Взаимодействие синтетических производных витамина е на реологические свойства эритроцитарных мембран

Автор: Панин Лев Евгеньевич, Мокрушников Павел Валентинович
Журнал: Science for Education Today @sciforedu
Рубрика: Биологические, химические, медицинские науки
Статья в выпуске: 5 (15), 2013 года.
Бесплатный доступ
Методами флуоресцентного анализа исследовалось влияние синтетических производных витамина Е тиофана и тиофана-М на реологические свойства мембран эритроцитов. Первый препарат снижал микровязкость как в области липид-липидных, так и белок-липидных взаимодействий. Второй препарат снижал микровязкость в области белок-липидных взаимодействий и повышал в области липид-липидных взаимодействий. Проверка эффективности антиоксидантной защиты эритроцитарных мембран показала, что тиофан значительно подавляет перекисное окисление липидов мембран и образование белково-липидных сшивок, восстанавливая реологические свойства эритроцитов. Данный препарат рекомендован нами для профилактики перекисного окисления липидов у лиц, работающих в условиях Арктики.
Микровязкость мембран, перекисное окисление липидов, тиофан
Короткий адрес: https://sciup.org/147137489
IDR: 147137489 | УДК: 538.9
Effects of synthetic vitamin e derivative on the rheological properties of red blood cell membranes
Methods of the fluorescent analysis was investigated action of derivatives of vitamin E tiofan and tiofan-M on rheological properties of membranes of erythrocytes. The first formulation reduced the microviscosity of a lipid-lipid, and protein-lipid interactions. The second drug reduced microviscosity of the protein-lipid interactions, and increased in the area of a lipid-lipid interactions. Checking the effectiveness of antioxidant erythrocyte membranes showed that thiophane significantly inhibits peroxidation of membrane lipids and the formation of lipid-protein cross-links, restoring the rheological properties of red blood cells. This drug is recommended by us to prevent lipid peroxidation in persons working in Arctic conditions.
Текст научной статьи Взаимодействие синтетических производных витамина е на реологические свойства эритроцитарных мембран
Электронный журнал «Вестник Новосибирского государственного педагогического университета» 5(15) 2013 ISSN 2226-3365
Все права защищены

5(15) 2013
ISSN 2226-3365
(геомагнитные бури), при действии на организм ионизирующей радиации. В высоких широтах это связано с активным проникновением в магнитосферу частиц «солнечного ветра» [3].
Естественной защитой биологических мембран от перекисного окисления липидов служит витамин Е (токоферол). Основным источником витамина Е и других антиоксидантов (витамины А, С) являются продукты питания. Нарушение режима питания, использование в пищу рафинированных продуктов создают дефицит природных антооксидантов в организме, что способствует усилению перекисного окисления липидов биологических мембран и нарушению их функций [4]. Нежелательным последствием этих изменений в эритроцитарных мембранах является увеличение их микровязкости и нарушение их реологических свойств. Это затрудняет продвижение эритроцитов по капиллярам и приводит к диффузной гипоксии тканей. На крайнем Севере это явление известно как
«полярная одышка» [5]. Перед современной медициной стоит важная задача обеспечить человека, работающего в условиях Арктики, дополнительным количеством антиоксидантов. Такие препараты были получены на кафедре химии Новосибирского государственного педагогического университета [6].
В данной работе была дана оценка эффективности антиоксидантных свойств производных витамина Е тиофан-М и тиофан с использованием в качестве модели биологических мембран плазматических мембран эритроцитов.
МЕТОДЫ И МАТЕРИАЛЫ
Материалы. В работе использовались производные витамина Е тиофан-М додецил-[3-(3´,5´-диметил-4-гидроксифенил)метил] сульфид и тиофан бис-[3-(3´,5´-ди-трет-бутил-4-гидроксифенил)пропил]сульфид производства ФГБОУ ВПО «НГПУ», НИИ химии антиоксидантов.
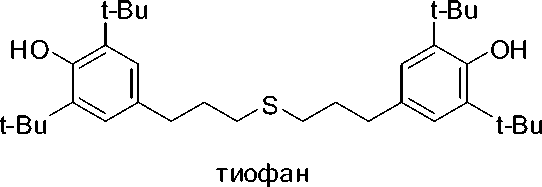
SC 12 H 25
т иофа н-М
Флуоресцентный анализ теней эритроцитов. Эритроциты получали из свежевыделенной крови после декапитации самцов крыс линии Вистар под легким нембуталовым наркозом. Кровь разбавляли впятеро изотоническим фосфатным буфером pH 7.35, содержащим 0.043 M КН2РО4 и 0.136 M Na2НРО4. После осаждения клеток с помощью центрифугирования при 330 g в течение 10 мин надосадочную жидкость сливали и процедуру повторяли еще 2 раза. Тени эритроцитов были получены после их гемолиза в гипотоническом фосфатном буфере (рН 7.35), содержащем 2.75 мM KH2РО4 и 8.5 мM Na2НРО4. Тени осаждались центрифугированием при 5500 g, надосадочная жидкость сливалась. Процедура
Все права защищены

5(15) 2013
ISSN 2226-3365
повторялась четыре раза. Получение и хранение теней производилось при 4 ° C.
Измерения флуоресценции проводились на спектрофлуориметре «Шимадзу» RF-5301(PC)SCE. В кварцевую кювету размером 1х1х4 см3 наливали 4 мл гипотонического фосфатного буфера содержащего 2,75 mМ КН 2 РО 4 и 8,5 mМ Na 2 HPО 4, рН=7,35 и тени эритроцитов. Концентрация белков теней определялась методом Варбурга и Кристиана по изменению оптической плотности взвеси [7]. В среднем она колебалась в пределах 0,200-0,350 мг/мл.
Кювета с взвесью теней помещалась в термостат спектрофлуориметра на 10 минут. Выход температуры в кювете на стационарный режим контролировался электронным термометром. Во всех экспериментах температура в кювете была 360С. После выхода температуры в кювете на стационарный режим проводились контрольные измерения интенсивности собственной флуоресценции остатков триптофана в белках мембран. Снимался спектр излучения триптофана в диапазоне 300нм < X < 400нм при длине волны возбуждения X = 281нм, при этом максимум интенсивности излучения приходился на X = 332нм. Спектральная ширина щелей 1,5/10.
Измерение микровязкости мембран эритроцитов. Измерения микровязкости мембран эритроцитов проводились на сперктрофлуориметре «Шимадзу» RF-5301(PC)SCE. Опытный образец готовился следующим образом. В кварцевую кювету размером 1х1х4 см3 наливалось 4 мл гипотонического фосфатного буфера (рН 7.35), содержащем 2.75 мM KH 2 РО 4 и 8.5 мM Na 2 НРО 4 , тени эритроцитов, необходимое количество производных витамина Е, флуоресцентный зонд пирен. Все компоненты до этого хранились при 40С.
Концентрация белка теней в кювете составляла 0.200–0.350 мг/мл, пирена – 7.76х10-6 М. Пирен разводился в этаноле, его исходная концентрация составляла – 1.5х10-3 М. Кювету помещали в термостат спектрофлуориметра на 10 минут, после этого проводили измерения флуоресценции при температуре 360С. Перед тем как поставить пробу в термостат спектрофлуориметра ее энергично встряхивали в течение 1 минуты. Для измерения флуоресценции теней при нагружении их другим количеством производных витамина Е каждый раз точно также готовилась новая проба. Такая процедура связана с тем, что пирен способствует быстрой деградации мембран эритроцитов.
Для измерения микровязкости липидного бислоя вблизи мембранных белков (область белок-липидного взаимодействия) использовалась длина волны возбуждения X = 281 нм и спектральная ширина щелей 1.5/5. Микровязкость липидного бислоя вдали от мембранных белков (область липидлипидного взаимодействия) использовалась длина волны возбуждения X = 337 нм и спектральная ширина щелей 1.5/3. Максимумы излучения наблюдались при X = 374 нм и X = 393 нм (вибронные пики излучения мономеров пирена), и X = 468 нм (максимум излучения димеров пирена).
Относительная микровязкость мембран т n ( A ) описывалась отношением L = , где n ( A )
П(0)
и η(0) микровязкости мембран при добавлении во взвесь тиофана концентрации А и без добавления тиофана, соответственно. Для области липид-липидного взаимодействия относительная микровязкость L вычислялась по формуле
Все права защищены

5(15) 2013
ISSN 2226-3365
L _ n ( A ) _ ^ 468 (0) F 393 ( A ) n ( 0 ) F 16 . ( a ) F 393 (0)
где F 468 ( A ) интенсивность флуоресценции пирена при X = 468 нм при концентрации А гормона во взвеси; F 468(0) интенсивность флуоресценции пирена при X = 468 нм при отсутствии гормона во взвеси. F 393( A ) and F 393 (0) интенсивность флуоресценции пирена при при X = 393 нм при концентрации А гормона во взвеси и при отсутствии гормона во взвеси, соответственно.
Для области белок-липидного взаимодействия относительная микровязкость L вычислялась по формуле
L _ п ( A ) _ F 468 (0) - I 468 _ F 393 ( A ) - I 393 n ( 0 ) F 468 ( A ) - I 468 F 393 (0) - I 393
где I 393 и I 468 интенсивность флуоресценции триптофановых остатков в мембранных белках при X = 393 нм и X = 468 нм, соответственно. Относительная погрешность измерения относительной микровязкости равна 6 %.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Была получена зависимость относительной микровязкости мембран эритроцитов L от удельной концентрации тиофана-М во взвеси (рис. 1). В работе рассчитывалась удельная концентрация тиофана-М и тиофана: молярная концентрация этих молекул поделена на концентрацию белков теней во взвеси. Относительная микровязкость мембран в области липид-липидного взаимодействия при увеличении концентрации тиофана-М вначале увеличивалась. После увеличения на 20 % она вышла на плато. В области белок- липидного взаимодействия напротив, микровязкость уменьшалась на 12 % и выходила на плато. Интенсивность собственной флуоресценции остатков триптофана в белках мембран при добавлении тиофана-М увеличивалась (рис. 2), при 10-9 моль/мг белка выходила на плато. При добавлении тиофана микровязкость уменьшалась как в области липид-липидных взаимодействий, так и в области белок-липидных взаимодействий мембран (рис. 3). Микровязкость достигала минимума при 0.7·10-9 моль/мг белка, при дальнейшем увеличении концентрации тиофана микровязкость увеличивалась. Максимальное уменьшение микровязкости составляло 20 % для области липид-липидного взаимодействия и 15 % для области белок-липидного взаимодействия. Интенсивность собственной флуоресценции остатков триптофана в белках мембран при добавлении тиофана также незначительно увеличивалась (рис. 4).
Можно предположить следующий механизм взаимодействия тиофана и тиофана-М с мембраной. Тиофан и тиофан-М, подобно холестерину, погружались в фосфолипидный бислой, заякоривались своими фенольными кольцами с гидрофильными ОН-группами на внешней поверхности мембран, где много гидрофильных групп фосфолипидов и белков мембраны. В гидрофобную область мембран проникали жирнокислотные хвосты, содержащие сульфидную группу. Можно предположить, что эти хвосты так перераспределяли диполи воды, содержавшиеся в биомембране, что микровязкость мембран уменьшалась. С другой стороны, подобно холестерину, тиофан и тиофан-М, по-видимому, могли увеличивать жесткость мембраны за счет
Все права защищены

5(15) 2013
ISSN 2226-3365
образования новых связей между своими активными группами и полярными группами фосфолипидов и белков мембраны. В случае тиофана-М механизм увеличения жесткости мембраны преобладал над его уменьшением, микровязкость в липид-липидной области увеличивалась. Диполи воды вытеснялись в область белок-липидных взаимодействий, где микровязкость уменьшалась, а интенсивность собственной флуоресценции остатков триптофана в белках мембран увеличивалось (рис. 3). Проникновение тиофана способствовало уменьшению микровязкости мембраны, но параллельно работал механизм увеличения микровязкости, что приводило к куполообразной форме зависимости микровязкости от концентрации тиофана.
Разницу во влиянии на микровязкость мембран тиофана и тиофана-М можно объяснить тем, что в тиофане атом серы находится дальше от бензольного кольца. Он проникал глубже в гидрофобную часть мембраны, мог более равномерно перераспределять диполи воды между жирнокислотными хвостами фосфолипидов. Молекулы воды, встраиваясь между жирнокислотными хвостами фосфолипидов, увеличивали расстояние между молекулами фосфолипидов и ослабляли взаимодействие между ними. Это приводило к уменьшению жёсткости мембраны.
Исследовалось защитное действие тиофана от перекисного окисления липидов (ПОЛ) (рис. 5 и 6). Для моделирования ПОЛ во взвесь теней добавляли 3мМ перекиси водорода Н 2 О 2 . В результате ПОЛ происходит образование радикалов в области связей С=С в жирнокислотных хвостах фосфолипидов. Это приводило к образованию сшивок между молекулами, усилению жесткости биомембран. При добавлении 3мМ перекиси водорода к взвеси
Тиофан является высокоэффективным ингибитором процессов свободнорадикального окисления, что обусловлено синергетическим сочетанием антирадикальной активности его фенольных групп с противопероксидной активностью бивалентной серы [6]. Тиофан уменьшает количество сшивок между молекулами фосфолипидов, тем самым уменьшая микровязкость биомембраны. Это позволяет эритроцитам более легко проходить по капиллярам, уменьшается артериальное давление, уменьшается возможность развития ишемической болезни сердца.
ЗАКЛЮЧЕНИЕ
Экспериментально установлено, что тиофан и тиофан-М при добавлении во взвесь эритроцитов взаимодействуют с мембранами и уменьшают их микровязкость. Тиофан активно защищает мембрану эритроцитов от перекисного окисления липидов, уменьшает микровязкость мембран и восстанавливает реологические свойства эритроцитов.
Все права защищены

5(15) 2013
ISSN 2226-3365
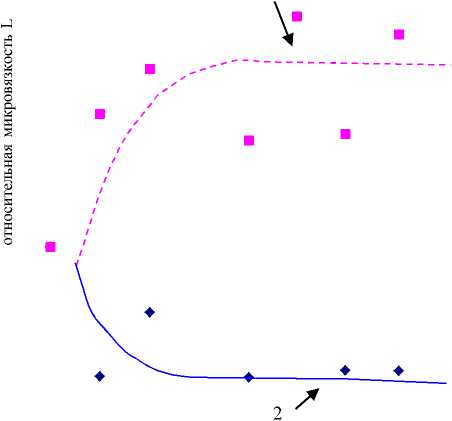
удельная концентрация тиофана-М (10-9 моль/мг белка)
Рисунок 1.
Зависимость относительной микровязкости мембран эритроцитов L от удельной концентрации тиофана-М во взвеси. Кривая 1 – область липид-липидного взаимодействия, кривая 2 – область белок – липидного взаимодействия.
Рисунок 2.
Зависимость тушения флуоресценции триптофана от удельной концентрации тиофана-М во взвеси.
F – интенсивность флуоресценции триптофановых групп мембранных белков при добавлении тиофана-М, F(0) – интенсивность флуоресценции триптофановых групп мембранных белков при нулевой концентрации тиофана-М.

удельная концентрация тиофана-М (10-9 моль/мг белка)
Все права защищены

5(15) 2013
ISSN 2226-3365
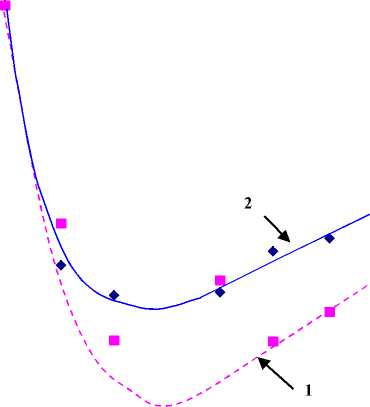
Рисунок 3.
Зависимость относительной микровязкости мембран эритроцитов L от удельной концентрации тиофана во взвеси.
Кривая 1 – область липид-липидного взаимодействия, кривая 2 – область белок – липидного взаимодействия.
удельная концентрация тиофана (10-9 моль/мг белка)
Рисунок 4.
Зависимость тушения флуоресценции триптофана от удельной концентрации тиофана во взвеси.
F – интенсивность флуоресценции триптофановых групп мембранных белков при добавлении тиофана-М, F(0) – интенсивность флуоресценции триптофановых групп мембранных белков при нулевой концентрации тиофана.

удельная концентрация тиофана (10-9 моль/мг белка)
Все права защищены
5(15) 2013
ISSN 2226-3365
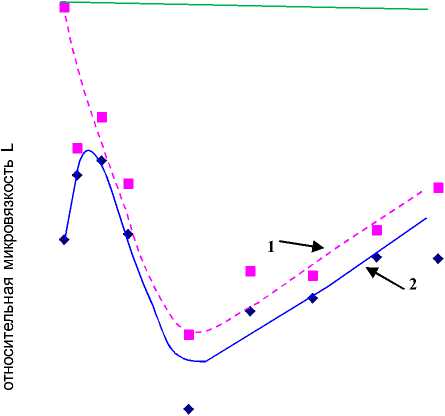
Рисунок 5.
Зависимость относительной микровязкости мембран эритроцитов L от удельной концентрации тиофана во взвеси при добавлении во взвесь 3 мМ перекиси водорода.
За единицу принята микровязкость мембран без добавления перекиси водорода и тиофана.
Кривая 1 – область липид-липидного взаимодействия, кривая 2 – область белок – липидного взаимодействия.
Горизонтальная прямая – значение относительной микровязкости в области липид-липидного взаимодействия при добавлении 3 мМ перекиси водорода без добавления тиофана.
удельная концентрация тиофана (10-9 моль/мг белка)
Рисунок 6.
Зависимость тушения флуоресценции триптофана от удельной концентрации тиофана во взвеси при добавлении во взвесь 3 мМ перекиси водорода. F – интенсивность флуоресценции триптофановых групп мембранных белков при добавлении тиофана, F(0) – интенсивность флуоресценции триптофановых групп мембранных белков при нулевой концентрации тиофана.
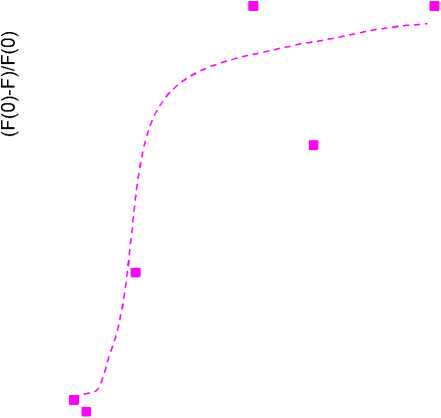
удельная концентрация тиофана (10-9 моль/мг белка)
Все права защищены

5(15) 2013
ISSN 2226-3365
Список литературы Взаимодействие синтетических производных витамина е на реологические свойства эритроцитарных мембран
- Панин Л. Е. Детерминантные системы в физике, химии, биологии. -Новосибирск: Сиб. унив. изд-во, 2006. -202 с.
- Панин Л. Е., Мокрушников П. В., Куницын В. Г., Панин В. Е., Зайцев Б. Н. Основы многоуровневой мезомеханики наноструктурных переходов в мембранах эритроцитов и их разрушения при взаимодействии с гормонами стресса//Физическая мезомеханика. -2011. -т. 14. -№ 1. -С. 5-17.
- Казначеев В. П., Куликов В. Ю., Панин Л. Е. Особенности экологических факторов высоких широт/Механизмы адаптации человека в условиях высоких широт/Отв. ред. Казначеев В. П. -Л.: Медицина, 1980. -С. 10-24.
- Панин Л. Е. Человек в экстремальных условиях Арктики//Бюлл. СО РАМН. -2010. -т. 30. -№3. -С. 92-97.
- Авцын А. П., Марачёв А. Г. Проявления адаптации и дезадаптации у жителей Крайнего Севера//Физиология человека. -1975. -т.4. -С. 3-14.
- Овчинникова Л. П., Роцкая У. Н., Васюнина Е. А., Синицина О. И., Кандалинцева Н. В., Просенко Н. В., Невинский Г. А. Антиокислительная активность тиофана [бис-[3-(3´,5´-ди-трет-бутил-4-гидроксифенил)пропил]сульфидa]//Биорг. Химия. -2009. -Т. 35. -№ 3. -С. 417-423.
