Выделение генотипов сои со скрытой полулетальностью в условиях эдафического стресса..

Автор: Мошненко Е.В.
Статья в выпуске: 1 (136), 2007 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/142150794
IDR: 142150794
Текст статьи Выделение генотипов сои со скрытой полулетальностью в условиях эдафического стресса..
Введение. Известно, что соя является строгим самоопылителем (Корсаков, Мякушко, 1975; Лещенко и др., 1987). Это не позволяет использовать в селекции такой эффективный механизм повышения продуктивности, как гетерозис. Поэтому одним из важнейших этапов селекции у сои является подбор родительских пар для скрещивания (Zelentsov et al., 2004).
Важный вклад в дальнейшее развитие признаковой концепции подбора родительских пар может внести теория компенсационных комплексов генов, выдвинутая В.А. Струнниковым в 1974 г. (Струнников, 1983; Кайданов, 1996). Согласно этой теории полулетальные мутации, не затрагивающие существенно репродуктивные функции организма, за счет горизонтального переноса генов и рекомбинационных процессов в мейозе способствуют накоплению комплекса доминантных и полудоминантных генов (ККГ), компенсирующих негативный эффект мутаций.
При расщеплении гибридов, полученных с участием источников ККГ, образуются формы с отсутствием полулетальной мутации и наличием активных компонентов ККГ, обеспечивающие и сохраняющие ККГ-эффект. В таких генотипах, при отсутствии вредной мутации, компенсирующий эффект коадаптированного блока генов ККГ реализуется в положительном аддитивном воздействии на весь организм, повышая его общую жизнеспособность и продуктивность (Ze-lentsov et al., 2004).
Ранее было установлено, что в посевах сортов – носителей полу-летальной мутации и ККГ-комплекса встречается незначительное количество (0,1-8,0 %) субвитальных растений. Тщательное удаление таких растений не приводит к уменьшению доли полулетальных растений в следующих поколениях. Более того, в большинстве случаев из семян, собранных на таких растениях, развиваются вполне нормальные особи (Зеленцов, Пендинен, 2002).
Генотипы сои с наличием в их геноме полулетальной мутации и активным ККГ-комплексом представляют собой ценный исходный материал для селекции. Практическим результатом использования идентифицированных источников ККГ в скрещиваниях явилось выведение в отделе сои ВНИИМК высокопродуктивных и экологически пластичных сортов Вилана, Рента, Дельта, Лакта (Зеленцов, 2005). В связи с этим особую актуальность приобретает дальнейший поиск и выделение новых источников ККГ для их вовлечения в селекционный процесс.
Материал и методы . Исследования проведены в 2000-2005 гг. во ВНИИ масличных культур. Материалом для морфологических и цитогенетических исследований служили сорта и линии сои селекции ВНИИМК, а также образцы мировой коллекции сои ВИР и сорта других научных учреждений. Посев, фенологические наблюдения, оценку и анализы селекционного материала проводили по методикам Государственной комиссии по испытанию и охране селекционных достижений и принятым во ВНИИМК.
Дополнительно исследовали около 700 сортообразцов коллекционного питомника, расположенного на участке поля с уплотненной и нарушенной структурой почвы.
Подсчёт и оценку нормальности прохождения метафазной и анафазной стадий митоза проводили в апикальных частях корешков длинной от 0,3 до 0,8 мм у 25 проростков каждого сорта на световом бинокулярном микроскопе при увеличении X500-1000. Для приготовления и окрашивания временных давленых препаратов использовали стандартную методику (Паушева, 1988), адаптированную для сои. Временные препараты окрашивали ацетолак-моидом.
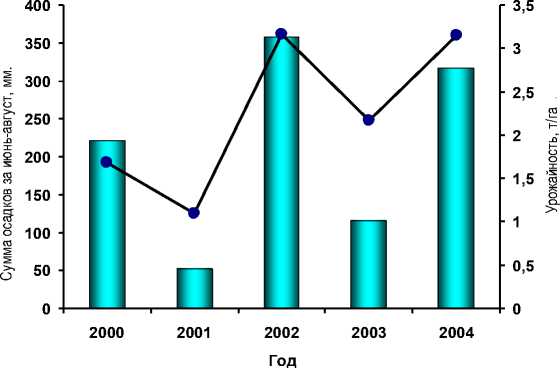
Результаты и обсуждение. Полевые наблюдения в течение 2000-2004 гг. показали, что на сортах Фора и Веста, ранее уже идентифицированных, как источники скрытой полулетальности, доля полулетальных растений возрастает в годы, отличающиеся засушливостью и связанной с ней низкой урожайностью сои (рис. 1).
Полученные данные позволили нам сформулировать гипотезу о зависимости эффективности действия коадаптированного блока ККГ при нейтрализации полу-летальной мутации от условий роста и развития растений. Предполагалось, что ухудшение условий внешней среды может привести к снижению эффективности ККГ-комплекса и как следствие, увеличению частоты встречаемости полулетальных растений.
С целью проверки данной гипотезы, а также для выявления новых источников ККГ для создания новых сортов исследовали около 700 коллекционных сортообраз-цов в условиях эдафического стресса (на низинном участке поля с уплотненной нарушенной структурой почвы), обеспечивающем стрессовые условия для роста и развития сои как в засушливые, так и во влажные годы. Параллельно резервы семян всех изучаемых сортообразцов высевались в опти-мальных почвенных усло-вииях. В качестве тест-контроля наличия стресса использовали сорт сои Фора, известный как носитель полулетальной мутации.
Визуальная оценка образцов, выращенных на участке с уплотнённой почвой дополнительно к сорту Фора, позволила предварительно выделить еще 4 сортооб-разца с выраженными полуле-тальными формами. При этом в оптимальных почвенных условиях полулетальные растения в выделенных сортообразцах отсутствовали (табл. 2).
Как следует из данных, представленных в табл. 2, частота появления полулетальных растений контрольного сорта Фора в условиях эдафического стресса увеличилась практически в 30 раз.
Эти же неблагоприятные почвенные условия спровоцировали появление слабофертильных растений и у ряда других сортооб-разцов. В частности, частота проявления пониженной фер тильности полулетального типа варьировала от 7,14 – у сорта
Приморская 56 до 20,00 % – у сорта Приморская 93. Аналогичная реакция на эдафический стресс была обнаружена и у коллекционного образца, принадлежащего к другому виду сои – Glycine gracilis Skvortz., достигшего максимального в опыте значения – 22,20 %. При этом в оптимальных почвенных условиях у всех выделенных сортов полулеталь-ные формы отсутствовали.
В 2005 г. с целью подтверждения наличия скрытой полулетальности вновь выделенные сортообразцы повторно высеяли на участке с уплотнённой почвой. Дополнительно в эксперимент был введен второй тест-контроль стрессовости почвенных условий – сорт Т-245 с идентифицированной хлорофилльной мутацией.
Полученные данные подтвердили наличие скрытой полуле-
Сумма осадков -е- Урожайность
Рисунок 1 – Сумма осадков за летний (июнь-август) период и средняя продуктивность сои в конкурсном сортоиспытании ВНИИМК
Таблица 1 – Частота проявления полулетальных растений сои в условиях эдафического стресса, %
Краснодар, ВНИИМК, 2004 г.
|
Сорт |
Оптимальные условия |
Эдафический стресс |
|
Фора (контроль) |
0,96 |
28,57 |
|
Приморская 56 |
0,00 |
7,14 |
|
Приморская 93 |
0,00 |
20,00 |
|
Е-Шен-Доу |
0,00 |
19,50 |
|
к-5683 ( G. gracilis ) |
0,00 |
22,20 |
тальной мутации у выделенных 4 сортообразцов.
Частота проявления хлорозной формы у сорта Т-245 возросла почти в 300 раз – с 0,02 до 5,88%. Это также подтверждает эффективность использования эдафиче-ского стресса для выделения генотипов сои со скрытой полулетальностью других типов.
В результате проведенного эксперимента дополнительно было выделено ещё 16 сортообраз-цов, часть растений у которых имела признаки пониженной стерильности различных форм (табл. 2).
Как следует из табл. 2, частота проявления растений с пониженной фертильностью в оптималь- ных условиях у тест-контролей – сортов Фора и Т-245 – составила 0,96 и 0,02 % соответственно. У остальных сортообразцов полу-стерильных растений не встречалось.
В условиях эдафического стресса проявление признака пониженной фертильности у сорта Фора увеличилось до 26,58 %, а у сорта Т-245 – до 5,88 %. Частота проявления признака пониженной фертильности у других выделенных сортов в условиях эдафиче-ского стресса варьировала от 2,50 у линии ДХМ до 22,72 % у сорта OS-87-4.
В основном у исследуемых генотипов обнаруживали субвитальную форму пониженной фер- тильности. У двух сортообразцов: Т-215 и Дива, выделены слабофертильные растения мелколистной формы с частотами 17,64 и 5,26 %. Хлорозная и карликовая формы не были обнаружена ни у одного генотипа.
В целом, ежегодное образование субвитальных, а также хлорозных и мелколистных форм сои с пониженной фертильностью свидетельствует о наличии определенного генетического фактора, экспрессирующего этот признак. Мы предполагаем присутствие субвитальной или полулетальной мутации в генотипе таких сорто-образцов. Невысокая частота фенотипического проявления мутаций может определяться наличием отдельных или комплексов генов-компенсаторов, нейтрализующих негативный эффект мутации. Увеличение частоты образования полулетальных растений в стрессовых условиях, в частности при воздействии эдафического стрессора, может указывать на уменьшение эффективности ККГ.
Ранее нами при исследовании митоза сорта Фора было установлено, что полулеталь, даже будучи скомпенсированной, может оказывать дестабилизирующее действие на прохождение хромосомой, в которой она локализована, отдельных стадий митотического цикла. Визуально чаще всего это выражается в отставании отдельных или групп хромосом на стадии метафазы или анафазы (Зеленцов, Мошненко, 2003).
С целью подтверждения нашего предположения о наличии скомпенсированных скрытых по-лулетальных мутаций исследовали стабильность митоза у 20 выделенных сортообразцов с частичным проявлением полулетальности (табл. 3).
Как следует из представленных в табл. 3 данных, частота митотических нарушений у контрольного сорта Лань составила всего 0,5 %. Количество метафазных нарушений у сорта Фора достигало 3,2 %. В аналогичных почвенно-климатических условиях половина из исследуемых сортообразцов отличались близкой к уровню сорта
Таблица 2 – Частота проявления разных форм полулетальных растений в условиях эдафического стресса, %
ВНИИМК, 2005 г.
|
Сорт |
Оптимальные почвенные условия |
Эдафический стресс |
Морфологический тип полулетальности |
|
Т-245 (контроль 1) |
0,02 |
5,88 |
Субвитальная |
|
Фора (контроль 2) |
0,96 |
26,58 |
Хлорозная |
|
ОS-87-4 |
0,00 |
22,72 |
Субвитальная |
|
Л-87-2418 |
0,00 |
22,22 |
«» |
|
к-5683 ( G. gracilis ) |
0,00 |
22,20 |
«» |
|
Stine 2252 |
0,00 |
20,58 |
«» |
|
Приморская 93 |
0,00 |
20,00 |
«» |
|
Холодостойкая линия 4 |
0,00 |
20,00 |
«» |
|
Stine 1570 |
0,00 |
20,00 |
«» |
|
Е-Шен-Доу |
0,00 |
19,50 |
«» |
|
Д-2861/3/05 |
0,00 |
18,18 |
«» |
|
К- 002 |
0,00 |
16,60 |
«» |
|
Д-8520 |
0,00 |
14,28 |
«» |
|
Stine 1386 |
0,00 |
12,50 |
«» |
|
Л-828 |
0,00 |
9,09 |
«» |
|
Волна |
0,00 |
7,89 |
«» |
|
Приморская 56 |
0,00 |
7,14 |
«» |
|
Куйбышевская 77 |
0,00 |
6,45 |
«» |
|
L-63 |
0,00 |
4,16 |
«» |
|
ДХМ |
0,00 |
2,50 |
«» |
|
Т-215 |
0,00 |
17,64 |
Мелколистная |
|
Дива |
0,00 |
5,26 |
«» |
Таблица 3 – Частота нарушений митоза у сортов сои с признаками полулетальности
Краснодар, ВНИИМК, 2005 г.
Фора или даже более высокой долей клеток с нарушениями мито-
за (см. табл. 3). Из них максимальное количество отставаний
хромосом было зафиксировано у сорта Stine 2252 – 6,4 % и линии ДХМ – 5,5 %. При этом у этих сортообразцов также отмечено максимальное в опыте количество аномальных анафаз – 1,2 и 1,1 % соответственно.
Визуальный анализ аномальных метафаз и анафаз позволил установить, что линейные размеры отстающих хромосом и их пространственная ориентация в объеме клетки были специфичны практически для каждого сорта. Однако очень маленькие размеры хромосом и ограниченные возможности светового микроскопа не позволили провести их более детальную идентификацию. В связи с этим удалось выделить только три морфологических типа (рис. 5 и рис. 6).
На рис. 6 представлено 3 типа регулярно отстающих, предположительно мутантных хромосом.
Тип I представлен ранее уже изученной нами спутничной субметацентрической парой хромосом с наличием вторичной перетяжки, характерной для сорта Фора и ее потомков.
Тип II, выявленный у сорта Stine 2252, представлен субметацентрической парой хромосом, предположительно локализуемой в пределах 5-7-ой пары хромосом на кариограмме.
Тип III выявлен у сорта Л-8520 и представляет собой пару мелких акроцентрических хромосом, предположительно локализованных в пределах 11-13-ой пары хромосом кариограммы.
Выводы. Выращивание сои в условиях эдафического стресса приводит к увеличению частоты образования полулетальных растений за счет предполагаемого снижения эффективности нейтрализации мутации ККГ-комплексом.
При оценке 700 коллекционных образцов сои различного происхождения в условиях эда-фического стресса выделено 20 сортообразцов с увеличенной частотой образования полулеталь-ных растений.
При цитогенетическом иссле-
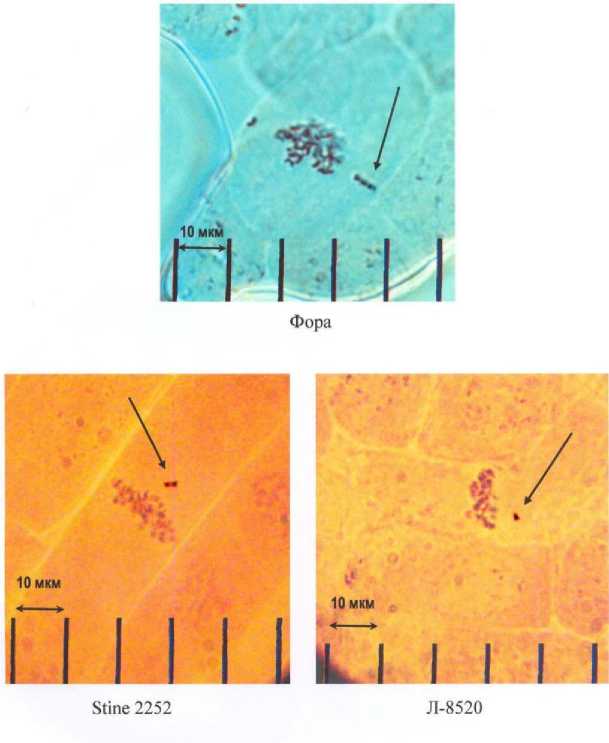
Рисунок 5 – Отставание мутантных хромосом у генотипов сои со скрытой полулетальностью
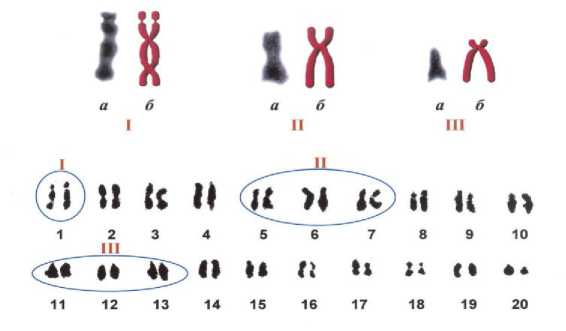
Рисунок 6 – Типы выявленных отстающих мутантных хромосом и их примерная локализация в кариотипе сои а – видимая морфология хромосомы;
б – реконструкция хромосомы.
довании полулетальных форм растений установлена повышенная (до 7,9 %) частота нарушений
митоза в виде отставаний отдельных хромосом в анафазе, позволяющая предположить наличие в
геноме полулетальной мутации.
