Who were the Denisovans?

Author: Derevianko A.P., Shunkov M.V., Kozlikin M.B.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Paleoenvironment, the stone age
Article in issue: 3 т.48, 2020.
Free access
Short address: https://sciup.org/145145503
IDR: 145145503 | DOI: 10.17746/1563-0110.2020.48.3.003-032
Text of the review article Who were the Denisovans?
In the last decade, numerous experts in human evolution and the origins of modern humans (physical anthropologists, specialists in paleogenetics, and archaeologists) have been trying to answer the question as to who Denisovans were. The discovery of Denisovan remains was a complete surprise for all of them. In 2010, for the first time, on the basis of the analysis of mitochondrial DNA extracted from a finger phalanx, found in the Pleistocene deposits of Denisova Cave (Fig. 1) in the northwestern part of the Russian Altai, dating to the Initial Upper Paleolithic, a new hominin taxon, genetically very different from both Homo sapiens and H. neanderthalensis, was described.
Since the early 1980s, specialists from the Institute of Archaeology and Ethnography SB RAS have excavated about two dozen stratified Paleolithic sites in the Altai, revealing several meters thick Pleistocene sequences consisting of up to ten habitation horizons each. Field and laboratory work is being jointly conducted by archaeologists, geologists, paleogeographers, geochronologists, paleontologists, physical anthropologists, paleogeneticists, etc. from major Russian and foreign research centers. To date, a large body of information has been accumulated,
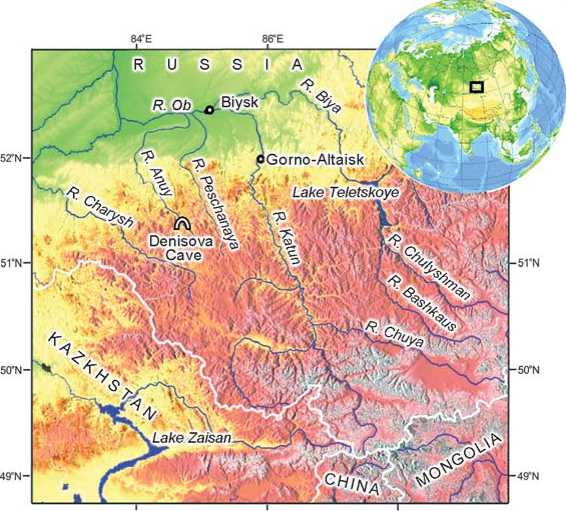
Fig. 1. Location of Denisova Cave.
evidencing the evolution of the Paleolithic cultural traditions in the Middle and Upper Pleistocene.
The most representative materials come from Denisova Cave (Fig. 2), where thirteen habitation layers with Paleolithic artifacts have been described. Owing to an unusually complete stratigraphic sequence, the evolution of the material culture of the cave dwellers has been traced over some 300 thousand years.
Especially intriguing were findings relating to layer 11 of the cave, characterized by the Early Upper Paleolithic blade industry, as well as numerous bone implements, and ornaments made of bone and semi-precious stone. These deposits span the chronological interval between 50–40 ka BP, implying that in the Altai, the Upper Paleolithic emerged earlier than in Europe or Africa. The appearance of stone and bone tools from layer 11 initially suggested their association
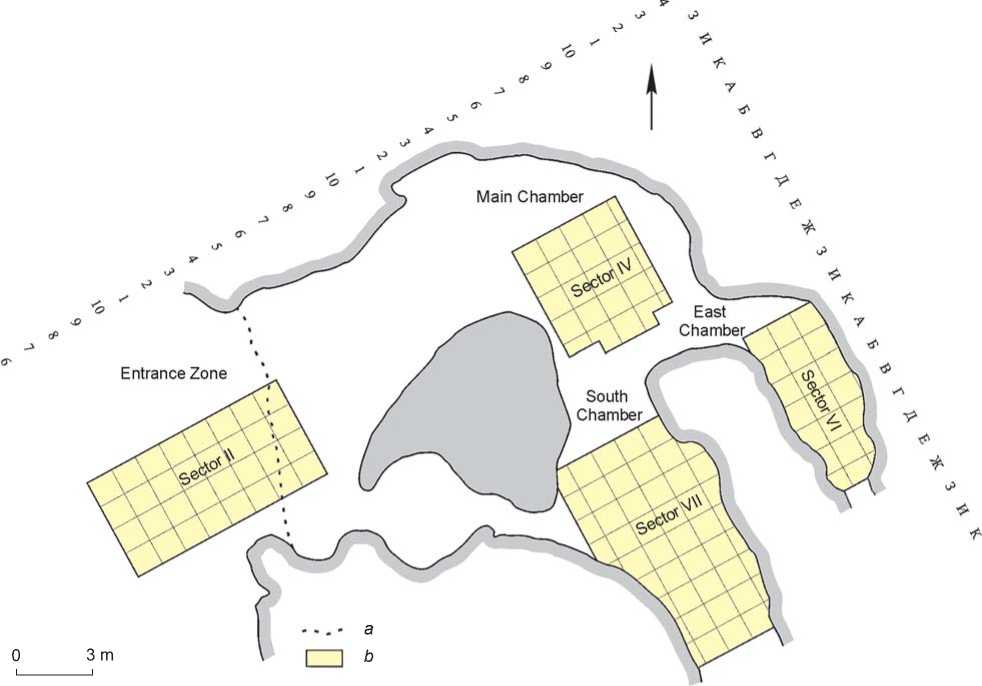
Fig. 2. Plan of Denisova Cave.
a – drip-line; b – excavations of Pleistocene deposits.
with H. sapiens , which was somewhat too early for anatomically modern humans to have appeared in Siberia.
In 2008, a distal phalanx of a girl’s little finger was found in the East Chamber of Denisova Cave, in layer 11.2. Part of it was handed to Svante Pääbo, the Head of the Department of Genetics at the Max Planck Institute of Evolutionary Anthropology. Previously, geneticists associated with this institute had demonstrated that the Okladnikov Cave dwellers were Neanderthals (Krause et al., 2007). The sequencing of the mitochondrial and then the nuclear DNA from the Denisova phalanx suggested that the bone belonged to a hitherto unknown hominin, who was termed Denisovan after the eponymous site (Krause et al., 2010). According to the genetic study of the aDNA extracted from the left upper molar from lithological unit 11 in the South Chamber of the cave, the young male to whom the tooth belonged was a Denisovan too (Reich et al., 2010). This tooth shows neither Neanderthal nor modern human apomorphies, suggesting that the Denisovan evolutionary trajectory was different from those of both H. neanderthalensis and H. sapiens .
The analysis of nuclear genomes of the newly described hominins suggests that they were a sister group of Neanderthals, i.e., their common ancestor was a sister taxon of anatomically modern humans. This means that at least two hominin taxa coexisted with H. sapiens in Eurasia: Neanderthals in its western part and Denisovans in its eastern part.
Multidisciplinary studies of materials from Denisova Cave suggest that the evolution of the Paleolithic tradition in that area resulted in the autochthonous emergence of the Upper Paleolithic culture on a Middle Paleolithic basis, indicating not only cultural but biological continuity as well. The analysis of human aDNA from fossil remains unearthed in the cave demonstrates that Denisovans were associated not only with lithological unit 11 and its Early Upper Paleolithic industry, but also with the middle part of the sequence (the boundary of layers 11.4 and 12) and its lower part (layer 22.1), representing various stages of the Middle Paleolithic (Sawyer et al., 2015; Slon et al., 2017b). It follows that those who manufactured both Middle Paleolithic and Early Upper Paleolithic tools were Denisovans. Also, judging by hominin fossils from layers 12 and 11.4 in the East Chamber of the cave, Denisovans coexisted with Neanderthals during the Middle Paleolithic, and occasionally hybridized with them (Mednikova, 2011a, 2013; Prüfer et al., 2014; Slon et al.,
2017a, 2018). Despite major stratigraphic gaps, archaeological finds from the Denisova sequence testify to cultural continuity in the evolution of the lithic industries from the Early Middle Paleolithic to the mid-Upper Paleolithic. Although Neanderthal fossils were found in Denisova Cave, no evidence of the Mousterian tradition has so far been detected there, which may tentatively be explained by the interbreeding between Denisovans and Neanderthals. This hypothesis has been supported by the analysis of DNA extracted from a girl’s bone from layer 12.3, revealing mixed female Neanderthal and male Denisovan ancestry (Slon et al., 2018).
Geochronologyof the Denisova Pleistocene deposits
Consistent studies of key topics of the evolution of Homo genus, including the time and routes of initial peopling of the Earth, evolution of the anatomically modern humans, and the development of primitive culture, are possible only on the basis of reliable chronostratigraphic data from the Paleolithic sites under study. These data are especially important in the studies of stratified sites with the long cultural sequence, such as Denisova Cave. A significant series of dates have been obtained in Russian and foreign laboratories, using various techniques, in the course of comprehensive studies of the Denisova Pleistocene deposits. The results of radiocarbon and optical dating carried out recently using the methods of the accelerator mass spectrometry (AMS) and optically stimulated luminescence (OSL) in laboratories of the Oxford and Wollongong Universities are crucially important for the study of geochronology of Denisova Cave.
The most complete chronostratigraphic sequence of the cave sediments was established using the OSL-technique by the team of specialists headed by Profs. R. Roberts and Z. Jacobs from the Center for Archaeological Science at the University of Wollongong (Jacobs Z. et al., 2019). Australian researchers, having profound experience of field works at Paleolithic sites in Africa and Eurasia, during five years have been collecting samples from Denisova Pleistocene deposits for the OSL-dating. On the basis of results of the optical dating of 103 deposits samples, they have produced chronological models of the Pleistocene sequence in three chambers of the cave. Dating of 92 samples was executed on the basis of measurements of more than 280 thousand separate seeds of quartz and potassium-fortified feldspar;
11 samples were dated using the analysis of multigrain aliquots of K-feldspar. Optical dates were calculated using four methods of assessment of the equivalent dose ( De ), and were used for the development of Bayesian model of sedimentation chronology in each cave chamber, with the aim of correlation of layers of the stratigraphic sequences, as well as establishing the complex chronological and climatic interval of the cave peopling by hominins.
At present, the Pleistocene deposits in the Main and East Chambers of the cave are the best studied in terms of archaeology, lithology, biostratigraphy, and chronostratigraphy (Fig. 3). The earliest OSL-date was established for the basal deposits in the East Chamber: 508 ± 40 ka BP for the top part of layer 17.2, and 305 ± 37 ka BP for the bottom part of layer 17.1; as well as 366 ± 43 ka BP for the bottom of layer 22.3 in the Main Chamber.
The earliest archaeological evidence of the cave is associated with the Early Middle Paleolithic artifacts recovered from the dense, light to dark brown loams in layer 22 in the Main Chamber, with the top part of the layer having the OSL-date of 287 ± 41 ka BP. Layer 22 is overlain by culture-bearing horizons 21 and 20, dating to the range from 250 ± 44 to 170 ± 19 ka BP. In the East Chamber, the Early Middle Paleolithic is represented by the dark gray sediments of layers 15 and 14, dating to the range from 203 ± 14 to 187 ± 14 ka BP. Available paleogeographic data of this sedimentation suggest that there were two warm and one comparatively cold climatic phases in the second half of the Middle Pleistocene corresponding to MIS 9–7.
East Chamber
Main Chamber

аbс de (D f ®g DNA h i
1 m
Fig. 3. Pleistocene deposits in the East Chamber and Main Chamber of Denisova Cave. Beginning and end of sedimentation is modeled using the Bayesian method on the basis of optical time estimates (after (Jacobs Z. et al., 2019)).
Climatic conditions: a – relatively warm; b – relatively cold; c – no data; d – sedimentation gap. Fossils: e – Denisovan, f – Neanderthal, g – hybrid. DNA from deposits: h – Denisovan; i – Neanderthal.
UP – Upper Paleolithic; EUP – Early Upper Paleolithic; IUP – Initial Upper Paleolithic; MP – Middle Paleolithic; EMP – Early Middle Paleolithic.
The next stage of the Middle Paleolithic is illustrated by the lithic industries from layers 19–14 in the Main Chamber and layers 13–11.4 in the East Chamber. These were OSL-dated to 156 ± 15 to 105 ± 11 / 97 ± ± 11 ka BP. The available chrono- and biostratigraphic data suggest that these deposits were formed after a long sedimentation gap during the last glaciation in the Middle Pleistocene and in the period of the last interglacial in the Early Upper Pleistocene, which periods correspond to MIS 6 and 5.
The terminal stage of the Middle Paleolithic is represented by the lithic industries of layer 12 in the Main Chamber and layer 11.3 in the East Chamber. The OSL age of these deposits has been established in the range of 80 ± 10 to 58 ± 6 ka BP, which corresponds to the cold period of MIS 4.
The thick series of the Middle Paleolithic cultural deposits in the Main Chamber of the cave was covered upon the long-lasting sedimentation gap with layer 11, and in the East Chamber (also after sedimentation gap) with layers 11.2 and 11.1, containing the Early Upper Paleolithic artifacts. Over 50 dates obtained using the radiocarbon and OSL analysis are available for these deposits. The AMS-dates for the upper part of the Pleistocene deposits were obtained from the charcoal and bones samples bearing signs of working, at the Research Laboratory for Archaeology and the History of Art (University of Oxford), by Profs. T. Higham and K. Douka. For the recent five years, these researchers have participated in the multidisciplinary studies at Denisova Cave (Douka et al., 2019).
The most early Upper Paleolithic complex, including (apart from lithic artifacts) a bone needle; ornaments of bone, semi-precious stone, and ostrich eggshell, as well as a distal phalanx of a little finger of the Denisovan Denisova 3 , was associated with layer 11.2 in the East Chamber. The OSL age of this complex is 63 ± 6 to 55 ± 6 ka; the AMS-dates are in the range of 50,300 ± 2200 years BP (OxA-V-2359-16) to 35,400 ± 900 years BP (OxA-30005).
There are several dates available for various parts of the cave deposits: the OSL-date of 49 ± 6 to 38 ± ± 9 ka BP and AMS-date of 47,900 ± 3100 years BP (OxA-29855) for layer 11.1 in the East Chamber; AMS-dates of 48,900 ± 1800 years BP (OxA-V-2359-18), 51,200 ± 2200 years BP (OxA-V-2359-17), 48,650 + 2380/– 1840 years BP (KIA 25285 SP 533.D19) for layer 11 in the South Chamber. The deposits of layer 11 in the Main Chamber of the cave are comparatively younger: the age of the five lithological sublayers (11.5–11.1) was determined in the range of 44 ± 5 to 38 ± 3 ka BP using OSL technique; the AMS-dates fall within the range from >50,400 years BP (OxA-34728) to 32,150 ± 450 years BP (OxA-33086). In addition, bone artifacts associated with the stone tools within this layer were directly dated. The bone point from layer 11.4 in the Main Chamber was dated to 42,660–48,100 cal years (OxA-30271); the elk tooth pendant from layer 11.2 in the East Chamber to 42,450–49,710 cal years (OxA-30963) (Ibid.). These assessments suggest that the said artifacts can be attributed to the oldest items of this type discovered so far in Northern Eurasia.
The top part of the Pleistocene deposits in Denisova Cave contains lithological layer 9 with the Middle Upper Paleolithic industry. The age of this layer in the Main Chamber was assessed as 36 ± 4 to 21 ± 8 ka BP using the OSL technique.
The geochronology of the anthropological remains discovered in the cave is of special interest. Their probable age was assessed using the Bayesian method, based on the chronometric (OSL- and 14C-dates), stratigraphic, and genetic data, for which purpose the chronological models were developed (Ibid.). The modeling of the oldest bone of the Denisovan Denisova 2 from layer 22.1 in the Main Chamber produced the date of 122,700–194,400 years. The date of the molar of the Denisovan Denisova 8 , recovered at the border of layers 12 and 11.4 in the East Chamber is estimated in the range of 105,600–136,400 years. The modeled age of the youngest bone of the Denisovan Denisova 3 from layer 11.2 in the East Chamber is 51,600– 76,200 years. Molar Denisova 4 from layer 11.1 in the South Chamber differs from the sample Denisova 3 only in two mtDNA mutations; hence, it has the same age.
The Middle Paleolithic layers in the East Chamber also yielded anthropological remains of Neanderthals and a bone of the girl-hybrid of a Neanderthal mother and a Denisovan father. The Neanderthal samples Denisova 5 and Denisova 15 from lithological layer 11.4 were dated to 90,900–130,000 years, and the phalanx Denisova 9 from layer 12.3 to 119,100–147,300 years. The bone Denisova 11 belonging to the daughter of Neanderthal mother and Denisovan father was identified among the indeterminable bone fragments from layer 12.3; the modeled age for this bone was 115,700–140,900 or 79,300–118,100 years.
The Neanderthal DNA was recorded in the deposits of lithological layers 19, 17, and 14 in the Main Chamber, with the OSL-age from 151 ± 17 to 97 ± 11 ka, as well as layers 14 (193 ± 12 to 187 ± 14 ka) and 11.4 (105 ± 11 ka) in the East Chamber. The Denisovan DNA was discovered in the deposits of layer 15 in the East Chamber, dated to 203 ± 14 to 197 ± 12 ka BP.
The modeled dates of the anthropological fossils are generally well correlated with the optical age of the cave deposits. The only exceptions are the molar Denisova 2 from layer 22.1 in the Main Chamber, and possibly the bone of the hybrid Denisova 11 from layer 12.3 in the East Chamber. Subsequent studies will provide additional data explaining the divergence between the modeled and the optical age of these samples; although, this partial discrepancy does not affect the general pattern of the cave peopling. The modeled age of the oldest Denisovan fossils suggest that this population emerged in the south of Siberia as early as 195 ka BP, and according to the OSL-dates of the sediments ca 300 ka BP. Judging by the youngest bone remain of Denisovans, this population survived in the Altai until ca 50 ka BP. The age of the Middle Paleolithic layers of the cave, yielding Neanderthal remains, including the bone revealing mixed female Neanderthal and male Denisovan ancestry, falls within the range of 193 ± 12 to 97 ± 11 ka BP. According to these data, both populations inhabited the northwestern Altai for a long time, had contacts, and hybridized with each other.
Hominin fossils
The first hominin fossil was discovered in the Pleistocene layers of Denisova Cave in 1984. This was a deciduous molar from lithological layer 22.1 in the Main Chamber, later named Denisova 2 (Shpakova, Derevianko, 2000). In the same year, layer 12 in the Main Chamber yielded a tooth, which was initially identified as a permanent upper incisor of a hominin (Turner, 1990; Shpakova, Derevianko, 2000), but later, on the basis of comparative analysis, was attributed to a bovid (Viola et al., 2011). In 2000, a permanent upper molar, named Denisova 4, was unearthed from layer 11.1 in the South Chamber of the cave; in 2008, a distal phalanx of a girl’s little finger (Denisova 3) was found in layer 11.2 of the East Chamber; and in 2010, fragments of a crown of the permanent upper molar (Denisova 8) were discovered in the bottom part of lithological layer 11.4, bordering with layer 12. The sequencing of the mitochondrial and nuclear DNA from the Denisova 3 phalanx and then from the Denisova 2, 4, and 8 molars, carried out under the guidance of Svante Pääbo at the laboratory of the Max Planck Institute of Evolutionary Anthropology, suggested that the fossils represent a previously unknown hominin species, named Homo altaiensis alias Denisovan. The new taxon was initially described on the basis of genetic rather than morphological criteria (Krause et al., 2010; Reich et al., 2010; Meyer et al., 2012; Sawyer et al., 2015; Slon et al., 2017b).
In 2010–2012, Neanderthal fossils were discovered in the East Chamber: proximal phalanx of the left little toe ( Denisova 5 ) and a morphologically indeterminable bone fragment ( Denisova 15 ) in layer 11.4, a distal phalanx of the third or fourth digit of the left hand in layer 12.3 ( Denisova 9 ) (Mednikova, 2011a, 2013; Prüfer et al., 2014; Slon et al., 2017a), and a morphologically indeterminable bone fragment belonging to the F1 hybrid of a Neanderthal mother and a Denisovan father ( Denisova 11 ) (Slon et al., 2018).
The identification of Denisova 5 and 9 phalanges as those of Neanderthals was based on the morphological and genetic analyses, whereas the generic and specific attribution of morphologically indeterminable bone fragments Denisova 11 and 15 became possible only after the ZooMS analysis followed by the sequencing of aDNA (Brown et al., 2016). The preservation of the endogenous DNA in fossils from Denisova Cave is generally very good. In certain specimens, its content exceeded 70 % (Ibid.), and that of bacterial DNA relating to microbes existing on bones after burial is 30–40 % (Reich et al., 2010). Normally, less than 1 % of endogenous DNA can be extracted from Late Pleistocene fossils (Viola, Pääbo, 2013).
In sum, Denisovan fossils available for morphological analysis to date include a distal phalanx of the hand and three teeth (Fig. 4).
Denisova 3 . Distal phalanx of the little finger of a juvenile from layer 11.2 in the East Chamber. In order to conduct the genomic analysis at two independent laboratories, the specimen was cut into two parts. The initial analysis of the proximal part, carried out at the Max Planck Institute of Evolutionary Anthropology, showed that the bone had been distally broken ~2 mm below the unfused proximal epiphyseal line. In modern man, the proximal epiphyses of distal phalanges normally begin to fuse with the shafts at the age of 13.5 years in females and 16 years in males, so the individual must have been younger. The age estimate is not quite accurate, but judging by the maximum width of the proximal shaft (7.5 mm) and by the maximum height (5.1 mm), this was a child no older than 6–7 years of age (Reich et al., 2010). Later, at the Institute Jacques Monod in Paris, a group of researchers headed by E.A. Bennett and E.M. Geigl, combining the photographs of the distal part of the phalanx and the three-dimensional model of the proximal parts (epiphysis and remains of the dorsal shaft), generated a virtual reconstruction
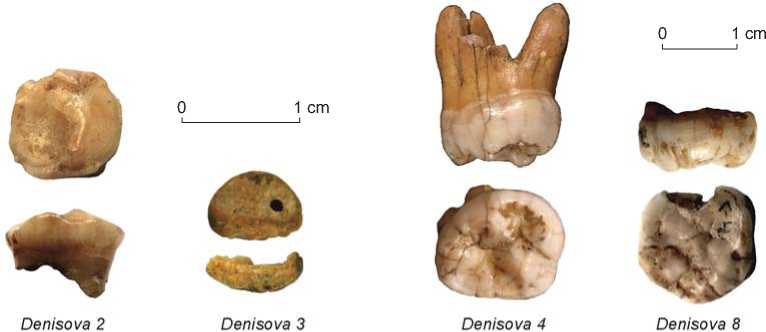
Fig. 4. Denisovan fossils.
of the entire phalanx in the dorsal and palmar views (Bennett et al., 2019). As the morphometric analysis of the reconstructed specimen demonstrated, both the size and the shape of the phalanx are within the variability limits in anatomically modern humans, thus linking Denisovans to H. sapiens . Because the distal phalanges of the little fingers of Neanderthals are markedly different, showing distinct apomorphies, a revision of micro-CT scans and photographs of the proximal fragments (articular surface and the semiring representing the dorsal half of the diaphysis) was conducted, and photographs of the distal fragment were compared with distal phalanges of Pleistocene hominins and recent humans at various developmental stages.
The comprehensive analysis has demonstrated that the phalanx belonged to an adolescent female, whose age at death, judging by modern standards, was approximately 13–16 years. In terms of maximum length, Denisova 3 is closest to the phalanges of modern humans. The asymmetry of the ungual tuberosity and the shaft’s curvature in the dorsal view suggest that the specimen may be from the right hand. The comparison of morphological information relating to Denisova 3 with that relating to Neanderthals, early anatomically modern humans, and recent modern humans from France and Belgium dating to the periods from the Neolithic to the Middle Ages links it to modern humans and opposes it to Neanderthals. Thus, while the nuclear DNA of this individual is closer to that of Neanderthals, the morphology of her hand bone is essentially modern (Ibid.).
The dental morphology of Denisovans, in contrast, is more archaic. The descriptions of teeth from various stratigraphic horizons of the cave are given below in descending order of their geological age.
Denisova 2 . Deciduous left lower second molar (dm2) from layer 22.1 in the Main Chamber. Initially, this was described as a deciduous right lower molar (Turner, 1990), but later the attribution was changed (Shpakova, Derevianko, 2000). The crown is completely abraded (grades 5–6), the roots are missing. In modern populations, similar changes in dental structure are typical of the age 10 ± 2.5 years. The occlusal surface has two abrasion platforms: anterior, smaller in size (1/3 S), inclined toward the mesial edge of the crown, and posterior (2/3 S), with a marked lowering of the level toward the distal edge and a small cup-like depression in the disto-lingual part. The enamel cingulum is 1–2 mm high, the cervix and part of the cervical portion of the mesial root are preserved. On the vestibular part of the crown, there are large dents in the enamel, and smaller ones in the disto-lingual corner of the crown. The anterior abrasion platform of the occlusal surface is delimited by a small preserved stretch of the first furrow on the exterior side. A large distal contact-facet is situated very low and evidences a prolonged and strong pressure from the crown of the first permanent molar, which had not yet erupted to its maximum height. In modern children, this tooth erupts from the alveole at 6 ± 2 years of age. Given the degree of abrasion, and disregarding the existing hypothesis about the earlier dental development, the child’s age at death (or at the time when the tooth was lost) can be estimated at approximately 10 years. However, the low position of the distal facet, implying that the position of the first permanent molar was low, suggests either that the child’s age was about 6–7 or that the complete eruption of the deciduous molar from the alveole was retarded. Given these estimates relating to two sets of teeth, the most acceptable estimate of age at the time of tooth loss is 7–8 years by modern standards (Ibid.).
According to other experts, the age of Denisova 2 might correspond to 10–12 years in modern children (Slon et al., 2017b).
Denisova 8 . Permanent left upper third molar (M3) from the bottom part of layer 11.4 near the border with layer 12 in the East Chamber. On the basis of the shape of the crown and the presence of a distinct oblique crest (a trait peculiar to upper molars) this tooth has been identified as the upper molar of a male (Sawyer et al., 2015; Zubova, Chikisheva, Shunkov, 2017). The anterior half of the crown is worn off; the protocone reveals a small exposed area of dentine, whereas no wear is seen on the distal part of the crown. The absence of the distal interproximal facet suggests that this is either the third or the second molar (in the latter case, the third molar had not yet erupted). Normally, by the age when upper second molars in Neanderthals and H. heidelbergensis showed the same degree of eruption as in Denisova 8 , the adjoining third molars had already erupted from the alveole and the interproximal facets were present. Therefore, the specimen might have been the second molar of an individual with a congenital absence of M3. This condition has been evidenced in late Asian H. erectus and Middle Pleistocene hominins.
Denisova 8 is very large—its length is more than three standard deviations larger than that of Neanderthal and modern human teeth, falling in the variation range of Pliocene hominins. Only two Late Pleistocene individuals exhibit third molars of comparable size: one is the Early Upper Paleolithic anatomically modern human Oase 2 in Romania (Trinkaus et al., 2003; Trinkaus, 2010), the other, a hominin with a mosaic morphology Obi-Rakhmat-1 in Uzbekistan (Glantz et al., 2008; Bailey et al., 2008).
Denisova 4 . Permanent left upper second or third molar (M2/3) from layer 11.1 in the South Chamber (Reich et al., 2010; Zubova, Chikisheva, Shunkov, 2017). The tooth evidently belonged to a young male. Its preservation is very good except for the apical part of the disto-buccal root. The crown, intersected by several cracks, is very high and has bulging buccal and lingual walls. In the occlusal view, the crown is rounded and tapers distally. It is slightly skewed lingually in the distal part, but is morphologically different from the crowns of the Neanderthal upper molars with their lingually projected hypocones and rhomboid shape.
The roots are relatively short (the length of the lingual root is 12.4 mm from the cervix, and that of the mesio-buccal root, 12.7 mm). The lingual root is very robust and lingually widened, and the two buccal roots are only slightly divided. The crown is very large (mesio-distal diameter, 13.1 mm, bucco-lingual diameter, 17.7 mm). As a third molar, its size exceeds most standards for fossil hominin taxa except H. habilis and H. rudolfensis, and is only comparable with Australopithecine standards. As a second molar, it is larger than those of Neanderthals or early modern humans, but smaller than those of H. erectus and H. habilis.
In sum, the morphological features of the Denisovan teeth indicate a very conservative evolutionary model, supporting the idea that a distinct hominin population existed in the Altai, differing from both H. sapiens and H. neanderthalensis not only genetically but morphologically as well.
Findings of genomic studies
The study of ancient DNA has demonstrated that Denisovan genomes differ from the standard human genome by 11.7 %, whereas the difference between the genomes of Neanderthals from Vindija, Croatia, and modern humans is 12.2 %. In terms of nuclear DNA, Denisovans and Neanderthals are sister taxa, opposed to H. sapiens (Reich et al., 2010). Genomic studies indicate the presence of 1.5–2.1 % of Neanderthal ancestry in the mitochondrial and nuclear genomes of modern non-Africans. Denisovans, on the other hand, did not participate in the supposed admixture between Neanderthals and anatomically modern humans, but 3–6 % of their ancestry is present in the gene pool of modern inhabitants of Southeast Asia, Australia, and Oceania (Reich et al., 2011).
The study of mitochondrial genomes suggests that the Denisovans separated from the Neanderthals ~640 ka BP, and from H. sapiens , ~1 Ma BP (Reich et al., 2010). On the basis of the analysis of nuclear DNA, on the other hand, the Denisovan-Neanderthal split occurred 430 ka BP or so (Meyer et al., 2014), whereas their common ancestors separated from those of H. sapiens about 800 ka BP (Meyer et al., 2012).
The results of another study are compatible with two scenarios of hominin evolution in the Pleistocene (Prüfer et al., 2014). According to the first, the common ancestor of H. sapiens , Neanderthals, and Denisovans lived ca 553–589 ka BP, and that of Neanderthals and Denisovans, 381 ka BP. According to the second scenario, the common ancestor of all the three species lived 550–765 ka BP, and the split between Neanderthals and Denisovans took place 445–473 ka BP.
The lithostratigraphic and geochronological studies of layers where the fossils were found in Denisova Cave attest to the prolonged existence of Denisovans in the Altai. After comparing the number of nucleotide substitutions in all Denisovan mitochondrial genomes and the mutation rate of modern human mtDNA, it was concluded that the Denisova 8 specimen is 60 thousand years older, and Denisova 2 100 thousand years older than the Denisova 3 and Denisova 4 specimens (Sawyer et al., 2015; Slon et al., 2017b).
Thus far, remains of Denisovans have been discovered only in one cave. However, judging by complete mtDNA sequences, the level of their genetic diversity is higher than that in seven Neanderthals from various regions of Western and Central Europe, while being lower than in modern humans (Sawyer et al., 2015). Consequently, the Denisovan population could have been larger and more diverse than the Neanderthal population; moreover, it had an extremely wide geographic distribution—from North Asia to the tropical zone of East and Southeast Asia (Meyer et al., 2012; Prüfer et al., 2014).
The Denisovan genomes included alleles that, in modern humans, control dark skin, brown hair, and brown eyes (Meyer et al., 2012). Also, the high-coverage genome of the individual whose DNA was extracted from the Denisova 3 phalanx revealed a component received from an unknown hominin, whose ancestors separated from those of Neanderthals, Denisovans, and modern humans 1–4 Ma BP (Prüfer et al., 2014). This ancestry is apparently present in several Denisovans in various proportions. Because the nuclear genome of Denisova 8 is much further from the Denisova 3 genome than from that of Denisova 4 , the Denisova 8 population, which existed earlier, could have been the carrier of such a component, controlling, for instance, large size (Sawyer et al., 2015).
To evaluate the taxonomic position of the Denisovans, it is important to note that the genomes of modern Australian aborigines and Oceanians contain 5–6 % or so of Denisovan ancestry, whereas in American and mainland Asian natives its proportion is minute—just 0.2 % (Prüfer et al., 2014). The presence of the Denisovan genetic legacy in the modern human gene pool suggests that during the early modern human dispersal in Asia, 80–50 ka BP, hybridization occurred, and that the hybrids were fertile. Denisovans and early anatomically modern migrants from Africa, then, were rather closely related, and reproductive barriers between them, if any, were relatively weak.
Therefore, all available data suggest that Denisovans might be a population within the species H. sapiens, and that hominins associated with the Denisovan Upper Paleolithic tradition, which emerged some 50 ka BP, can be termed H. sapiens altaiensis*.
The subspecific status of Upper Paleolithic Denisovans, reflected by the trinomen H.s. altaiensis , is supported by several considerations relating to modern human origins. Having migrated from Africa to Asia, early H. sapiens came in contact with the native populations such as Denisovans, whose immune systems were better adapted to resist local pathogenic microorganisms. Through admixture with natives the immigrants received genes enhancing adaptation to local ecological conditions, protecting them from diseases, and reinforcing their immune systems.
The part played by Denisovans in the adaptive introgression is evidenced by human leukocyte antigens (HLA) present in modern humans, helping the immune system to recognize and combat pathogenic organisms. Judging by the genome of the Denisova 3 individual, represented by a phalanx, one of the highly important components of the immune system (the HLA-B*73 allele) was inherited by modern humans from the Denisovans (Abi-Rached et al., 2011).
An illustrative example of adaptive introgression is the adaptation of Tibetans to hypoxia, caused by deprivation of oxygen supply at extreme altitudes. Whereas in plains dwellers, acclimatization to high mountain conditions is accompanied by increased hemoglobin levels, resulting in a greater risk of thrombosis, Tibetans show a comparatively low hemoglobin level and, accordingly, lower risk of cardiovascular diseases. Tibetan females are more fertile than those living on the plains; they do not suffer from pre-eclampsia, and the weight of their infants at birth is more stable.
One of the peculiar genetic features of Tibetans is the hypoxia pathway gene, EPAS1 , responsible for adaptation to low oxygen supply at high altitudes. The sequencing of the region around this gene in Tibetan and Han individuals, in 26 other genetically diverse populations from the Human Genome Diversity Panel, and in fossil hominins demonstrated that EPAS1 gene variation is found only in Denisovans and Tibetans. Its frequency is Han Chinese is quite low, and in the other modern peoples it is absent. Furthemore, comparison of DNA of Denisovans and Tibetans showed that the latter are genetically much closer to Denisovans than other modern humans, and that archaic, specifically Denisovan introgression helped them to adapt to high altitudes (Huerta-Sánchez et al., 2014).
Evidence of adaptive introgression from Denisovans into humans was also found in other populations of South, East, and Southeast Asia (Skoglund, Jakobsson, 2011; Reich et al., 2011; Lalueza - Fox, Gilbert, 2011; Prüfer et al., 2014; Qin, Stoneking, 2015; Sankararaman et al., 2016; Jacobs G.S. et al., 2019).
Archaeological materials
The Paleolithic cultural sequence established in Denisova Cave is the longest among those identified in North and Central Asia. Studies inside the cave were performed in four areas: the Main, East and South Chambers, and the entrance zone (see Fig. 2). The most complete picture of the development of the Paleolithic traditions is given in the deposits of the Middle and Upper Pleistocene in the Main and East Chambers (see Fig. 3). On the basis of the chronostratigraphic data and the techno-typological features of the lithic industries, the archaeological artifacts recovered from layers 22– 20 in the Main Chamber and layers 15 and 14 in the East Chamber belong to the Early Middle Paleolithic; from layers 19–12 in the Main Chamber and layers 13– 11.3 in the East Chamber to the Middle Paleolithic; from layer 11.2 in the East Chamber to the Initial Upper Paleolithic; from layer 11 in the Main Chamber, layer 11.1 in the East Chamber, and layer 11 in the South Chamber to the Early Upper Paleolithic; and from layer 9 in the Main, East, and South Chambers probably to the Middle Upper Paleolithic.
The earliest artifacts were recovered from layer 22 in the Main Chamber; the upper border of the corresponding deposits was OSL-dated to 287 ± ± 41 ka BP (Jacobs Z. et al., 2019). The overlying culture-bearing horizons 21 and 20 are dated to 250 ± ± 44 to 170 ± 19 ka BP. The East Chamber deposits corresponding to the above were determined as layers 15 and 14 with the age of 203 ± 14 to 187 ± 14 ka BP.
The geochronology of the basal cave deposits makes it possible to attribute the archaeological materials from these layers to the Early Middle Paleolithic. These industries are characterized by unifacial or bifacial radial and Levallois flake cores (Fig. 5, 1 , 6 , 8 ); and parallel single-platform cores (Fig. 5, 2–5 , 7 ), prepared mainly on large flakes, massive in a cross-section, and small boulders. The category of blanks is dominated by the shortened flakes with plain or natural platforms and longitudinal unidirectional or orthogonal dorsal scar patterns. Rare blades show planar striking-platforms and longitudinal dorsal scar patterns.
The predominant tools are the typologically distinct series of artifacts made on standard blanks: single straight (Fig. 6, 13 ), diagonal (Fig. 6, 12 ), transverse (Fig. 6, 3 , 5 ), convergent (Fig. 6, 1 , 7 , 8 ), and angle side-scrapers, including those fashioned by stepped retouch of Quina type; as well as spur-like (Fig. 6, 2 ), denticulate and notched tools. Numerous artifacts with ventral thinning are noteworthy; these tools were mostly fashioned on thick flakes with extensive preparation of the distal edge, and one or two lateral edges, by large ventral removals (Fig. 6, 11 ). Other typical implements are flakes with truncated basal parts, and truncated faceted flakes, most often truncated from the ventral face, more rarely from the dorsal face. The tools include naturally-backed knives, burins (a transverse one on a blade, and an angle one on a large flake (Fig. 6, 4 )), an elongated Levallois point with a faceted chapeau de gendarme platform (Fig. 6, 10 ), and blade-like flakes with discontinuous retouch (Fig. 6, 6 , 9 ).
The next stage in the Middle Paleolithic development in Denisova Cave is associated with the deposits of the Final Middle and the first half of the Upper Pleistocene. This period is illustrated by the lithic industries from layers 19–12 in the Main Chamber and layers 13– 11.3 in the East Chamber, dated to the period from 156 ± 15 to 58 ± 6 ka BP.
In the Middle Paleolithic industries, primary lithic reduction was carried out by radial (Fig. 7, 6 ) and parallel (Fig. 7, 2 , 4 , 8 ) flaking, including flaking from narrow-faced (Fig. 7, 1 , 5 ) and volumetric sub-
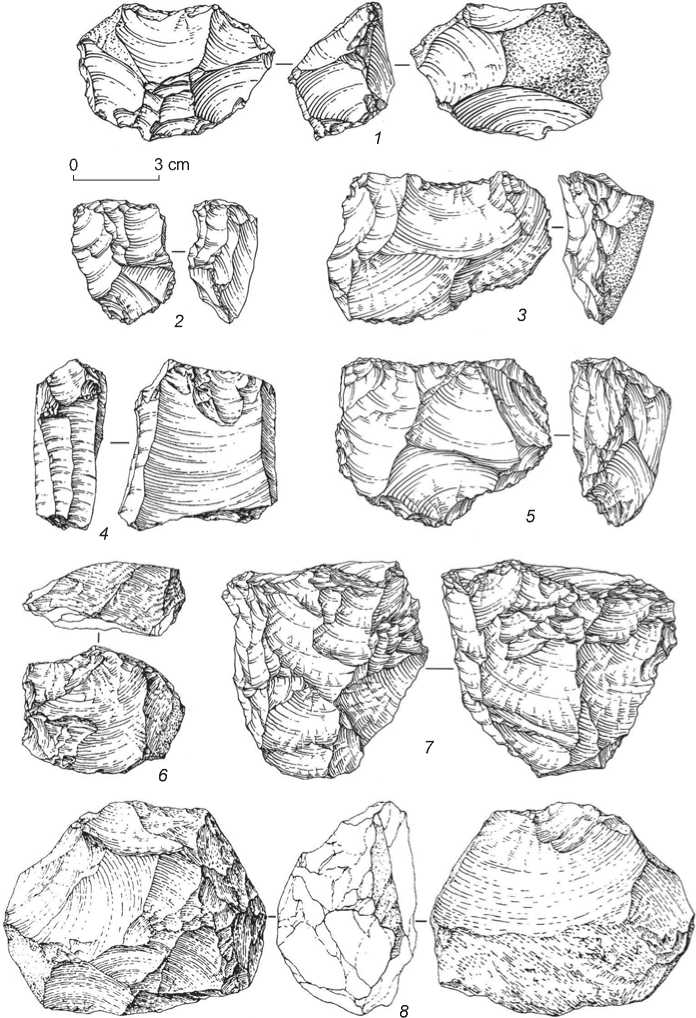
Fig. 5. Cores of the Early Middle Paleolithic from Denisova Cave.
prismatic (Fig. 7, 7 ) cores, prepared on large pebbles and boulders, more rarely on massive spalls. The Levallois reduction technique is well represented in cores intended for producing flakes (Fig. 7, 3 ) and blades, although it had no noticeable influence on the appearance of the industries. Tools were fashioned mainly on large and short flakes with a smooth or cortex residual striking-platform and longitudinal unidirectional dorsal scar pattern. The blade-like flakes are mostly medium-sized and small, more rarely large;
the proportion of blades among the blanks increases upwards in the profile. Their residual striking-platforms are mostly smooth; although there are some blades with dihedral and faceted platforms. All the Denisova Middle Paleolithic industries contain a small number of Levallois spalls: flakes, blades, and points with thoroughly faceted chapeau de gendarme platforms (Fig. 8, 16–25 ). The Levallois spalls were used mostly without any secondary working, although some blades show signs of regular retouch.
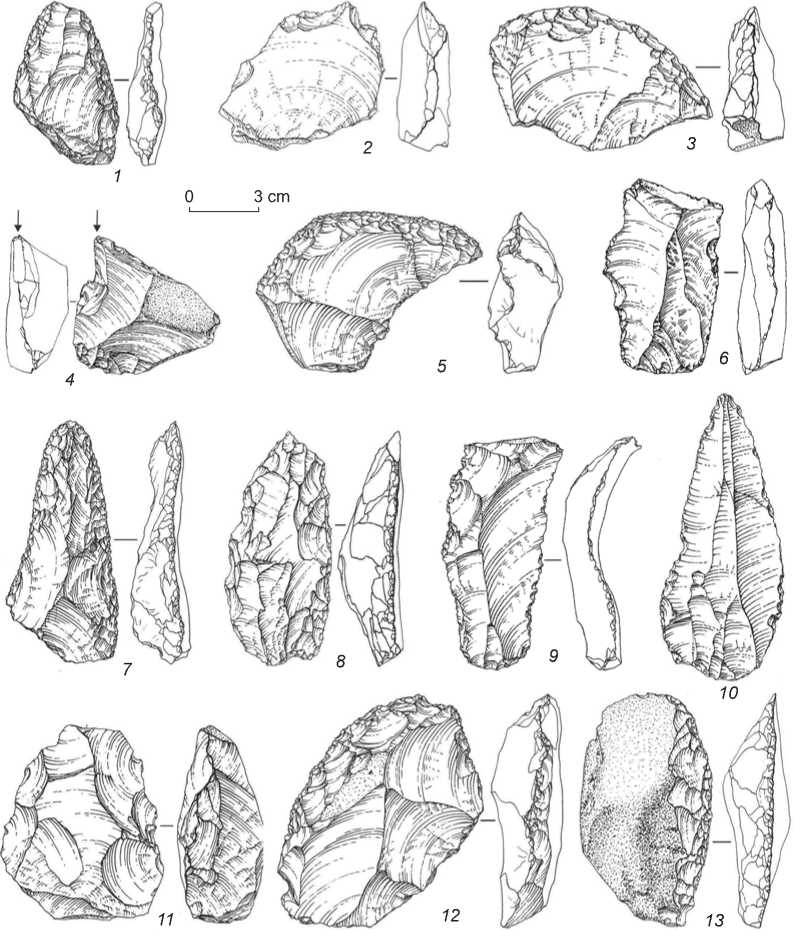
Fig. 6. Stone tools of the Early Middle Paleolithic from Denisova Cave.
1, 3, 5, 7, 8, 12, 13 – side-scrapers; 2 – spur-like tool; 4 – burin; 6, 9 – blade-like flakes with retouch; 10 – Levallois point;
11 – ventrally-thinned flake.
Side-scrapers are the most typical tools; these are dominated by single or double straight (Fig. 8, 6 , 7 ), transverse (Fig. 8, 1 ), diagonal (Fig. 8, 5 , 8 ), convergent (Fig. 8, 15 ), and angle forms. Denticulate, notched, and beak-shaped tools, fashioned mostly by retouch, more rarely by Clactonian encoches, are also numerous. There are comparatively small but typologically distinct series of the Mousterian pointed tools (Fig. 8, 10–14 ), Levallois points (Fig. 8, 18 , 19 , 21 , 22 ), and Levallois blades (Fig. 8, 16 , 20 , 23–25 ), flakes with ventral thinning, and the Upper Paleolithic tools: end- and angle-scrapers, angle burins (Fig. 8,
2–4 ), borers and truncated spalls (Fig. 8, 9 , 17 ), as well as bifacial tools.
The Initial Upper Paleolithic at Denisova Cave is represented by the materials from lithological layer 11.2 in the East Chamber. The OSL-age of these deposits is 63 ± 6 to 55 ± 6 ka BP; a series of AMS-dates has also been generated, among which the oldest are over 50 ka BP, the majority of the dates falling within the interval of 50–40 ka BP.
The archaeological materials from this layer clearly show the continuous development of technological traditions from the Middle Paleolithic to the Early
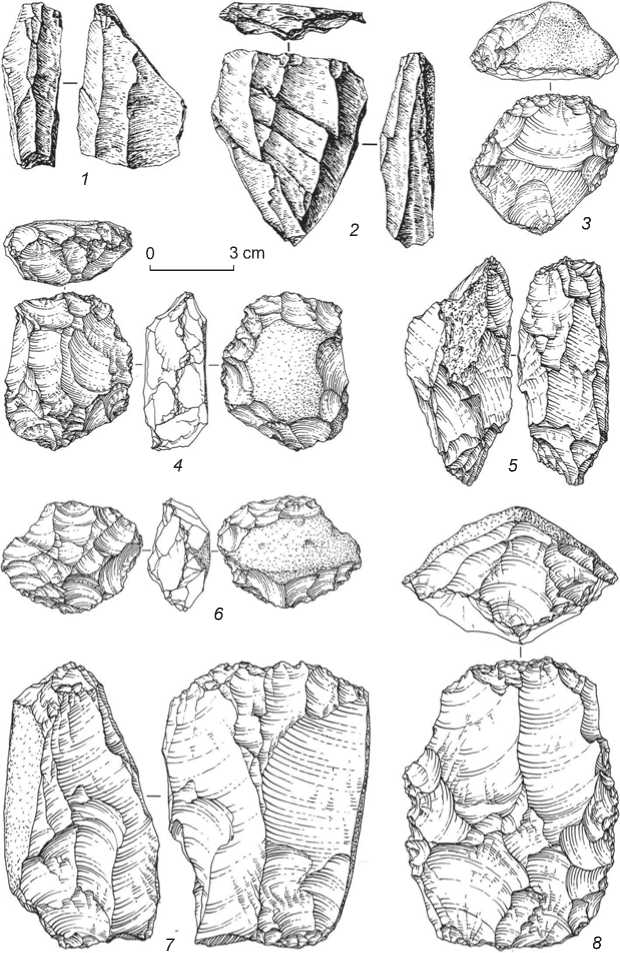
Fig. 7. Cores of the Middle Paleolithic from Denisova Cave.
Upper Paleolithic techniques of stone reduction. The major technique of stone working was parallel flaking, including from sub-prismatic (Fig. 9, 18–20 ) and narrowfaced (Fig. 9, 15 ) cores, often intended for producing large blades. The radial and Levallois reduction strategies were less common. The debitage category is dominated by the flakes with the parallel dorsal scar pattern and smooth residual striking-platform; although there are also series of elongated spalls removed by direct percussion with a hammer-stone.
In the toolkit, Middle Paleolithic side-scrapers are most numerous; these are mainly straight (Fig. 9, 14), diagonal (Fig. 9, 13), and transverse (Fig. 9, 16). The tool collection also contains a comparatively small series of typologically distinct elongated Levallois spalls, mostly pointed (Fig. 9, 11). The proportion of the denticulate, notched, and beak-shaped tools is also considerable. Another noteworthy category includes the Upper Paleolithic tool types: elongated points (Fig. 9, 10), endscrapers (Fig. 9, 1, 2, 17), and angle burins (Fig. 9, 8), some of which are fashioned on elongated spalls (Fig. 9, 7); borers, chisel-like tools (Fig. 9, 12), large truncated and truncated-faceted spalls (Fig. 9, 9), and prismatic blades with retouched long edges (Fig. 9, 3–6).
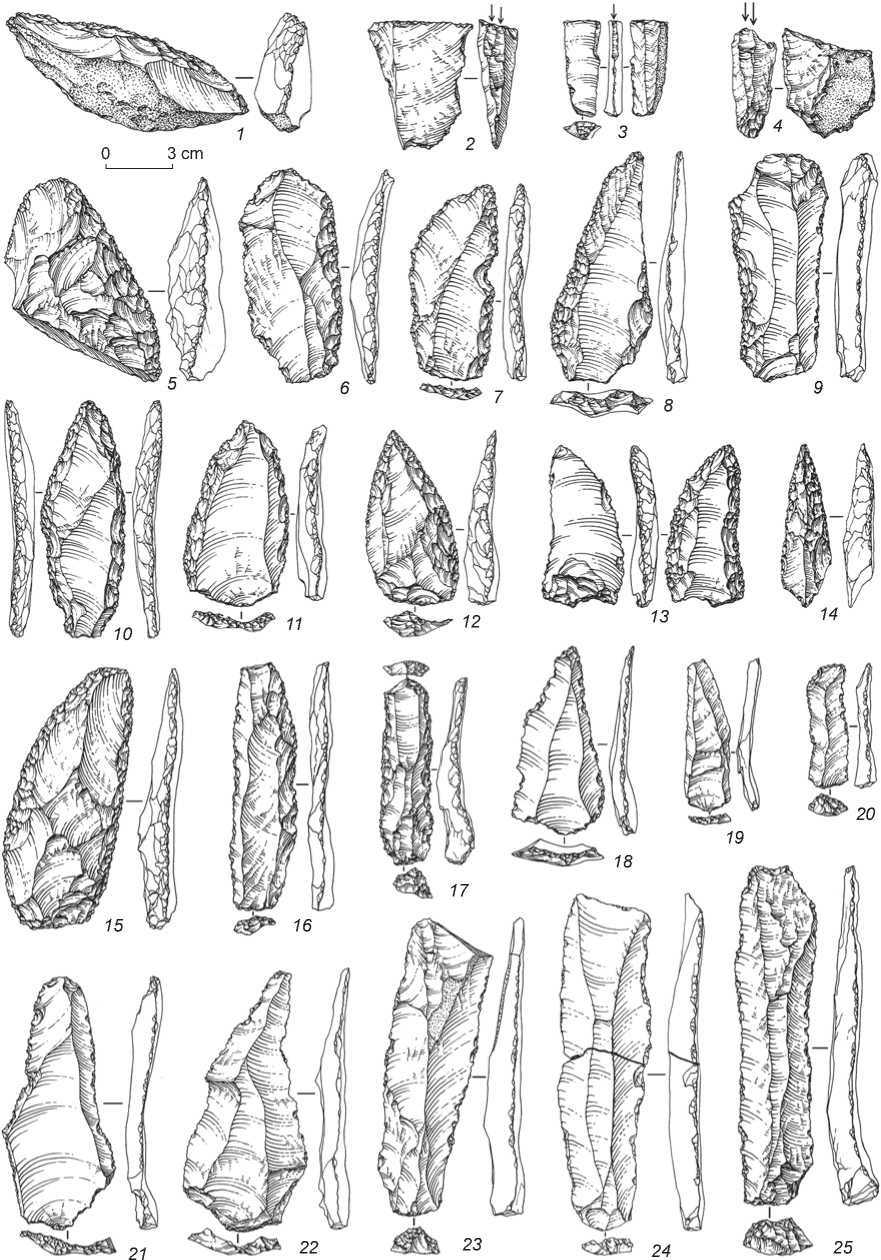
Fig. 8. Stone tools of the Middle Paleolithic from Denisova Cave.
1, 5–8, 15 – side-scrapers; 2–4 – burins; 9, 17 – truncated blades with retouch; 10–14 – Mousterian points; 16, 20, 23–25 – blades;
18, 19, 21, 22 – Levallois points.
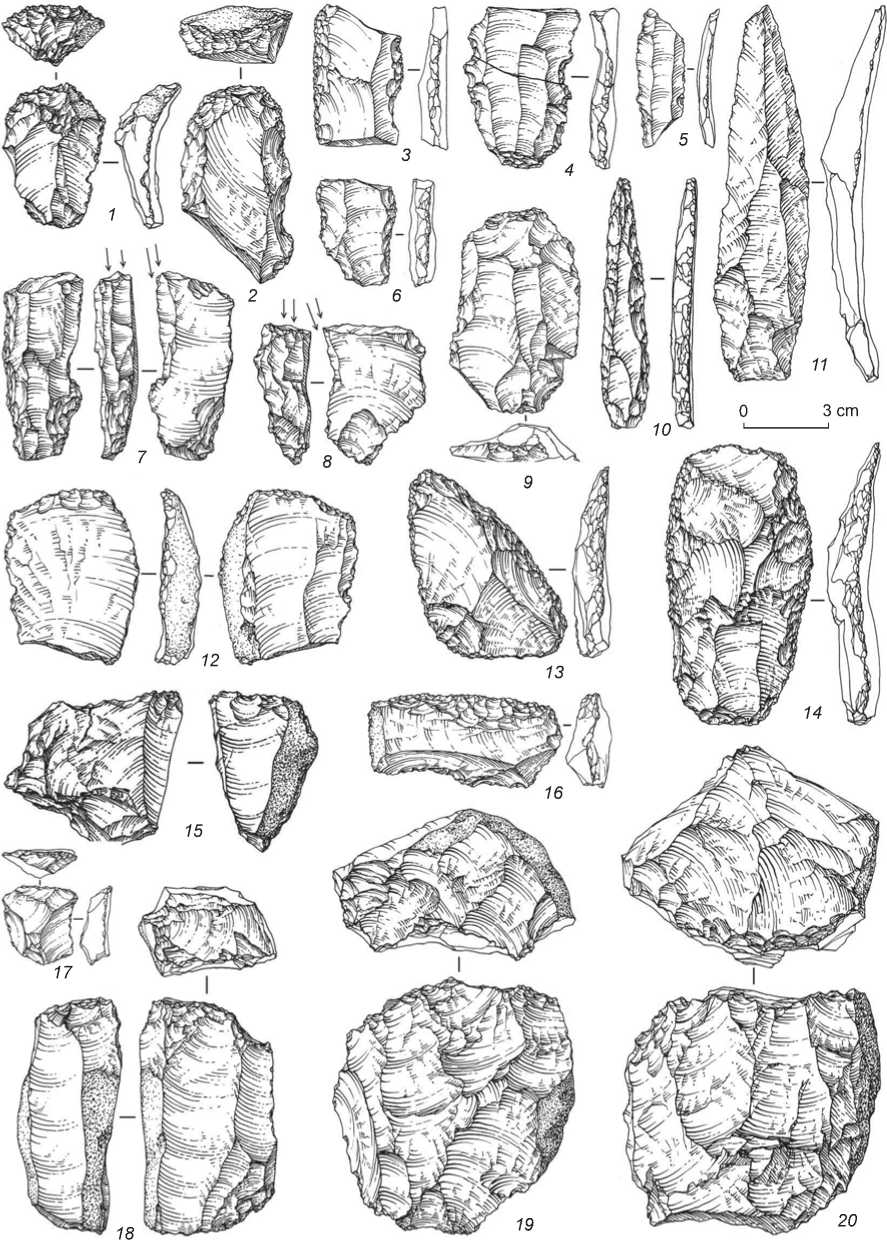
Fig. 9. Stone tools of the Initial Upper Paleolithic from the East Chamber of Denisova Cave.
1, 2, 17 – end-srapers; 3–6 – retouched blades; 7, 8 – burins; 9 – truncated-faceted spall; 10 – retouched point; 11 – pointed blade;
12 – chisel-like tool; 13, 14, 16 – side-scrapers; 15, 18–20 – cores.
Layer 11.2 also yielded bone tools and personal ornaments made of mammoth tusk, animal teeth and bones, as well as of semi-precious stone and eggshell. The bone toolkit includes retouchers, unshaped chisellike tools, and a fragment of a thickened needle with a broken eye (Fig. 10, 16 ). The ornaments include long beads or stems with engravings made of hollow tubular bones bearing symmetrical rows of deep circular grooves (Fig. 10, 9 , 10 ); pendants of maral fangs (Fig. 10, 12–14 ) and elk incisors (Fig. 10, 17 ) with biconical drilled holes; mammoth tusk blades consisting of two crescent fragments with biconical holes (Fig. 10, 15 ); a mammoth tusk ring (Fig. 10, 11 ); beads of bone (Fig. 10, 4 ) and ivory (Fig. 10, 5–8 ) ovoid and sub-rectangular in shape, with one or three

holes; a ring of white marble (Fig. 10, 21 ); serpentine pendants, each with a hole drilled out at one of the transverse sides (Fig. 10, 19 , 20 ); and flat ring-shaped beads made of fossilized ostrich eggshell—a unique material for the Altai Paleolithic (Fig. 10, 1–3 ). Among the ornaments, an elk-tooth pendant is noteworthy (Fig. 10, 18 ); its direct date is cal 42,450–49,710 years (OxA-30963), suggesting that this find is the oldest artifact of such a type in Northern Eurasia.
The Early Upper Paleolithic in Denisova Cave is represented by the materials from lithological layer 11.1 in the East Chamber, layer 11 (stratigraphic horizons 11.5–11.1) in the Main Chamber, and layer 11 in the South Chamber. The age of these deposits falls within the chronological range of 50–
30 ka BP, according to the set of OSL- and AMS-dates.
The main reduction technique of that period was parallel flaking. Radial and Levallois flaking techniques were also used. Among cores, noteworthy are single-and double-platform prismatic forms with well-prepared overhangs, from which elongated spalls were detached (Fig. 11, 22 ); this category also includes small coneshaped cores, intended for the detachment of small blades (Fig. 11, 25 , 26 ). The debitage set shows elongated spalls (Fig. 11, 12 , 13 , 18 ), including large long blades with punctiform or linear residual striking-platforms (Fig. 11, 19 , 23 , 24 ), produced using soft hammer; blades with the Levallois morphology (Fig. 11, 20 , 21 ); and also bladelets (Fig. 11, 1–4 , 6 ) and micro-blades (Fig. 11, 5 ).
The toolkit includes series of sidescrapers, mostly of straight varieties (Fig. 11, 27 , 28 ), and notched-denticulate tools. The Upper Paleolithic tool forms are represented by various end-scrapers (Fig. 11, 7–11 ), angle burins (Fig. 11, 16 , 17 ); miniature borers on blades (Fig. 11, 1 ), large prismatic blades with
Fig. 10. Bone needle ( 16 ), and ornaments of the Initial Upper Paleolithic, made of egg shell ( 1–3 ), bone ( 4, 9, 10 ), mammoth tusk ( 5–8, 11, 15 ), animal teeth ( 12–14, 17, 18 ) and stone ( 19–21 ) from the East Chamber of Denisova Cave.
1–8 – beads; 9, 10 – long beads; 11, 21 – rings; 12–14, 17–20 – pendants; 15 – blade with hole; 16 – eyed needle.
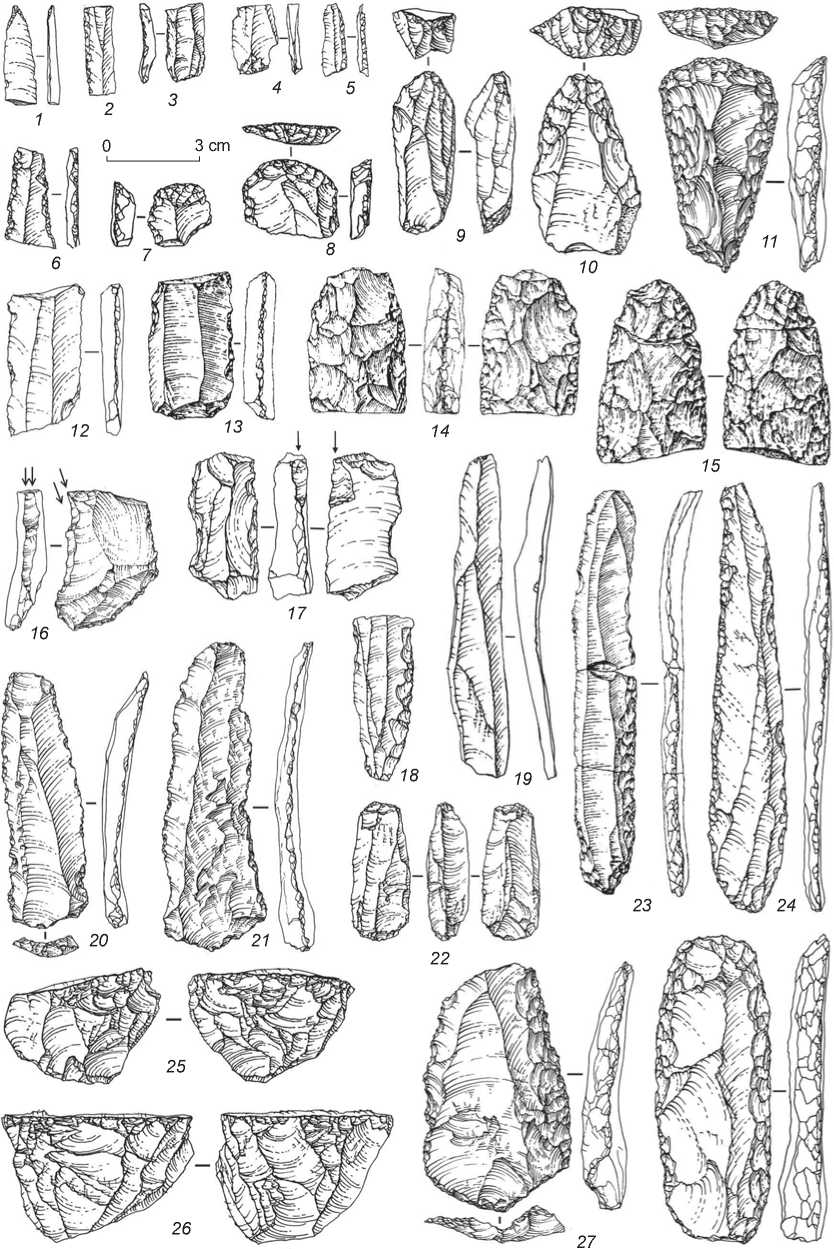
3 cm
Fig. 11. Stone tools of the Early Upper Paleolithic from Denisova Cave.
1 – borer; 2, 3, 6 – retouched bladelets; 4 – bladelet; 5 – backed micro-blade; 7–11 – end-scrapers; 12, 19–21 – blades; 13 , 18, 23, 24 – retouched blades; 14, 15 – bifaces; 16, 17 – burins; 22, 25, 26 – cores; 27, 28 – side-scrapers.
abrupt modifying retouch on the long margins (Fig. 11, 23 , 24 ), retouched blades (Fig. 11, 13 , 18 ) and bladelets (Fig. 11, 2 , 3 , 6 ), and backed micro-blades (Fig. 11, 5 ). The toolkit also includes comparatively small series of the Levallois blades with use-wear signs (Fig. 11, 20 , 21 ), and foliate bifaces (Fig. 11, 14 , 15 ).
The Early Upper Paleolithic collection from Denisova Cave contains bone tools and personal ornaments of bone, mammoth tusk, and animal teeth. The tool group comprises bone eyed needles (Fig. 12, 2 , 3 , 5 ), including an intact one 75 mm long (Fig. 12, 1 ) and a flattened ivory one with a broken tip, bearing lines of incised dots on both surfaces (Fig. 12, 4 ); points made of mammoth tusk (Fig. 12, 7 ) and of fractured long bones of large mammals (Fig. 12, 8 , 9 ), and a bone pointed tool (Fig. 12, 6 ) from lithological layer 11.4 in the Main Chamber, whose direct AMS-date is cal 42,660–48,100 ka BP (OxA-30271) and which is the oldest such tool known in Northern Eurasia.
The ornaments include pendants with biconical drilled holes or deep concentric fixing incisions encircling the root, made from the fangs of fox


Fig. 12. Tools made from bone ( 1–3, 5, 6, 8, 9 ) and mammoth tusk ( 4, 7 ) of the Early Upper Paleolithic from Denisova Cave. 1–5 – eyed needles; 6–9 – awls-borers.
(Fig. 13, 11–13 ), weasel (Fig. 13, 10 ), or maral (Fig. 13, 16, 17 ), from the incisors of elk (Fig. 13, 14 ), bison (Fig. 13, 18 ), hyena, or bear (Fig. 13, 15 ), Siberian ibex (Fig. 13, 19 ), or the sesamoid bone of horse (Fig. 13, 20 ); hollow cylinder beads or stems made from tubular bones of mammals (Fig. 13, 2 , 8 ) or large birds (Fig. 13, 3 –7 ), ornamented with deep circular incisions; a part of a bone thin-walled cylindrical implement, consisting of two fragments, ornamented with deep parallel rows of transverse incisions (Fig. 13, 1 ); three fragments of a long bone blade with short incisions grouped into local rows along the edges (Fig. 13, 21 ); small flat beads made of broken tubular bones (Fig. 13, 9 ); a thin-walled ring representing a transversely cut piece of a tubular bone belonging to a large bird; and fragmented artifacts with parallel or fanshaped incisions, made from ribs of ungulates.
The ivory collection contains diadems or pectoral plates, each represented by a medial part of a rectangular straight plate bearing lengthwise lines of triangular dimples on both surfaces (Fig. 14, 15 ) and a marginal part of a wide plate, convex in profile, with biconical hole at the end (Fig. 14, 13 ); a bracelet consisting of two flattened convex fragments (Fig. 14, 12 ); a large and massive rectangular pendant (Fig. 14, 11 ); rings (Fig. 14, 7 ) and beads of rounded, ovoid, or sub-rectangular shape, with one (Fig. 14, 1–4 ) or two (Fig. 14, 5 , 6 , 8 ) biconical holes; and a blade consisting of three fragments, with six biconical holes cut along the long axis, which blade was probably a blank for manufacturing large beads (Fig. 14, 14 ). The unique piece is a zoomorphic figurine, probably depicting a feline predator, whose head and forelegs are missing (Fig. 14, 10 ). The figurine is ornamented on all sides with rows of four short incisions bearing residues of a red mineral pigment.
Other noteworthy components of this collection are ornaments of semi-precious stone, eggshell and mollusk shell. This set includes a bracelet of dark green chloritolite (Fig. 15, 16 ); various pendants of green kaolinite agalmatolite (Fig. 15, 12 ), yellowish-brown laminated chrysotile (Fig. 15, 13 ), white marble (Fig. 15, 8 ), gray and light green serpentine (Fig. 15, 10 , 11 , 14 ), or light brown talc-steatite (Fig. 15, 9 ), each with a biconical drilled hole at one of the transverse edges; beads made of marble (Fig. 15, 3 ), talc (Fig. 15, 4–6 ), serpentine, or clay slate (Fig. 15, 7 ); bead-rings of ostrich eggshells (Fig. 15, 1 , 2 ); and ornaments made of freshwater mollusk Corbicula tibetensis shells, with sawed holes at the bases (Fig. 15, 15 ).
The category of items showing the use of coloringagents includes the sub-triangular fragment of a hard deep-red mineral pigment, and a pear-shaped pebble of milky white marble bearing residues of red ocher. Microscopic analysis of the ornaments showed traces of use of ocher pigments in the holes and microrelief unevenness of the pendants made of chrysotile and animal teeth, and eggshell beads.
Use-wear analysis and technological data on the bone and stone ornaments have shown that these were manufactured using such operations as planing, cutting, biconical drilling, boring, grinding, and polishing.
The latest stage in the cultural sequence of the Denisova Pleistocene deposits is illustrated by the Upper Paleolithic materials from lithological layer 9 in the Main, East,
Fig. 13. Ornaments made from bone ( 1–9, 20, 21 ) and animals’ teeth ( 10–19 ) of the Early Upper Paleolithic from Denisova Cave.
1 – fragment of an implement with engraving; 2–8 – long beads; 9 – a bead; 10–20 – pendants; 21 – ornamented blade.


Fig. 14. Ornaments made from mammoth tusk of the Early Upper Paleolithic from Denisova Cave. 1–6, 8, 9 – beads; 7 – ring; 10 – zoomorphic figurine; 11 – pendant; 12 – bracelet; 13, 15 – diadems; 14 – blade with holes.

Fig. 15. Ornaments made of eggshell ( 1, 2 ), semi-precious stone ( 3–14, 16 ), and mollusk shell ( 15 ) of the Early Upper Paleolithic from Denisova Cave. 1–7 – beads; 8–15 – pendants; 16 – bracelet.
angle and transverse burins, borers, chisel-like tools, atypical Levallois points, and notched and beak-shaped tools are comparatively few.
The Upper Paleolithic collection comprises a small series of tools and ornaments of bone and mammoth tusk: eyed needles (Fig. 16, 1 ), awl-borers, ivory pendants with biconical hole (Fig. 16, 3 ), and fragments of blades with artificial holes (Fig. 16, 4 ) and rows of symmetrical transverse incisions (Fig. 16, 2 ).
Despite the incompleteness in the lithologo-stratigraphic sequence, resulting from the long sedimentation gaps during the Pleistocene, the materials of the stratified Denisova complex generally illustrate the development of ancient technologies throughout the second half of the Middle and almost the whole Upper Pleistocene, spanning the period from the Early Middle Paleolithic to the Middle Upper Paleolithic, and attest to the continuity of the technological traditions of the Middle and Upper Paleolithic. The lithic industry of the Initial Upper Paleolithic was formed on the basis of the local Middle Paleolithic tradition. The similarity between the Middle and Upper Paleolithic industries was reflected by the use of one and the same raw material—pebbles of sedimentary and volcanic rocks from the alluvium of the Anui and South Chambers. According to the OSL-dates, the age of these deposits in the Main Chamber is in the range of 36 ± 4 to 21 ± 8 ka BP, which corresponds to the Middle Upper Paleolithic.
The comparatively small collection of artifacts recovered from this layer attests to the further development of blade flaking techniques based on parallel flaking of the sub-prismatic and narrow-faced cores. As compared to the previous developmental stage, the proportion of elongated blanks among the spalls increased considerably (Fig. 16, 5–7 , 11–14 , 17–19 ); the proportion of micro-blades also grew (Fig. 16, 8–10 ). The majority of blade-like flakes have residual striking-platforms with the signs of direct reduction of overhang. This lithic industry is characterized by a broad use of blades in the manufacture of tools, mostly as blanks for straight side-scrapers (Fig. 16, 13 , 18 ). In the toolkit, of special interest are retouched blades (Fig. 16, 14 , 17 ), backed bladelets (Fig. 16, 5–7 , 11 , 12 ) and micro-blades (Fig. 16, 9 , 10 ), carinated endscrapers (Fig. 16, 15 ), and a fragment of foliate biface (Fig. 16, 16 ). Such tool types as end- and side-scrapers,
River, and by the permanent application of the parallel reduction strategy. Parallel reduction of the flat cores was widely used throughout the Middle Paleolithic alongside Levallois and radial reduction. During the Initial Upper Paleolithic, parallel flaking of prismatic and narrow-faced cores came into regular use, aimed at producing series of small and large blades. Typologically, the lithic industries of the Final Middle Paleolithic are observed to be associated with the Initial Upper Paleolithic. The Middle Paleolithic industries contain artifacts with the distinct Upper Paleolithic typology: end- and angle-scrapers, angle burins, angle borers, large retouched blades, and truncated spalls. Few, but diagnostic, Levallois implements can also be regarded as an interlink between the Early Upper Paleolithic collections and the previous industries.
Emergence of signs of micro-flaking, backed micro-blades, eyed bone needles, sets of personal ornaments and other traces of symbolic behavior in the Denisovan cultural sequence as early as about 50 ka BP indicates the formation in the Altai of one of the earliest Upper Paleolithic industries in Eurasia, which was created, according to the available
Fig. 16. Bone needle ( 1 ), ornaments made of bone ( 2 ) and ivory ( 3 , 4 ), stone tools ( 5–19 ) of the Middle Upper Paleolithic from Denisova Cave.
1 – eyed needle; 2 – blade with engraving; 3 – pendant; 4 – blade with a hole; 5–7, 9–12 – backed bladelets and micro-blades; 8 – microblade; 13, 18 – side-scrapers; 14, 17 – retouched blades; 15 – end-scraper; 16 – biface; 19 – blade.
anthropological and paleogenetic data, by Denisovans. In terms of behavior, Denisovans were close to those of anatomically modern humans, who arrived in Western Siberia not later than 45 ka BP (Fu et al., 2014). However, no fossils of the anatomically modern humans were found in Denisova Cave nor elsewhere in the Altai Paleolithic sites. This fact makes us believe that the Upper Paleolithic development in the Altai was associated with the Denisovan culture.
According to the anthropological and paleogenetic evidence from layer 22.1 in the Main Chamber and layer 15 in the East Chamber (Sawyer et al., 2015; Slon et al., 2017b), Denisovans populated the cave at the very beginning of its habitation, in the Early Middle Paleolithic. The evidence of Denisovan presence has been recorded in the stratigraphic sequence up the profile: in the East Chamber, at the border of the Middle Paleolithic layers 12.1 and 11.4, and in layer 11.2, bearing the Initial Upper Paleolithic industry; in the South Chamber, in layer 11, containing the artifacts of the Early Upper Paleolithic (Reich et al., 2010; Sawyer et al., 2015). The anthropological evidence for the long-term habitation of Denisovans in the cave agrees with the cultural continuity in the development of lithic industries, and suggests that Denisovans were the autochthonous population developing the Middle and Early Upper Paleolithic cultural traditions. At the same time, the Middle Paleolithic layers in Denisova Cave yielded remains of Neanderthals (Mednikova, 2011a, 2013; Prüfer et al., 2014; Slon et al., 2017a) and a bone from the hybrid girl with a Neanderthal mother and a Denisovan father (Slon et al., 2018). These materials, together with the absence of any drastic changes in the composition of the lithic industries, make it possible to

hypothesize on some forms of cohabitation, rather than alternate habitation, of the cave by Denisovans and Neanderthals; the role of the latter in the development of the Middle Paleolithic technocomplex still remains unclear.
About 60 ka BP, a new Neanderthal population, genetically different from that in Denisova Cave, arrived in the Altai (Slon et al., 2018; Mafessoni et al., 2020). Numerous remains of this people were found in the Okladnikov and Chagyrskaya Caves, located 100 and 120 km from Denisova Cave, respectively (Krause et al., 2007; Mednikova, 2011b; Buzhilova, 2013). The late Neanderthal Paleolithic traditions with Mousterian-like industries, characterized by the radial reduction technique, and the Micoquien tools of déjeté and bifacial forms, sharply differ from the traditions recorded in Denisova Cave (Derevianko, Markin, Shunkov, 2013; Kolobova et al., 2020). According to the chronostratigraphic data of the cave deposits, the upper chronological boundary of the Neanderthal habitation in Chagyrskaya Cave corresponds to 49 ka BP, and in Okladnikov Cave to at least 44 ka BP (Kolobova et al., 2020). Hence, in the range of 50–45 ka BP, the northwestern Altai was simultaneously inhabited by the creators of Mousterian and Early Upper Paleolithic traditions—Neanderthals and Denisovans. Gradually, the ecdemic Neanderthal population was likely assimilated by the indigenous Upper Paleolithic people.
Discussion
Denisovan origins and their role in the origin of modern humans
Available facts demonstrate that in the Late Middle to Early Upper Pleistocene, the long evolution of the genus Homo in Africa and Eurasia resulted in the emergence of three taxa—early modern humans in Africa, Neanderthals in Europe, and Denisovans in Asia. These taxa were open genetic systems, and reproductive isolation between them was incomplete, as evidenced by hybridization (Krause et al., 2010;
Reich et al., 2011; Skoglund, Jakobsson, 2011; Meyer et al., 2012; Prüfer et al., 2014; Fu et al., 2015). Therefore, S. Pääbo merged the three taxa into a single metapopulation (Pääbo, 2015). One example of admixture is the Denisova 11 girl, hybrid of a Neanderthal mother and a Denisovan father (Slon et al., 2018). On their journey from Africa 80–40 ka BP, early anatomically modern humans encountered Neanderthals and Denisovans, and mixed with them, receiving genes strengthening the immune system and enhancing the adaptation to new environments.
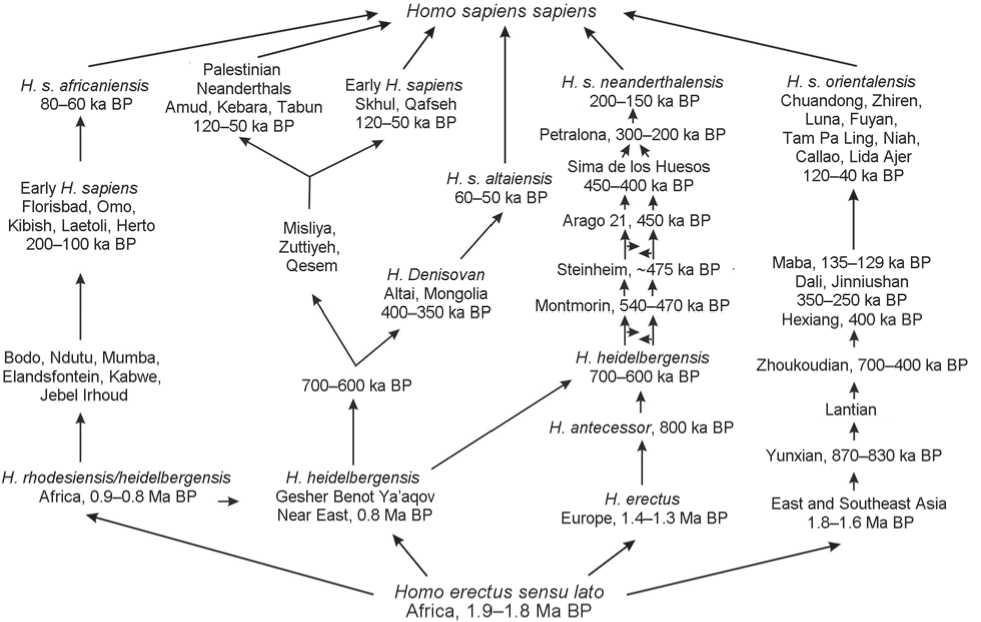
Archaic introgression demonstrates that H. erectus , having originated in Africa, migrated outside it 1.8 Ma BP or so. It was a polymorphic species, which accounts for its ability to colonize large parts of Eurasia (Fig. 17). Most hominin species living in Africa, Europe, and Asia between 1.8–0.2 Ma BP were descendants of a single biological species, who intermixed and whose hybrid progeny were fertile (Derevianko, 2019).
The evolutionary history of H. erectus includes three stages. The first stage, dating to 800– 900 thousand years, was the time when H. erectus gave rise to a new species, variously termed H. heidelbergensis, H. rhodesiensis , or archaic H. sapiens (Rightmire, 1996, 1998; Hublin, 2001,
Homo sapiens sapiens
Homo erectus sensu iato Africa, 19-18 Ma BP
H. s. orientalensis Chuandong, Zhiren, Luna, Fuyan, Tam Pa Ling, Niah, Callao, Lida Ajer 120-40 ka BP
H. erectus
Europe, 1.4-1.3 Ma BP
Maba, 135-129 ka BP Dali, Jinniushan 350-250 ka BP
Hexiang, 400 ka BP f
Zhoukoudian, 700-400 ka BP
Lantian
+
Yunxian, 870-830 ka BP
t
East and Southeast Asia
1.8-1.6 Ma BP
H. rhodesiensis/heidelbergensis Africa, 0.9-0.8 Ma BP
H. heidelbergensis Gesher Benot Ya’aqov Near East, 0.8 Ma BP
H. s. africaniensis 80-60 ka BP
Early H. sapiens Florisbad, Omo, Kibish, Laetoli, Herto 200-100 ka BP
700-600 ka BP
H antecessor, 800 ka BP
Misliya, Zuttiyeh, Qesem
Early H. sapiens Skhul, Qafseh 120-50 ka BP
H. s. altaiensis 60-50 ka BP
H. Denisovan Altai, Mongolia 400-350 ka BP
H. heidelbergensis 700-600 ka BP
Palestinian Neanderthals Amud, Kebara, Tabun 120-50 ka BP
♦ N
Sima de los Huesos 450-400 ka BP
+ +
Arago 21,450 ka BP
Steinheim, ~475 ka BP ■
I 4
Montmorin, 540-470 ka BP
H. s. neanderthalensis 200-150 ka BP
Petralona, 300-200 ka BP
Bodo, Ndutu, Mumba, Elandsfontein, Kabwe, Jebel Irhoud
Fig. 17. Phylogenetic tree of the genus Homo (after (Derevianko, 2019: Fig. 254)).
2009; Bräuer, 2008, 2010, 2012). Homo heidelbergensis and Homo rhodesiensis were chronospecies with open genetic systems, in which several evolutionary lines emerged in the Middle Pleistocene. Those taxa were associated with the spread of the Acheulean industry in Eurasia. H. heidelbergensis was the ancestor of H. sapiens , Neanderthals, and Denisovans alike (Stringer, 2012), as demonstrated by its entire further evolution (Derevianko, 2019). The migration of H. heidelbergensis to Eurasia ~800 ka BP, documented by the Gesher Benot Ya’aqov site in Israel, triggered the split of a single metapopulation into evolutionary lines leading to modern humans on the one hand, and Neanderthals and Denisovans on the other (Meyer et al., 2014).
Homo rhodesiensis remained in Africa, becoming ancestral to anatomically modern humans, who emerged 200–150 ka BP, as evidenced by their remains found in eastern, northern, and southern Africa. The transition from H. rhodesiensis to H. sapiens was a prolonged and gradual anagenetic evolution, spanning the period of 800–200 ka BP (Bräuer, 2008, 2012; Mbua, Bräuer, 2012).
The origin of anatomically modern humans is a contentious matter. G. Bräuer attributes fossils from the 300–200 ka BP interval to the late transitional archaic group of H. sapiens . This group includes remains from Ileret (the KNM-ER 3884 cranium, 270 ka BP), Laetoli 18 (250 ka BP), Florisbad (260 ka BP), Jebel-Irhoud 1 and 2 (190–170 ka BP)*, and Eliye-Springs (the age of this site is uncertain). As Bräuer believes, continuity between early and late archaic humans is evidenced by the Rabat fossil (250 ka BP), whereas the transition from archaic to early H. sapiens is documented by Omo 1 and 2, Herto, Singa, etc. (Bräuer, 2008, 2012; Mbua, Bräuer, 2012). G.P. Rightmire claims that after the emergence of the species H. heidelbergensis some 800 ka BP, its evolution followed two trajectories— toward Neanderthals and toward modern humans. At the end of the Middle Pleistocene, H. heidelbergensis gave rise to H. neanderthalensis and H. sapiens . In Rightmire’s view, the idea that the first anatomically modern humans originated in Africa is supported by fossils such as Florisbad, Laetoli, and Jebel Irhoud. In the beginning of the Upper Pleistocene, modern humans appeared in southern Africa (Klasies River Mouth) and in the Near East (Skhul and Qafzeh) (Rightmire, 2001, 2009).
Certain variation in both the cranial morphology and the lithic industries of early modern African humans suggests that they evolved from H. rhodesiensis across large areas of northern, eastern, and southern Africa, rather than in a single center.
Finds from the Gesher Benot Ya’aqov site in Israel attest to a migration of H. heidelbergensis associated with the Acheulean industry from Africa to the Near East ca 800 ka BP. In the Near East, those humans probably encountered native hominins—descendants of late H. erectus , likewise associated with the Acheulean industry, who were the first migrants from Africa (this migration is documented by the Ubeidiya site in Israel).
All Middle Pleistocene fossils found in the Levant demonstrate a mosaic combination of morphological traits typical of modern humans and Neanderthals and possibly testifying to hybridization between H. heidelbergensis and late H. erectus . As a result of these procceses, 800–200 ka BP, there emerged anatomically modern humans such as Skhul and Qafzeh, and Palestinian Neanderthals such as Tabun, Amud, and Kebara (Derevianko, 2016, 2019). Various ideas were expressed about the evolution and relationships of anatomically modern humans and Palestinian Neanderthals. B. Arensburg and A. Belfer-Cohen, based on the studies of Middle Paleolithic fossils from Israel, concluded that the so-called Neanderthals lacked Neanderthal specializations, whereas those traditionally described as anatomically modern humans displayed certain Neanderthal traits. Both the former and the latter hominins exhibit high levels of morphological variation. The analysis of fossils suggests that “Neanderthals” and anatomically modern humans coexisted in the same territory and sometimes even in the same caves (Arensburg, Belfer-Cohen, 1998: 320). This makes it difficult to subscribe to the idea that both Levantine groups were mutually antagonistic and could completely replace one another (Shea, 2001, 2007, 2008). Numerous archaeological finds from Paleolithic sites of the Late Middle to Early Upper Pleistocene in Levant demonstrate homogeneity and continuity; stone tools show similar technological and typological characteristics, which disagrees with the idea that Neanderthals had migrated to the Near East with a different lithic industry.
In the Levant, the remains of anatomically modern humans span the period between ~130–75 ka BP, whereas the earliest remains of Neanderthals date to ~130/125 ka BP, suggesting that in the beginning of the Upper Pleistocene, two human taxa coexisted in the region, showing high variation and mosaic combinations of cranial traits. Both these taxa, however, are morphologically rather similar. The same applies to their lithic industries and to their burial rites. While no remains of anatomically modern humans dating to the 55–75 thousand years interval have been found in the Levant so far, archaeological finds relating to Paleolithic sites indicate the presence of these humans in that territory in the first half of the Upper Pleistocene. The homogeneity of Middle Paleolithic industries in the Levant attests that neither gene flow nor any appreciable cultural influence from Africa or Europe was present. The Upper Paleolithic industry in that territory appears to have been mostly autochthonous, with a possible contribution from anatomically modern humans, creators of the late variant of the Nubian industry (Derevianko, 2011, 2019), or H. sapiens represented by the fossils from the Manot Cave (Hershkovitz et al., 2015).
About 450–350 ka BP, certain H. heidelbergensis from the Levant began to migrate to the Iranian Plateau and further to Central and North Asia, up to southern Siberia (Denisova Cave). This was the second stage in the formation of anatomically modern humans, when the ancestral metapopulation of late H. heidelbergensis split into Neanderthals and Denisovans. As the sequencing of the Denisovan genomes demonstrates, this occurred 430–380 ka BP (Prüfer et al., 2014; Meyer et al., 2014). Having dispersed over North and Central Asia, late H. heidelbergensis gave rise to Denisovans, who were identified in the Altai, specifically in Denisova Cave. Exactly late H. heidelbergensis became ancestral to Neanderthals and Denisovans, who emerged in the chronological interval from 400 to 200 ka BP. The lithic industry of Denisovans in the lower cultural horizons of Denisova Cave shows technical and typological parallels with the Acheulo-Yabrudian complex of the Levant (Derevianko, 2001, 2018).
Also, the eastward migration of late H. heidelbergensis during the 450–350 ka BP interval is documented by the Acheulean appearance of certain industries in Turkmenia, Kazakhstan, and Mongolia (Derevianko, 2019). Their route lay likely north of the Himalayas and Tibet. A hominin’s mandible from Baishiya Cave, in the northeastern Tibetan Plateau, dating to at least 160 ka BP, was diagnosed as Denisovan on the basis of the paleoproteomic analysis of the dentine sample (Chen et al., 2019). While no Paleolithic finds from Baishiya are available, 300– 400 km northwest of it, at several sites in Xingjian, superficially deposited Middle and Upper Paleolithic artifacts—showing elements of Levallois and blade flaking and similar to those from Denisova—were found (Derevianko et al., 2012). These have no parallels among the Late Middle to Early Upper
Pleistocene industries of China. Yet another fact demonstrating that Denisovans were present in that region is the adaptation of Tibetans to high altitudes through introgression, apparently received from those hominins (see above) (Huerta-Sánchez et al., 2014).
Having taken an eastward migration-path from the Levant, late H. heidelbergensis hybridized with late Asian H. erectus and inherited a small share of archaic ancestry from them by adaptive introgression (Prüfer et al., 2014). This might account for the erectus-like features seen in the archaic skullcap from the Late Pleistocene site of Salkhit in northeastern Mongolia (Devièse et al., 2019). The archaeological context of that find is uncertain, as it was found during gold-mining operations, but the closest parallels to the Paleolithic industries of that region are in the Denisovan complex (Rybin, 2014).
Some 50 ka BP, an Upper Paleolithic blade industry that originated in the Altai on a local basis was apparently associated with Denisovans. Combined with skeletal and paleogenetic evidence, it suggests that this taxon, at the late stage of its evolution, was a subspecies of H. sapiens : H. s. altaiensis . Indeed, paleogenetic facts make it possible that Denisovans and anatomically modern humans were immediate descendants of one and the same biological species, and did not have enough time to develop full hybrid sterility (Derevianko, 2019).
Apart from Denisova, the autochthonous emergence of the Upper Paleolithic in the Altai is documented by several other sites with a distinct stratigraphy— Ust-Karakol, Anui-3, and Kara-Bom (Derevianko, Shunkov, 2004). These sequences clearly demonstrate the continuous development of technological traditions from the Middle to the Upper Paleolithic. This continuity suggests that no external influences were involved in the formation of the Early Upper Paleolithic traditions in that region.
An evolution of the lithic industry similar to that of Denisova is evidenced by finds from the Obi-Rakhmat grotto in western Tian Shan (Grot Obi-Rakhmat, 2004). The thick Upper Pleistocene deposits of this grotto reveal a cultural sequence reflecting autochthonous evolution from the Middle to the Upper Paleolithic in the 90–45 ka BP time span. The Obi-Rakhmat industry is characterized by features of Levallois and parallel flaking technique, predominance of tools (including points) on blades, and an early appearance of small-and micro-blade flaking in the Early Upper Paleolithic. One of the habitation layers, dating to ~60 ka BP, yielded six upper teeth and about 150 small cranial fragments from an adolescent aged 9–12. The large dimensions and archaic features of the teeth, and the morphology of the preserved cranial bones, combining modern and Neanderthal traits, do not warrant an unequivocal taxonomic attribution. However, the largest of the teeth, M3, is comparable in size only with three Upper Pleistocene third molars, namely those of the early anatomically modern human Oase 2 and of two Denisovans (Denisova 8 and Denisova 4) (Glantz et al., 2008; Bailey et al., 2008; Sawyer et al., 2015). The characteristics of the lithic industry (see above), as well as the dental and cranial features of the Obi-Rakhmat individual, favor the idea that Denisovans had taken part in the evolution of the Middle and Early Upper Paleolithic traditions in western Central Asia.
Early Upper Paleolithic industries broadly similar to that of Denisova are known at Ushbulak in eastern Kazakhstan (Anoikin et al., 2019), Moiltyn Am, Orkhon-7, Tolbor-4, and Tolbor-21 in northern Mongolia (Okladnikov, 1981, 1986; Derevianko et al., 2007; Derevianko, Kandyba, Petrin, 2010; Rybin, 2014), as well as in southern Siberia, northern China, and Korea (Derevianko, 2001, 2005, 2006), indicating an expansion of the Denisovan traditions to vast territories of North, Central, and East Asia.
About 700–600 thousand years ago, the H. heidelbergensis population with Acheulean industry left the Levant and migrated to Europe. Here, the Heidelbergians encountered late members of the H. erectus species. In the Middle Pleistocene, that territory was associated with various industries (Acheulean, pebble-flake, Clactonian, Buda, smalltool (Tayacian?)) and with various morphologies of hominins. The most informative human fossils, dating to the Final Early Pleistocene, were found at Atapuerca, Spain, specifically at Sima del Elefante Cave (level TE 9, 1.3–1.2 Ma BP) (Carbonell et al., 2008) and at Gran Dolina (level TD 6, 0.9–0.7 Ma BP) (Bermúdez de Castro, Nicolás, 1997). On the basis of hominin remains from the TD 6 level, a separate species (H. antecessor) was described. It is believed to be ancestral to H. heidelbergensis (Martinón-Torres et al., 2006, 2007; Carbonell et al., 2008; Dennell, 2009) and might thereby be a common ancestor of Neanderthals and H. sapiens (Bermúdez de Castro, Rosas, Nicolás, 1999). The morphological characteristics of H. antecessor make it possible to attribute this taxon to the late H. erectus descending from the Sima del Elefante hominins and likewise associated with the pebble-flake industry. The common ancestor of the late H. erectus and H. heidelbergensis was H. erectus and, accordingly, hybridization was still possible. Members of the H. antecessor species may have been the first hominins to have hybridized with the migrant Heidelbergians; and cultural exchange occurred as well.
For the interval between 600–200 ka BP in Europe, about ten hominin populations were identified. They were named after eponymous sites—Caune de l’Arago, Ceprano, Steinheim, Sima de los Huesos, Fontéchevade, Petralona, Bilzingsleben, Lazaret, etc. They display both similarities and differences, and their lithic industries were likewise variable— Acheulean, Clactonian, pebble-flake, etc. Despite biological and cultural variation, possibly caused by diverse environmental conditions and different subsistence strategies, all Middle Pleistocene hominin populations of Europe were open genetic systems capable of hybridization. The ultimate outcome of their gene exchange and acculturation was the emergence of European Neanderthals in the 200–150 ka BP time window. In taxonomic terms, this population, associated with the Mousterian tradition, should be termed H. s. neanderthalensis (Derevianko, 2019). In short, this was a subspecies of modern humans, hybridizing with other early members of the H. sapiens species. For instance, the proportion of the Neanderthal component in the genome of an anatomically modern human from Oase, Romania, dating to 37–42 thousand years, is about 7 % (Fu et al., 2015), whereas the corresponding proportion in the genomes of modern non-Africans is about 2 % (Prüfer et al., 2014).
Paleolithic traditions in East and Southeast Asia followed different courses from those in Africa and western Eurasia. In the former areas, modern humans evolved from the Asian H. erectus , whose ancestors were representatives of the first wave of migration. They developed their own lithic technologies, which were adapted to local ecological conditions and differed from those practiced in the more westerly parts of Eurasia.
Several physical anthropologists, noting the morphological similarity between the late H. erectus populations of Africa, Europe, and East Asia, attribute hominins from Jinniushan and Dali in China to H. heidelbergensis (Groves, 1994; Rightmire, 1996, 1998). In the last 20–30 years, more than a dozen fossils representing modern humans and dating to the 40–120 thousand years interval have been discovered in East and Southeast Asia, indicating the formation of one more subspecies, H. s. orientalensis , in those regions (Derevianko, 2011, 2019).
The third stage in the evolution of modern humans dates to 60–30 ka BP. It starts from the migration of H. s. africaniensis from Africa to Eurasia. In the
Near East, those people mixed with early modern humans and Palestinian Neanderthals; in Europe they hybridized with H. s. neanderthalensis ; in North and Central Asia, with H. s. altaiensis ; and in East and Southeast Asia, with H. s. orientalensis (Fig. 17).
Conclusions
The entire body of available archaeological, paleoanthropological, and paleogenetic facts suggests that anatomically modern populations in Africa and Eurasia evolved from the same ancestral species— H. erectus . This polytypical species apparently gave rise to all later human groups. H. rhodesiensis , H. heidelbergensis , H. antecessor , and other taxa that existed in Africa and Europe, and late H. erectus in East and Southeast Asia, were ancestral to four human subspecies: H. s. africaniensis in Africa, H. s. neanderthalensis in western Eurasia, H. s. altaiensis in North and Central Asia, and H. s. orientalensis in Southeast and East Asia. All taxa filial with regard to H. erectus , and showing both similarities and differences, retained the ability to hybridize. This conclusion is based on the hypothesis that after almost 2 mln years of evolutioin of H. erectus , three taxa were formed to the beginning of the Upper Pleistocene in Africa and Eurasia: H. s. africaniensis , H. s. neanderthalensis , and H. s. altaiensis , who were able to intermix and whose hybrid progeny was fertile. Thus, these were not different species, but subspecies, and interbreeding took place inside one species.
Obviously, the contributions of those taxa to the modern gene pool were unequal. According to paleogenetic data, modern humans first emerged in Africa by way of the evolutionary continuity H. erectus – H. rhodesiensis – H. s. africaniensis . The last-named taxon displayed the highest genetic diversity, and its contribution to the gene pool of modern humankind was maximal.
In the Near East, early modern humans of the Skhul-Qafzeh type and Palestinian Neanderthals were formed evidently by way of hybridization between H. heidelbergensis and late H. erectus populations of the first wave of migration.
In western Eurasia, about a dozen taxa of Middle Pleistocene hominins have been described, displaying various proportions of archaic and modern features, apparently due to gene exchange between late H. erectus populations associated with the pebble-flake tradition, and those of H. heidelbergensis associated with the Acheulean industry. About 200–150 thousand years ago, this intricate evolutionary process resulted in the emergence of European H. s. neanderthalensis, associated with the Mousterian-type industries. At the final stage of the Middle Paleolithic, Neanderthals showed highly efficient technological complexes and certain elements of modern human behavior. The results of recent archaeological, skeletal, and paleogenetic studies make it possible to associate late Neanderthals with transitional industries such as the Châtelperronian, Uluzzo, and Bacho-Kiro, which had apparently taken part in the emergence of the Upper Paleolithic in Western and Central Europe. Biological facts, in addition, suggest that H. s. neanderthalensis contributed to the evolution of H. sapiens sapiens, and that this contribution, estimated at ~2 %, can be traced in the gene pool of modern non-Africans (because the distribution area of Neanderthals did not extend to Africa, virtually no such trace is observed in modern African genomes).
The population of late Heidelbergians, having migrated eastwards from the Levant in the second half of the Middle Pleistocene, made contact with H. erectus descendants in South and Central Asia, resulting in the emergence of the Denisovan population, which appeared in the Altai about 300 ka BP. Since the Early Middle Paleolithic, the peculiar lithic industry of the Denisovans had been evolving under occasional influences from the Altaian Neanderthals, culminating in the emergence of the autochthonous variety of the Upper Paleolithic ca 50 ka BP. Notably, the level of the Early Paleolithic culture (material and apparently spiritual) associated with Denisovans was no lower than that shown by the culture of early H. sapiens in Africa. These facts in toto enable us to speak of a separate human subspecies, H. s. altaiensis . This label is all the more warranted because Denisovans, like Neanderthals, participated in the origin of the modern human species, having transmitted about 6 % of their genetic ancestry to modern populations in Australia, Oceania, and insular Southeast Asia.
Given the absence of external influences on the lithic industries of the Middle and Early Upper Pleistocene in East and Southeast Asia, it can be suggested that the migration waves of late H. heidelbergensis had not reached those regions. This implies the preservation and a continuous evolution of local Asian populations of H. erectus, and the autochthonous origin of one of the subspecies of modern man—H. s. orientalensis, as well as an independent model of the evolution of Paleolithic industries in the eastern part of Asia. Hominins of that territory hybridized with populations of adjacent regions, as evidenced by a small genetic introgression from Denisovans and Neanderthals in the modern gene pool of East and Southeast Asia, and by a small genetic component, which the Denisovans had evidently received from the Asian H. erectus.
The presence of Neanderthal and Denisovan ancestry in modern human genomes, then, attests to the emergence of autochthonous cultural traditions in Africa and Eurasia, underlying the independent models of transition to the Upper Paleolithic. Those processes were accompanied by the emergence of early modern populations, which, in one way or other, had taken part in the evolution of modern humankind.
Acknowledgments
We are deeply thankful to all our colleagues, whose contribution to the multidisciplinary studies at Denisova Cave was invaluable: S. Pääbo, R. Roberts, Z. Jacobs, T. Higham, K. Douka, V. Slon, S. Brown, B. Viola, A.K. Agadzhanyan, N.S. Bolikhovskaya, A.P. Buzhilova, M.B. Mednikova, V.A. Ulianov, and to all the numerous participants of projects focusing on the Paleolithic sites in the Altai.
