверхности М 1 , присущие только ювенильным, полувзрослым и взрослым особям [4]. Наши данные вполне согласуются с литературными и позволяют сделать вывод о большей наследственной обусловленности формы талонидного комплекса и параконидного отдела по сравнению с формой основных треугольников и задней непарной петли, в фенотипической изменчивости которых обнаружена значительная доля возрастной составляющей.
Таким образом, из результатов исследования следует, что значительная доля изученных дискретных признаков зубной системы рыжей полевки в той или иной степени подвержена возрастной изменчивости (около 80 признаков из 111). В основном это признаки, описывающие форму вершин углов эмалевых призм, форму сторон основных треугольников и количество замкнутых дентиновых полей. Возрастные изменения рисунка жевательной поверхности связаны с процессами фор- мирования и стирания коронки зуба. В популяционных исследованиях возможно использование данных фенов либо в одновозрастных группах, либо при объединении смежных возрастных групп, различие между которыми по частоте встречаемости вариаций статистически недостоверно. Однако в этом случае фенокомплексы скорее будут отражать специфику кормовой базы, влияющую на степень снашивания моляров, а не генетические особенности группы особей.
On the course of the phenetics research in the natural populations it is crucial to exclude characters influenced by variability due to the age. The presence of the age component in the polymorphic variability of chewing surface of molars of the root-toothed Bank Vole has been studied. It has been found, that the majority of variations connected to the morphology of angles and sides of the enamel loops as well as the number of closed dentin fields are intimately related to the process of teeth formation and wearing.
