Взаимосвязь метаболизма жирных кислот и перекисного окисления липидов у двух видов байкальской эндемичной водоросли Draparnaldioides

Автор: Васильева Ульяна Андреевна, Кириченко Кузьма Анатольевич, Тимофеев Максим Анатольевич, Грабельных Ольга Ивановна
Журнал: Вестник Бурятского государственного университета. Биология, география @vestnik-bsu-biology-geography
Рубрика: Биология
Статья в выпуске: 1, 2023 года.
Бесплатный доступ
Макроводоросли являются одним из важных звеньев водных экосистем, служат источником органического вещества, местообитанием и источником биологически активных веществ для многих бентосных организмов. Изучение метаболических процессов в клетках эндемичных водорослей оз. Байкал представляет особый интерес. В работе оценивали жирнокислотный состав (ЖК-состав), содержание пероксида водорода и малонового диальдегида (МДА) у двух видов эндемичного рода Draparnaldioides (D. arnoldi и D. baicalensis) сразу после вылова и при культивировании в лабораторных условиях. При культивировании выявлены разнонаправленные изменения в содержании ЖК и МДА, но не пероксида водорода. Обсуждается связь метаболизма ЖК и перекисного окисления липидов у водных организмов.
Перекисное окисление липидов, жирные кислоты, эндемик, озеро байкал, водоросли, малоновый диальдегид
Короткий адрес: https://sciup.org/148326438
IDR: 148326438 | УДК: 57(577.2) | DOI: 10.18101/2587-7143-2023-1-3-11
Possible relationship between fatty acids and lipoperoxidation in two species of the Baikal endemic algae Draparnaldioides
Macroalgae are one of the important parts of aquatic ecosystems. They are a habitat and a source of organic matter and biologically active substances for many benthic organisms. The Study of metabolic processes in the cells of endemic algae of Lake Baikal is of particular interest. We have evaluated fatty acid composition (FA), hydrogen peroxide and malondialdehyde (MDA) content in two species of endemic genus Draparnaldioides (I) , arnoldi and D. baicalensis) immediately after harvesting and during cultivation in laboratory. During cultivation we have revealed multidirectional changes in the content of fatty acids and MDA, as distinguished from the content of hydrogen peroxide. We also discussed the relationship between FA metabolism and lipoperoxidation in aquatic organisms.
Текст научной статьи Взаимосвязь метаболизма жирных кислот и перекисного окисления липидов у двух видов байкальской эндемичной водоросли Draparnaldioides
Введение. Эндемичные макроводоросли рода Draparnaldioides sp. выполняют немалую роль в растительной продукции оз. Байкал и являются основной водорослью третьего растительного пояса и захватывают глубины от 2,5 до 12–25 м [2; 4]. Род Draparnaldioides относится к отделу Chlorophita , класс Ulotrichophyceae , порядок Ulotrichales, подпорядок Chaetophorineae , семейство Chaetophoraceae , подсемейство Chaetophorideae , род Draparnaldioides (Meyer et Skabitsch ex Vishnyakov, 2020). С сентября 2020 г. род Draparnaldioides насчитывает 10 видов: D. arenaria, D. arnoldii, D. baikalensis, D. goroschankinii, D. lubrica, D. pumila, D. pilosa, D. plumose, D. simplex и D. vilosa [9]. Описание водоросли наиболее полно отображено у Л. А. Ижболдиной в книге «Атлас и определитель водорослей бентоса и перифитона озера Байкал (мейо- и макрофиты) с краткими очерками по их экологии» (2007). Талломы водоросли кустиковидные, высотой до 10–40 см, имеют главный ствол, от которого отходят крупные ветви 2-го и 3-го порядка, несущие боковые веточки, располагающиеся мутовками, реже одиночные или супротивные [4]. Водоросль имеет мощный ризоидный покров и слизистый чехол, защищающие растение от сильного волнового воздействия. Также у водоросли наблюдается сетчато-продырявленный хлоропласт, занимающий всю высоту клетки [4]. Активная вегетация водоросли приходится на весенне-осенний временной период (по данным Л. А. Ижболдиной — с июня по октябрь).
Все растительные и животные организмы оз. Байкал обитают в условиях невысоких температур, малого количества органических веществ, низкой минерализации и высокого содержания кислорода (до 14 мг/л). Макроводоросль ведет прикрепленный образ жизни, вследствие чего не может уйти от воздействия неблагоприятных факторов окружающей среды. Высокое содержание кислорода и прозрачность воды оз. Байкал в той или иной мере должны отражаться на процессах защиты водоросли от окислительного стресса. Индикатором окислительного стресса в талломах водорослей может служить оценка уровня перекисного окисления липидов (ПОЛ) в клеточных мембранах [5]. У прикрепленной формы морской красной водоросли Ahnfeltiopsis flabelliformis показана активация ПОЛ в периоды высокой освещенности (апрель, июль — сентябрь) [5]. В этот же период авторы наблюдали снижение концентрации фотосинтетических пигментов в результате фотоповреждения. В литературе отсутствуют какие-либо сведения по физиологии и биохимии макроводоросли рода Draparnaldioides sp. и ее устойчивости к неблагоприятным факторам среды. Ранее нами были обнаружены отличия между отдельными видами Draparnaldioides sp. (D. arnoldi и D. baicalensis) по содержанию фотосинтетических пигментов и жирных кислот (ЖК) суммарных липидов при культивировании макроводоросли в лабораторных условиях [1]. Разнонаправленные изменения ЖК-состава могут свидетельствовать о различиях в содержании активных форм кислорода (АФК) и/или интенсивности протекания ПОЛ [6].
Целью работы являлось изучение физиолого-биохимических параметров у макроводоросли в момент вылова и в процессе культивирования в лабораторных условиях.
В задачи данного исследования входило: 1) оценить содержание пероксида водорода и продуктов ПОЛ в талломах D. arnoldii и D. baicalensis сразу после вылова и в процессе культивирования в лабораторных условиях; 2) провести сравнительный анализ содержания продуктов ПОЛ и Н 2 О 2 с изменениями ЖК-состава у изученных видов.
Материалы и методы. Объектом исследования являются два вида — D. ar-noldii и D. baicalensis . Виды различаются между собой по форме боковых веточек и наличию/отсутствию у них рахиса. У D. arnoldii рахис в боковых веточках выражен, боковые веточки в очертаниях округлые, веерообразно разветвлены [4]. У D. baicalensis рахис в боковых веточках короткий, состоящий обычно из 1–2 клеток, от которых сразу начинаются ветви более-менее одинакового размера, обильно ветвящиеся в верхней части и имеющие округло-треугольную форму [4]. Если же сравнивать визуально, то D. arnoldii является более тонкой и изящной, не имеет массивного слизистого чехла, в то время как D. baicalensis — достаточно массивная и у нее присутствует заметный слизистый чехол, наблюдаемый при окраске препарата тушью (рис. 1).
Для работы пробы макроводоросли отбирали в конце августа — начале сентября на оз. Байкал в районе пос. Большое Голоустное по координатам от 52°01'33.6"N 105°25'06.7"E до 52°01'33.5"N 105°25'59.3"E. Температура воды в момент вылова составляла 11 ° С. Водоросль брали с уреза воды на глубинах до 0,5 м. Идентификацию проводили согласно определителю Л. А. Ижболдиной [4]. Часть талломов фиксировали сразу на месте в жидком азоте. Основную биомассу водоросли перевозили в сумках-холодильниках для постановки опыта по культивированию в лабораторных условиях. Культивировали водоросль в байкальской воде в течение 14 суток при непрерывном освещении с интенсивностью 2 500 лк в инкубаторе MIR-153 (Sanyo, Япония). На 7-е и 14-е сутки фиксировали материал (талломы) в жидком азоте для сравнения с точкой отбора.
Содержание H 2 O 2 определяли с ксиленоловым оранжевым [7]. Для этого навеску таллома водоросли (0,25 г) замораживали в жидком азоте и затем растирали в 2,5 мл 0,1%-ной трихлоруксусной кислоты (ТХУ) на льду. Гомогенат центрифугировали 15 мин на 12 000 g при 4 оС. Для определения содержания пероксида водорода супернатант разводили 0,1%-ной ТХУ (1:4) и готовили смесь

а б
Рис. 1. Внешний вид исследуемых макроводорослей. а — D. arnoldi ; б — D. baicalensis. Показан внешний вид без микроскопа (нижнее фото в чашках Петри), микроскопия талломов (среднее фото) и окрашенные тушью препараты (верхнее фото) Бар равен 500 μm.
супернатанта и реагента в соотношении 1:1 (по 0,5 мл каждого). Состав реагента: 0,5 мМ FeSO 4 × (NH 4 ) 2 SO 4 × 6H 2 O, 0,5%-ной (v/v) H 2 SO 4 , 200 мкМ ксиленолового
оранжевого (AppliChem, Германия) и 200 мМ сорбитола (Gerbu, Германия). Смесь инкубировали в течение 25 мин на мини-термошейкере TS-100 (BioSan, Латвия) при температуре 26 °С, затем пробы центрифугировали при 12 000 g в течение 5 мин. Определяли поглощение конечного продукта спектрофотометрически (SmartSpec Plus, Bio-Rad, США) при 560 нм. Содержание Н 2 О 2 рассчитывали по калибровочной кривой и выражали в мкМ/г сырого веса [3].
Содержание продуктов ПОЛ оценивали по накоплению в тканях продуктов, реагирующих с тиобарбитуровой кислотой (ТБК-РП) [8]. Для определения содержания ТБК-РП 0,5 мл супернатанта добавляли к 1 мл 0,5%-ной (w/v) ТБК (Диаэм, Россия) в 20%-ной ТХУ. Смесь инкубировали на кипящей водяной бане 30 мин, останавливали реакцию охлаждением, помещая пробирки в лед. Образцы центрифугировали 5 мин при 12 000 g и измеряли абсорбцию при 532 и 600 нм. Содержание ТБК-РП рассчитывали с использованием коэффициента экстинкции ТБК 155 мМ–1 см–1 после вычитания неспецифического поглощения при 600 нм и выражали в нМ/г сырого веса [3].
Данные представлены как среднее арифметическое с разбросом значений в виде стандартного отклонения в случае нормального распределения данных или как медианное значение с разбросом значений в виде интерквартильной широты (25; 75% процентиль) в случае распределения, отличающегося от нормального. Нормальность распределения проверяли с помощью критерия Шапиро — Уилка. Для доказательства значимых различий в случае нормального раcпределения данных использовали метод множественного сравнения средних Фишера или метод Тьюки, в случае распределения данных, отличающихся от нормального, использовали критерий Крас-келла — Уоллиса. Различия между данными считали статистически значимыми при p <0,05. Количество биологических повторностей было 2–3. Статистические расчеты осуществляли с помощью программного пакета SigmaPlot 12.0.
Результаты и обсуждение. Ранее было показано, что культивирование D. ar-noldii и D. baicalensis в лабораторных условиях приводило к изменению жирнокислотного состава талломов водоросли. Содержание ЖК у двух видов макроводоросли на момент вылова не различалось, но в процессе культивирования происходили разнонаправленные изменения. У D. arnoldii в процессе культивирования количество ненасыщенных ЖК увеличилось, в то время как у D. baicalensis количество ненасыщенных ЖК снизилось (рис. 2).
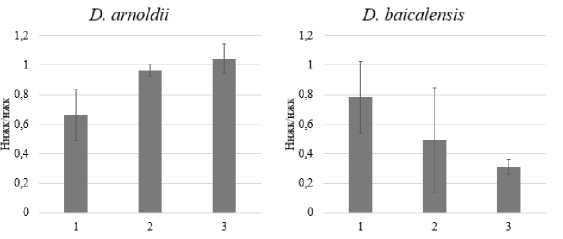
1 — 0 суток; 2 — 7 суток; 3 — 14 суток
Рис. 2. Изменение отношения суммы ненасыщенных ЖК к сумме насыщенных у двух видов Draparnaldioides сразу после вылова (0 суток), через 7 и 14 суток культивирования в лабораторных условиях . Представлено среднее значение и его стандартное отклонение
Снижение содержания ненасыщенных ЖК может свидетельствовать об интенсификации процессов ПОЛ. Одним из продуктов ПОЛ является малоновый диальдегид (МДА), который рассматривается в качестве маркера окислительного стресса. МДА наряду с другими «тиобарбитуро-реактивными продуктами» вступает в реакцию с тиобарбитуровой кислотой с образованием состава, который можно определить спектрофотометрически.
На рисунке 3 представлено содержание ТБК-РП в талломах анализируемых видов сразу после вылова и в течение 14 суток культивирования. Содержание МДА у двух видов макроводоросли изначально отличается и разнонаправленно изменяется в процессе культивирования (рис. 3). Содержание ТБК-РП было выше у D. baicalensis по сравнению с D. arnoldii на 39%. При этом у D. arnoldii наблюдали небольшой рост данных продуктов на 14-е сутки культивирования, однако различия в содержании были статистически незначимы (Р=0,069). У D. baicalensis происходило значимое снижение данного показателя в процессе культивирования (P = <0,001), на 7-е сутки снижение составило 30%, а на 14-е — 74% относительно точки вылова (0 суток).
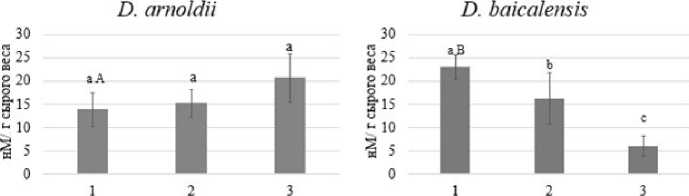
1 — 0 суток; 2 — 7 суток; 3 — 14 суток
Рис. 3. Изменение количества ТБК-РП у двух видов Draparnaldioides сразу после вылова (0 суток), через 7 и 14 суток культивирования в лабораторных условиях. Одинаковыми строчными буквами обозначены статистически не различающиеся варианты внутри вида. Прописными буквами обозначены различия между видами. Представлено среднее значение и его стандартное отклонение
Полученные результаты свидетельствуют, что у обоих видов макроводоросли отсутствует активация процессов ПОЛ, а изменения в содержании ТБК-РП, по-видимому, являются следствием изменений в содержании ненасыщенных ЖК-субстратов ПОЛ.
Процесс ПОЛ запускается активными формами кислорода, поэтому проанализировали содержание одного из видов АФК — пероксида водорода в талломах макроводоросли (рис. 4). Значимых изменений в содержании Н 2 О 2 как сразу после вылова, так и в процессе культивирования отдельных видов, не обнаружили, хотя у D. baicalensis видна тенденция к снижению данного параметра на 14-е сутки культивирования.
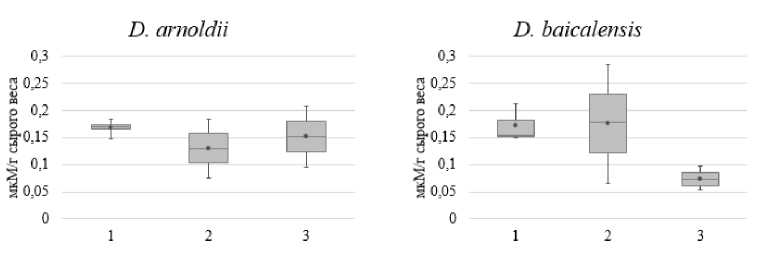
1 — 0 суток; 2 — 7 суток; 3 — 14 суток
Рис. 4. Изменение содержания пероксида водорода у двух видов Draparnaldioides сразу после вылова (0 суток), через 7 и 14 суток культивирования в лабораторных условиях
Макроводоросли ведут прикрепленный образ жизни и как и высшие растения они не могут изменить место своего произрастания для того, чтобы покинуть неблагоприятные условия окружающей среды. Интенсивность освещения и температура среды выступают основными абиотическими факторами, определяющими географическое распространение морских макроводорослей, их выживание и продуктивность [5]. Перенос из естественной среды обитания в лабораторные условия может также выступать стрессором, и его действие на разные виды может различаться. Кроме того, безусловно на физиолого-биохимические параметры влияет сезон вылова, так как существует различие в температуре вод и их освещенности.
Одними из первых на изменение условий обитания реагируют мембранные системы клетки, от состава и соотношения липидов зависит текучесть мембран, определяющая структуру и функции мембранных белков и жизнедеятельность клетки в целом. ЖК-состав липидов и интенсивность ПОЛ — это взаимосвязанные параметры. D. arnoldii и D. baicalensis отличались между собой по содержанию ТБК-РП и по направленности изменений ТБК-РП и ЖК-состава липидов.
Заключение. Таким образом, два изученных вида Draparnaldioides проявили различную реакцию при культивировании талломов в лабораторных условиях. Эти данные предполагают, что отдельные виды, возможно, имеют и различный адаптационный потенциал. Необходимы дальнейшие исследования состояния мембран клеток и других физиолого-биохимических показателей, отражающих адаптационные способности организмов, у представителей байкальского эндемичного рода Draparnaldioides.
Список литературы Взаимосвязь метаболизма жирных кислот и перекисного окисления липидов у двух видов байкальской эндемичной водоросли Draparnaldioides
- Сравнительный анализ состава жирных кислот двух видов байкальского эндемичного рода Draparnaldioides / У. А. Васильева, К. А. Кириченко, М. А. Тимофеев, О. И. Грабельных // Физиология, биотехнология и биоинформатика растений и микроорганизмов - путь в будущее: к 85-летию Р. А. Карначук: материалы всероссийской научной конференции / ответственный редактор О. В. Карначук. Томск, 2022. С. 35-37. Текст: непосредственный.
- Таксономический список макроводорослей прибрежной зоны бухты Большие Коты и залива Лиственичный (Южный Байкал) / В. С. Вишняков, О. А. Тимошкин, Л. А. Ижболдина [и др.] // Известия Иркутского государственного университета. Серия: Биология. Экология. 2012. Т. 5, №. 3. С. 147-159. Текст: непосредственный.
- Разнонаправленные эффекты тебуконазол-содержащего протравителя семян "Бункер" на рост побегов и корней озимой пшеницы / О. И. Грабельных, Е. А. Полякова, А. В. Корсукова [и др.] // Известия Иркутского государственного университета. Серия: Биология. Экология. 2020. Т. 34. С. 3-19. Текст: непосредственный.
- Ижболдина Л. А. Атлас и определитель водорослей бентоса и перифитона озера Байкал (мейо- и макрофиты) с краткими очерками по их экологии. Новосибирск: Наука, 2007. 248 с. Текст: непосредственный.
- Сезонные изменения содержания фотосинтетических пигментов у красной водоросли Ahnfeltiopsis flabelliformis Японского моря / А. О. Кравченко, Е. С. Белоциценко, И. М. Яковлева [и др.] // Известия ТИНРО (Тихоокеанского научно-исследовательского рыбохозяйственного центра). 2011. Т. 166. С. 138-148. Текст: непосредственный.
- Свободнорадикальное окисление: учебное пособие / Ф. Е. Путилина, О. В. Галкина, Н.Д. Ещенко [и др.]; под редакцией Н. Д. Ещенко. Санкт-Петербург: Издво С.-Петерб. ун-та, 2008. 161 с. Текст: непосредственный.
- Bellincampi D., Dipierro N., Salvi G. [et al.]. Extracellular H2O2 induced by oligogalacturonides is not involved in the inhibition of the auxin-regulated rolB gene expression in tobacco leaf explants // Plant physiology. 2000. T. 122, №. 4. C. 1379-1386.
- Heath R. L., Packer L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation // Archives of biochemistry and biophysics. 1968. T. 125. № 1. C. 189-198.
- Vishnyakov V. S. Nomenclatural novelties in the endemic Lake Baikal genus Draparnaldioides (Chlorophyta, Chaetophoraceae) //Notulae Algrum. 2020. No. 158. P. 1-7.
