Взаимосвязь между параметрами стабильности и адаптивности сортов

Автор: Дьяков А.Б., Трунова М.В.
Статья в выпуске: 1 (142-143), 2010 года.
Бесплатный доступ
В связи с различными, нередко прямо противоположными представлениями авторов разных публика-ций о том, мерой каких биологических свойств генотипов являются их оценки коэффициентами регрессии урожаев на индексы среды bi, изучены причины варьирования этого параметра с использованием данных эко- логических сортоиспытаний сои. Экспериментально и теоретически доказано, что значение параметра bi i-того сорта практически полностью определяется величиной стандартного отклонения оценок его урожайности вследствие разных условий в опытах экологического сортоиспытания. Изучение зависимости величин пара-метра bi от индексов стрессоустойчивости сортов показало, что значения bi могут характеризовать различия генотипов не только по отзывчивости на благоприятные условия, но и по степени их устойчивости к стрессам.
Адаптивность, стабильность, отзывчивость, параметры оценок
Короткий адрес: https://sciup.org/142150936
IDR: 142150936
Correlation between parameters of ecological stability and adaptability of the cultivated plants varieties
Authors of various publications present different, frequently antipodal views on the following problem: measure of which biological traits of genotypes are their evaluations by the yield regression rates on bi environmental parameters. In connection with this problem factors of variation of this parameter with the help of soybean cultivars tests data were studied. It was experimentally and theoretically proved that value of bi parameter of the i-th cultivar is practically entirely determined by the value of the average divergence of its yield evaluations as a result of various conditions of ecological tests of cultivars. Studying the dependence of bi parameter values on the stress-resistance indices showed that bi can characterize the differences of genotypes not only by the sensitiveness to favorable condi-tions but also by their stress-resistance degree.
Текст научной статьи Взаимосвязь между параметрами стабильности и адаптивности сортов
Введение. Оценки урожайности сортов и гибридов растений зависят не только от потенциалов продуктивности генотипов, но и от условий внешней среды, в которых эти потенциалы реализуются. Поэтому изменчивость условий возделывания обусловливает ту или иную степень нестабильности оценок урожайности генотипов в соответствии с различиями их реакций на изменения внешних условий. Специфику таких реакций селекционер не может детально изучить по результатам предварительного и конкурсного сортоиспытаний (Бороевич, 1984, с. 327-328). Методы биометрической генетики не базируются на знании механизма взаимодействия генотип – среда, поэтому позволяют только измерять его проявления, но не способны обеспечить прогноз поведения генотипа в новых условиях (Аверьянова, Драгавцев, 1988).
Создаваемые сорта чаще оказываются невостребованными производством не из-за сниженного уровня потенциала продуктивности, а вследствие недостаточной экологической стабильности и адаптивности (Кадыров и др., 1984). Методы отборов на такие свойства в процессе селекции неэффективны, а рекордную адаптивность лучших сортов нельзя перенести в создаваемый сорт методом беккроссов (Крупнов, 1981; Кадыров и др. 1984). Для выявления степени стабильности и адаптивности созданных сортов проводятся их экологические испытания, по результатам которых вычисляются те или иные биометрические параметры, предложенные для оценки особенностей норм реакции генотипов на диапазон условий испытаний.
В соответствии с большим практическим значением определения стабильности и адаптивности предлагаемых производству сортов и гибридов постоянно увеличивается количество публикаций по этой теме, однако пропорционально возрастают различия как в принципах и методах оценок этих свойств, так и в биологической интерпретации значений вычисляемых параметров, что вызывает растерянность ученых при выборе методов оценок (Lin et al., 1986; Пешек и др., 1988). Даже в понятие «стабильность» часто вкладывается разный смысл, приводящий к путанице по этому вопросу в литературе, что особенно ясно выявляется при использовании разных способов количественного измерения стабильности (Одум, 1986, с. 65; Lin et al., 1986). Разная биологическая интерпретация одного и того же биометрического параметра также снижает эффективность его использования в селекции растений. Это можно показать даже на примере наиболее употребляемого параметра b i – коэффициента регрессии оценок урожайности x ij i-го генотипа на индексы среды x. j . K.W. Finlay и G.N. Wilkinson (1963) предложили этот коэффициент в качестве показателя адаптивности сорта, а S.A. Eberhart и W.A. Russell (1966) расценивали величину bi как параметр стабильности урожаев генотипа. В бывшем же СССР, а теперь в странах СНГ величины bi считают мерой пластичности генотипов (Пакудин, 1976; Пакудин, Лопатина, 1980; Комаров, Дубина, 1996; Сапега, 1997). Однако, если С. Бороевич (1984, с. 313) к числу пластичных относит сорта с b i <1,0, то в большем числе публикаций, например, в статье П.П. Литуна (1980), наоборот, высокопластичными считаются генотипы с b i >1,0, а в статье В.З. Пакудина и Л.М. Лопатиной (1980) в том же сборнике утверждается, что показателем высокой экологической пластичности генотипа является величина b i =1,0. Как и S.A. Eberhart и W.A. Russell (1966), многие зарубежные селекционеры, например D. Jowett (1972), считают оптимальным значение b i =1,0. После того, как академик Н.И. Вавилов (1966, с. 368, 433) определил показателем высокой пластичности сорта обширный размер ареала его возделывания, такой точки зрения придерживаются большинство отечественных селекционеров, в том числе академик В.С. Пустовойт (1966, с. 359), но сорт с показателем b i >1,0 не может быть высокопластичным в соответствии с определением этого понятия Н.И. Вавиловым. Академик П.П. Лукьяненко (1973, с. 116-117) считал показателем высокой пластичности способность сорта давать удовлетворительные урожаи в неблагоприятных условиях, а этому соответствуют величины bi<1,0.
Во многих публикациях значения bi>1,0 расцениваются как показатель хорошей отзывчивости генотипа на благоприятные условия, как характеристика сорта интенсивного типа (Пакудин, 1976; Стельмах, 1976; Вольф, Литун, 1978; Лещенко, Михайлов, 1978; Фукс, 1980; Сапега, 1997). В.Г. Вольф и П.П. Литун (1978) расценивают сорта с b i <1 как малоперспективные из-за слабой отзывчивости на комфортные условия. Однако A.A. Rosielle и J. Hamblin (1981) показали, что по величинам b i <1,0 можно идентифицировать генотипы, высоко толерантные к стрессам.
Хотя S.A. Eberhart и W.A. Russell (1966) считали величину параметра b i показателем степени стабильности генотипа, C.S. Lin с соавторами (1986), H.C. Becker и J. Leon (1988), И. Пешек и др. (1988) утверждают, что средовая дисперсия S i 2 (или стандартное отклонение S i ) является теоретически более обоснованным показателем стабильности, но она мало пригодна для селекционеров, которые связывают стабильность этого типа с низкой урожайностью. По мнению этих авторов, при селекции растений на стабильно высокую урожайность эффективней использовать коэффициент регрессии на индексы среды bi, который они относят к принципиально иной группе параметров.
Полагая, что углубленное понимание сути принципиальных различий показателей S i и b i позволит корректней использовать их при оценках генотипов, мы изучили характер связи между этими параметрами по данным экологических сортоиспытаний сои. Поскольку известно, что адаптивность сортов сои к климату юга России определяется их устойчивостью к летним засухам, результаты изучения по тем же данным связи между индексами засухоустойчивости сортов и их оценками коэффициентами регрессии на индексы среды должны были показать, возможна ли оценка адаптивности генотипов величинами параметра b i и в какой мере этот параметр является показателем отзывчивости сорта на благоприятные условия возделывания.
Материал и методы. Объектами исследования служили 2 набора южнороссийских сортов сои, экологические испытания которых проводили на центральной экспериментальной базе ВНИИ масличных культур (г. Краснодар). 6 сортов разной продолжительности вегетации изучали 7 лет – с 1999 по 2005 гг., а испытания 12 сортов четырёх периодов её селекции продолжались 9 лет – с 2001 по 2009 гг. Опыты закладывали в 4-кратной повторности на делянках с защитными рядами и учетной площадью 6,75 м2. Биометрическому анализу подвергали результаты оценок урожайности сортов. Экологические стандартные отклонения, коэффициенты корреляции и регрессии вычисляли общепринятыми методами. Индекс (коэффициент) засухоустойчивости определяли как выраженное в процентах отношение оценок урожайности сортов в условиях засухи к величинам их урожаев при хорошей влагообеспеченности, как это принято в исследованиях разных авторов (Vidal et al., 1981; Sagar et al., 1984; Мамонов, Ким, 1986). При этих вычислениях усредняли оценки урожайности каждого сорта за 3 наименее урожайных года и за 3 благополучных года.
Результаты и обсуждение. Обработка данных сортоиспытания 2001-2009 гг. выявила очень высокую зависимость оценок урожайности генотипов сои от индексов среды (табл. 1). У большинства изученных сортов эта связь выражалась значениями коэффициентов корреляции от r i =0,98 до r i =0,99. Однако степень изменения продуктивности тех же сортов с увеличением индексов среды варьировала в большей мере от b i =0,86 до b i =1,22. Ещё более значительной оказалась наследственная изменчивость индекса засухоустойчивости, величина которого составила в среднем 18,2 % у созданных в период 1959-1972 гг. сортов, отличающихся позднеспелостью и избыточной площадью листьев; 25,6 % у сортов сои довоенной селекции; 28,0 % у более скороспелых сортов, созданных с 1976 по 1987 годы, и 36,8 % у сортов последнего периода селекции, когда особое внимание уделялось повышению стабильности урожаев в разные по количеству осадков годы. Средние оценки нес т абильно с ти урожа е в за годы испытаний у сортов тех же периодов селекции составили s i=1,27, s i=1,25, s i=1,06 и s i=1,10. Это свидетельствует о том, что устойчивость к дефициту влаги и стабильность урожаев сои может быть повышена селекцией в двух направлениях. Сокращением длительности вегетации можно повысить вероятность избежания июльско-августовских засух, но у скороспелых сортов снижен потенциал урожайности, реализующийся в благоприятные годы. Отбор же на истинную засухоустойчивость позволяет сочетать этот признак с высоким потенциалом урожайности.
Таблица 1 – Параметры стабильности, устойчивости и потенциала урожайности сортов сои, вычисленные по данным экологического испытания 2001-2009 гг.
г. Краснодар
|
Сорт |
Год создания |
Стандартное отклонение, S i |
Коэффициенты связи урожаев с индексами среды |
Индекс засу-хоустойчи-вости, % |
Средний урожай семян за 3 благоприятных года, т/га |
|
|
корреляции, r i |
регрессии, b i |
|||||
|
Кубанская 276 |
1937 |
1,19 |
0,987* |
1,02 |
26,6 |
3,49 |
|
Кубанская 4958 |
1939 |
1,40 |
0,992* |
1,22 |
22,5 |
4,05 |
|
ВНИИМК 9186 |
1942 |
1,16 |
0,992* |
0,99 |
27,8 |
3,42 |
|
ВНИИСК 7 |
1959 |
1,26 |
0,986* |
1,09 |
18,7 |
3,37 |
|
Комсомолка |
1965 |
1,33 |
0,978* |
1,14 |
12,0 |
3,59 |
|
ВНИИМК 8 |
1972 |
1,22 |
0,984* |
1,05 |
23,9 |
3,60 |
|
Кубань |
1976 |
1,13 |
0,990* |
0,97 |
24,0 |
3,13 |
|
ВНИИМК 3895 |
1981 |
1,02 |
0,996* |
0,86 |
31,6 |
3,13 |
|
ВНИИМК 20 |
1987 |
1,04 |
0,990* |
0,89 |
28,5 |
3,02 |
|
Лань |
1993 |
1,13 |
0,982* |
0,95 |
29,3 |
3,35 |
|
Вилана |
1996 |
1,05 |
0,981* |
0,90 |
42,3 |
3,68 |
|
Рента |
2000 |
1,12 |
0,957* |
0,92 |
38,9 |
3,60 |
Примечание: коэффициенты корреляции переходят уровни существенности * – 0,1 %-ный
Сорта сои экологического испытания 1999-2005 гг. в большей степени различались по продолжительности вегетационных периодов. В число этих сортов был включен также самый адаптивный к климату юга России сорт Вилана. Поэтому действие этих двух факторов, влияющих на засухоустойчивость и потенциал урожайности, по данным этого испытания, проявилось в ещё большей мере (табл. 2). Кроме того, обнаружилось, что при наибольшей скороспелости снижается коэффициент корреляции ri оценок урожайности сорта с индексами среды вследствие несовпадения по времени критических периодов формирования урожая семян сортов, значительно различающихся по продолжительности вегетации. С увеличением числа дней от всходов до созревания значения параметра bi возросли вдвое. Такая же тенденция была обнаружена при экологических испытаниях различающихся по длительности вегетации 14 сортов и линий сои в более благоприятных условиях штата Айова США (Walker, Fehr, 1978).
Таблица 2 — Параметры стабильности, устойчивости и потенциала урожайности сортов сои, вычисленные по данным экологического испытания 1999-2005 гг.
г. Краснодар
|
Сорт |
Дней от всходов до созревания |
Стандартное отклонение, Si |
Коэффициенты связи урожаев с индексами среды |
Индекс засухоустойчивости, % |
Средний урожай семян за 3 благоприятных года, т/га |
|
|
корреляции, ri |
регрессии, bi |
|||||
|
Юг 30 |
95 |
0,70 |
0,882+ |
0,64 |
61,2 |
2,42 |
|
Быстрица 2 |
105 |
0,76 |
0,951* |
0,75 |
54,7 |
2,65 |
|
Руно |
106 |
1,00 |
0,986* |
1,03 |
47,7 |
3,04 |
|
Вилана |
116 |
1,11 |
0,985* |
1,14 |
48,1 |
3,76 |
|
Астра |
120 |
1,21 |
0,963* |
1,22 |
33,0 |
3,54 |
|
Лань |
117 |
1,18 |
0,980* |
1,21 |
38,0 |
3,63 |
Примечание: коэффициенты корреляции переходят уровни существенности + – 2 %-ный; * – 0,1 %-ный
Сложившемуся в специальной литературе представлению о принципиальном отличии от параметра b i и меньшей ценности для селекционеров показателей экологической вариабельности оценок урожайности генотипов Si2 или Si противоречат данные, приведенные в таблицах 1 и 2. И в том, и в другом случаях даже при беглом взгляде обнаруживается явная связь между величинами стандартных отклонений S i и коэффициентов регрессии на индексы среды b i . Оцененная коэффициентами корреляции, эта связь оказалась положительной и очень высокой: r=0,994 для испытаний 2001-2009 гг. и r=0,996 для опытов 1999-2005 гг. (значительно переходят 0,1 %-ный уровень существенности). Регрессионный анализ показал, что эта зависимость описывается уравнениями прямой линии, проходящей вблизи начала координат (рис. 1).
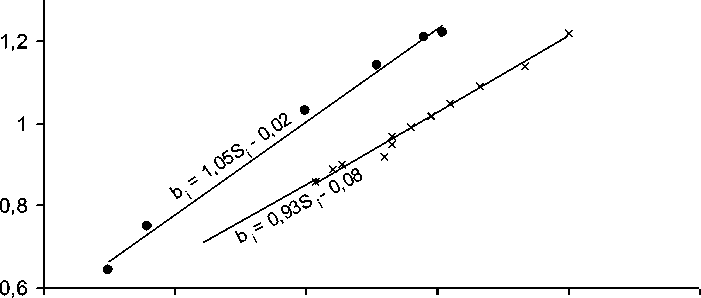
0,6 0,8 1
1,2 1,4 1,6
Параметр S i
Рисунок 1 – Регрессии значений параметра b i сортов сои на средовые стандартные отклонения оценок их урожайности S i по данным испытаний 1999-2005 гг. (•) и 2001-2009 гг. (×)
Важно выяснить, является ли обнаруженная зависимость (рис. 1) только особенностью данных наших испытаний сортов сои или в её основе лежит фундаментальная причина. С этой целью рассмотрим общую формулу вычисления коэффициента регрессии (Рокицкий, 1973, с. 151):
by/x = r Sy , Sx где by/x – коэффициент регрессии y на x; r – коэффициент корреляции между величинами y и x; Sy и Sx – стандартные отклонения величин y и x соответственно. При вычислениях параметра bi i-го сорта ri есть корреляция между величинами его урожаев и индексами среды; Sy есть Si урожаев этого сорта, а Sx – стандартное отклонение индексов среды x.j от центральной средней х.., поэтому Sx одинаково для всех генотипов одной серии испытания, т.е. для них Sx = const. Поэтому различия этих генотипов по значениям bi обусловлены только величинами Si и ri. Судя по величинам коэффициентов корреляции между Si и bi и по распределениям точек на графике (см. рис. 1), изменчивость bi определяется в основном вариацией значений Si, хотя коэффициенты корреляции ri между урожаями i-го сорта и индексами среды варьировали от ri=0,882 до ri=0,986 в испытании 1999-2005 гг. и от ri=0,957 до ri=0,996 в испытании 2001-2009 гг. Из этого следует, что при таких диапазонах вариации ri, которые позволяют вычислять bi, их величины почти полностью определяются значениями средовых стандартных отклонений Si. Следовательно, S.A. Eberhart и W.A. Russell (1966) с полным основанием определили bi параметром стабильности, а все иные интерпретации биологического содержания этого параметра нельзя считать обоснованными. Однако у параметра bi есть преимущество по сравнению с использованием Si или Si2: с его помощью можно оценить различия генотипов по потенциалам урожайности и устойчивости к стрессам путем вычисления математических ожиданий их урожайности при максимальном и минимальном значениях индексов среды. Для таких вычислений недостаточно знать только оценку bi, необходимо еще определить среднюю по этой серии испытаний урожайность генотипа, а также значения индексов среды.
Во многих публикациях величина коэффициента регрессии оценок урожайности i-го генотипа на индексы среды b i расценивается как показатель степени отзывчивости этого генотипа на благоприятные условия возделывания. Однако R.M. Habgood (1977) писал, что одинаково правильно считать параметр bi индикатором нежелательной для селекционера восприимчивости генотипа к стрессам, если нет доказательств, что в конкретном испытании проявился положительный отклик на благоприятные условия. Например, нашими исследованиями (Дьяков и др., 1997) было установлено, что значения bi для оценок урожайности гибридов и сортов подсолнечника тесно коррелировали с показателями степени восприимчивости их растений к поражению фомопсисом. Поскольку причиной наследственных различий по адаптивности является в основном разнообразие генотипов по степени устойчивости к основным абиотическим и биотическим стрессам конкретного местообитания, величина параметра bi должна отражать степень адаптивности в тех случаях, когда регрессия оценок урожайности i-го генотипа на индексы среды обусловлена его реакцией на неодинаковую степень проявления стресс-фактора в разных опытах экологического сортоиспытания.
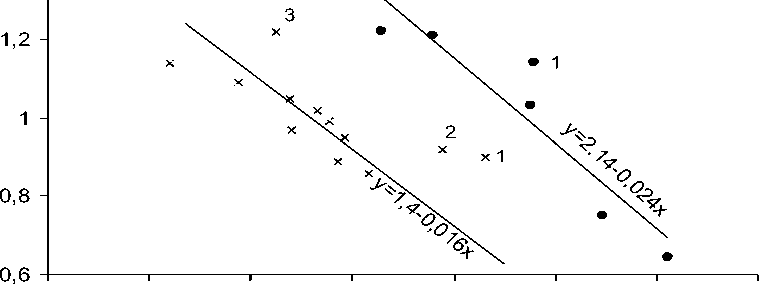
Исходя из того, что адаптивность сортов к климату юга России обусловлена степенью их устойчивости к засухам июля и августа, мы изучили характер регрессий величин параметра b i на индексы засухоустойчивости соответствующих сортов сои по данным таблиц 1 и 2. Эти зависимости оказались прямолинейными, отрицательными (рис. 2). Следовательно, в условиях данных сортоиспытаний сои важной причиной вариации значений bi являлись различия сортов по степени засухоустойчивости, а значит и по адаптивности к климату юга России. При этом оказалось, что точки сортов Вилана, Рента и Кубанская 4958 легли выше соответствующих линий регрессий. Обусловлено это высокими потенциалами урожайности этих сортов, реализующимися в благоприятные годы (см. табл. 1 и 2), что искажает оценки индексов засухоустойчивости, которые вычисляются как соотношение урожаев при засухах и в комфортных условиях.
Несмотря на то, что проведенный регрессионный анализ (рис. 2) выявил ограничение использования индекса засухоустойчивости, его результаты свидетельствуют о том, что при определенных условиях величина параметра b i может отражать степень адаптивности генотипа. А поскольку с параметром b i тесно коррелируют величины Si, их тоже можно использовать в таких ситуациях в качестве показателей степени адаптивности генотипа. Однако из приведенных в таблицах данных следует, что при этом сравнивать между собой можно только одинаковые по продолжительности вегетации генотипы.
0 10 20 30 40 50 60 70
Индекс засухоустойчивости, %
Рисунок 2 – Регрессии значений параметра b i сортов сои на индексы их засухоустойчивости по данным испытаний 1999-2005 гг. (•) и 2001-2009 гг. (×). Отклоняются от регрессий точки сортов Вилана (1), Рента (2) и Кубанская 4958 (3)
Выводы. 1. Доказана ошибочность представления о принципиальных различиях методов оценки взаимодействия генотип – среда величинами экологических стандартных отклонений S i и коэффициентов регрессии оценок урожайности на индексы среды b i .
-
2. Оба параметра можно использовать для оценки отзывчивости сортов, если установлено, что главной причиной средовой вариации урожаев в опытах экологического испытания являлась реакция генотипов на разные уровни благоприятности условий внешней среды.
-
3. Величинами параметров Si и bi можно оценивать степень адаптивности сортов, если основной причиной средовой вариации оценок урожайности в опытах была реакция генотипов на типичные для местообитания абиотические и биотические стрессы.
-
4. Индекс устойчивости к засухе (и другим стрессам) можно использовать лишь при сравнении генотипов с одинаковыми потенциалами урожайности, реализующимися в благоприятные годы.
-
5. Во всех ситуациях использования изученных параметров сравнивать между собой следует сорта одинаковой продолжительности вегетации.
