A genetic analysis of human remains from the Bronze age (2nd millennium bc) cemetery Bertek-56 in the Altai mountains
 cemetery Bertek-56 in the Altai mountains")
Author: Pilipenko A.S., Trapezov R.O., Cherdantsev S.V., Zhuravlev A.A., Molodin V.I.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 4 т.44, 2016.
Free access
Short address: https://sciup.org/145145282
IDR: 145145282 | DOI: 10.17746/1563-0102.2016.44.4.141-149
Text of the article A genetic analysis of human remains from the Bronze age (2nd millennium bc) cemetery Bertek-56 in the Altai mountains
Cemetery Bertek-56 at the Ukok Plateau in the Altai Republic (Fig. 1) was studied in the summer of 1992 by the Western-Siberian branch of the North Asian Joint Expedition of the Institute of Archaeology and Ethnography (Siberian Branch of RAS) led by
V.I. Molodin. The excavation was a part of international project “Pazyryk” (Molodin, 1993). The site is a stone ring, 14 m in diameter, filled with stones inside (Fig. 2, 3), under which two burials at the level of subsoil were discovered. The deceased were placed in stone “cysts” consisting of long side walls (but without end walls), covered by massive slabs (Molodin et al., 2004: 205– 206). They were buried in extended positions on their left sides, with heads in an eastward direction (Fig. 4, 5). The absence of some skeletal bones might suggest the

Fig. 1. Ukok Plateau.
secondary nature of the burials. On the basis of skeletal morphology, one of the deceased (bur. 1) was an infant (8–9 years old) and the second (bur. 2), an adult male who died at the age of 30– 35 (Chikisheva, 2012: 129–131).
Fragments of three ceramic vessels found at the site of Bertek-56 suggest that it can be reliably dated to the Middle Bronze Age (2nd millennium BC). Numerous previous studies (Molodin, 1993; Molodin et al., 2004: 206; Molodin, 2006) have demonstrated that this cemetery is atypical of the contemporaneous Karakol culture of the Altai Mountains, and thus cannot be attributed to that culture.
The cranial morphology of the deceased shows, in general, a close similarity to the Caucasoid pattern; but their cranial features and dental traits suggest a mixed ancestry for these individuals (Chikisheva, 2012: 129–131) (see also Results and Discussion).
Modern methods of paleogenetics can potentially provide more comprehensive and reliable data on ancestry and relationships between ancient populations, as well as on the sex of skeletons and kinship among individuals buried in double or communal graves. As the skeletons from Bertek-56 were well-preserved, it was

Fig. 2. Cemetery Bertek-56 after cleaning.
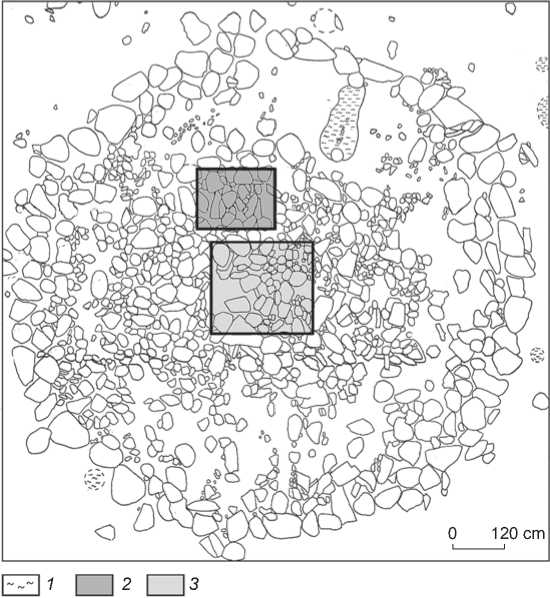
Fig. 3. Layout of the cemetery. 1 – fireplace; 2 – burial 1; 3 – burial 2.
0 20 cm
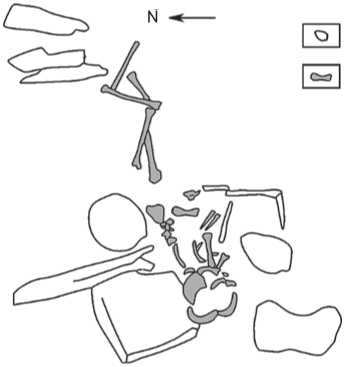
Fig. 4. Burial 1.
1 – stone construction; 2 – human remains.
possible to conduct a thorough molecular genetic investigation, including study of sex markers, uniparental markers (important from phylogenetic and phylogeographic points of view), and other markers of genetic affinity. In this article, we present the results of that investigation, and assess it in the light of previous archaeological and paleoanthropological studies of the site.
Materials and methods
Skeletal samples. The best-preserved postcranial bones (visually assessed) were sampled for the genetic analysis: the femur of the first individual (child) and the tibia of the second individual (adult). Notably, the bone tissue of the adult male was very well preserved while bone of the child exhibited low density, different color, and porosity, which indirectly pointed towards a reduced amount of ancient DNA in that specimen.
In order to exclude the possibility of crosscontamination between the specimens of individuals 1 and 2, they were treated separately at different times.
0 20 cm
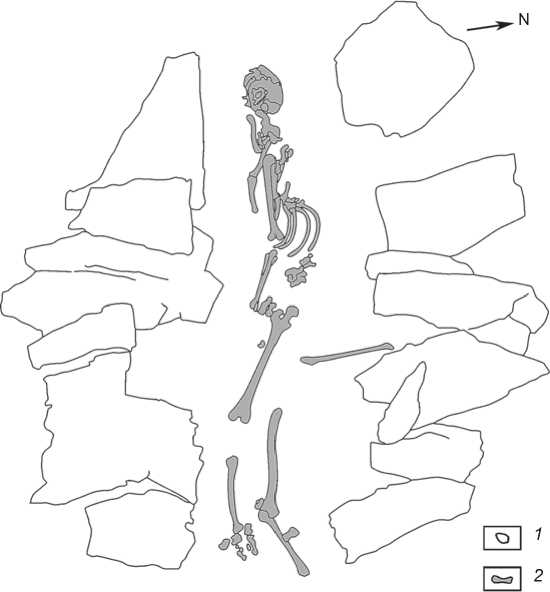
Fig. 5. Burial 2.
1 – stone construction; 2 – human remains.
Pre-treatment of the skeletal samples and DNA extraction. The protocol of this study is thoroughly described in our previous publications (Pilipenko, Romashchenko, Molodin et al., 2010; Pilipenko, Trapezov, Zhuravlev et al., 2015). In order to eliminate any modern-DNA contamination, the external surfaces of the samples were treated by a 5 % solution of bleach, and then irradiated with UV for 1 hour. After that, the external layer (1–2 mm thick) was mechanically removed, and the samples were irradiated again for not less than 1 hour. Fine bone-powder was then drilled out from the cortical layer.
Before DNA extraction, the bone powder was incubated in 5M guanidine thiocyanate buffer at 65 ºC, while being constantly mixed during incubation. DNA extraction was performed using a phenol/ chloroform protocol with subsequent sedimentation with isopropanol.
Analysis of genetic markers. This study employed four systems of genetic markers: mtDNA (HVR I region and some informative polymorphisms in the mtDNA coding region); fragment of the amelogenin gene (sex marker); hypervariable autosomal STR-loci (universal markers of kinship); STR-markers of the Y-chromosome (phylogenetically and phylogeographically important markers, male line kinship markers) (see: (Pilipenko, Trapezov, Polosmak, 2015)). Study of mtDNA and STR-loci of the Y-chromosome permits the reconstruction of population-history through both female and male lines, respectively. Genotyping protocols for both markersystems are described below.
Amplification of the mtDNA HVR I region was carried out using two different protocols: four short overlapping fragments in one round of PCR (Haak et al., 2005); and one long fragment by nested PCR (consisted of two reaction rounds) (Pilipenko et al., 2008). Amplification of informative fragments of the mtDNA coding region was performed using the primers described in (Wilde et al., 2014).
DNA sequencing was carried out with an ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). Sequencing extracts were analyzed with an ABI Prism 3100XL Genetic Analyzer automatic capillary sequencer (Applied Biosystems, USA) at the SB RAS Genomics Core Facility . The obtained results were interpreted using phylogenetic and phylogeographic analysis, as described earlier (Pilipenko, Trapezov, Polosmak, 2015).
Profiling of nine autosomal STR-loci and analysis of the amelogenin gene region polymorphism was performed using a commercial AmpFlSTR® Profiler® Plus PCR Amplification Kit (Applied Biosystems, США), following the manufacturer’s protocol. Profiles of 17 STR-markers of the Y-chromosome were determined using commercial
AmpFlSTR® Y-filer® PCR Amplification Kit (Applied Biosystems, США), following the manufacturer’s protocol. Haplogroups of the STR-haplotypes of the Y-chromosome were determined using freeware “Haplogroup predictor” .
Measures against contamination, and verifi cation of the results. All procedures with the skeletal specimens were carried out in a specially-equipped Laboratory for Molecular Paleogenetics (Institute of Cytology and Genetics of the SB RAS and Institute of Archaeology and Ethnography of the SB RAS, Novosibirsk, Russia). A description of the anti-contamination measures and verification of the results can be found in our previous publication (Ibid.). The employment of those measures and the consistency of the obtained results ensure the reliability of our experimental data.
Results and discussion
DNA preservation. Our analysis has shown different states of preservation of the ancient DNA in the two skeletal samples from Bertek-56. For both individuals, complete data on mtDNA structure were obtained, including HVR I sequence and the status of certain informative positions in the coding regions of mtDNA. We have successfully amplified both short fragments of mtDNA (up to 150 bp, using one round of PCR) and a long sequence (using two rounds of nested PCR). The efficiency of amplification of the latter was less for individual 1 (child) than for individual 2 (adult). Difference in DNA preservation between the two individuals most strongly affected analysis of the nuclear loci. For individual 2 it was possible to obtain full data for all systems of nuclear markers (polymorphic part of amelogenin gene, full allelic profiles of the autosomal and Y-chromosome STR-loci). This suggests a high degree of preservation of nuclear DNA in the adult remains. For the child (individual 2) it was only possible to amplify fragments of DNA less than 15 bp long, including the polymorphic part of the amelogenin gene, and the shortest fragments containing some autosomal STR-loci (4 out of 9) and Y-chromosome STR-loci (5 out of 17). Amplification of DNA extracted from the child’s skeleton demonstrated low efficiency for all systems of nuclear markers. This fact suggests that the reason for the low efficiency was the poor preservation of the DNA, but not the ineffectiveness of the PCR techniques employed in our study. Notably, the poor DNA-preservation in the child’s remains is associated with generally worse preservation of the bone tissue in that individual. This finding provides an example of a different degree of DNA preservation in bones of individuals buried at the same burial ground, in similar ambient conditions. Such a situation we already observed earlier, during the investigation of human remains from Pazyryk burials in the Altai Mountains (Pilipenko, Trapezov, Polosmak, 2015). In our opinion, this difference can be explained both by local environmental differences within a burial ground, and by different conditions in which the remains were kept before inhumation. The latter seems to be the case in Bertek-56. It was previously noted that the absence of some bones, despite the right anatomical position of the skeletons, could suggest that they were reburied.
Sex and kinship determination. Determination (or confirmation) of the sex of the deceased, as well as assessment of their kin relationships, are among important aims of a paleogenetic study of double and communal burials. Both individuals from Bertek-56 were identified as males. There was initially no doubt that the adult individual was male. The sex of the child was also tentatively determined as male on the basis of the skull’s size and robustness (Chikisheva, 2012: 130). Our results (presence of two alleles of amelogenin gene; amplification, albeit partial, of STR-loci) confirm this determination (Table 1).
For assessment of kinship between individuals, it is necessary to have data on mtDNA, and full (or almost full) profiles of autosomal and Y-chromosome STR-loci. However, owing to the poor preservation of the boy’s nuclear DNA (see above), only partial profiles of the STR-loci were obtained (Tables 1, 2). Nevertheless, even these incomplete data provide strong evidence that direct kinship between the two individuals can be excluded. In other words, they could not have been father and son. This case illustrates the necessity for objective assessment of kinship between individuals buried in double and communal graves instead of making “common-sense” assumptions about genetic relationships between the buried. It is of note, though, that for final rejection of direct kinship between the individuals, more complete profiles of STR-loci need to be obtained.
Phylogenetic and ph ylogeographic interpretation of mtDNA and Y-chromosome data. In both individuals buried at Bertek-56, mtDNA HVR I was sequenced (Table 3). We have also determined the status of the positions in the coding region of mtDNA that mark haplogroups of mtDNA previously determined on HVR I. The results obtained for HVR I and coding region of mtDNA in both individuals were similar. The structure of the haplotypes of HVR I suggest a definite phylogenetic position of the studied samples of mtDNA.
The mitochondrial DNA of the man possesses the 16223T-16287T-16298C-16327T haplotype, and belongs to Eastern Eurasian haplogroup C. The structure of the haplotype does not permit determination of the subgroup to which this variant belongs. The mitochondrial DNA of the boy possesses the 16145A-16224C-16311C haplotype, and belongs to the Western Eurasian
Table 1. Genotypes of the autosomal STR-loci and polymorphic part of the amelogenin gene
|
Locus |
Genotype |
|
|
Individual 1, child |
Individual 2, adult |
|
|
D3S1358 |
15/17 |
15/15 |
|
vWA |
17/17 * |
18/18 |
|
FGA |
No data |
21/23 |
|
D8S1179 |
14/14 * |
14/16 |
|
D21S11 |
No data |
30/32.2 |
|
D18S51 |
Same |
22/22 * |
|
D5S818 |
13/13 * |
10/13 |
|
D13S317 |
No data |
10/12 |
|
D7S820 |
Same |
10/12 |
|
Amelogenin (sex) |
XY (male) |
XY (male) |
*Signal from the second allele can be absent, since the allele was not amplified owing to DNA degradation.
Table 2. Y-chromosome STR-loci profile
|
Locus |
Genotype |
|
|
Individual 1, child |
Individual 2, adult |
|
|
DYS19 |
No data |
13 |
|
DYS385a/b |
Same |
15/21 |
|
DYS389I |
13 |
14 |
|
DYS389II |
No data |
30 |
|
DYS390 |
Same |
24 |
|
DYS391 |
" |
9 |
|
DYS392 |
" |
14 |
|
DYS393 |
13 |
14 |
|
DYS437 |
14 |
14 |
|
DYS438 |
No data |
12 |
|
DYS439 |
Same |
11 |
|
DYS448 |
" |
19 |
|
DYS456 |
15 |
16 |
|
DYS458 |
No data |
16 |
|
DYS635 |
Same |
22 |
|
YGATAH4 |
13 |
10 |
|
Haplogroup |
? |
Q |
Table 3. mtDNA specimens structure
|
Individual |
Age and sex (based on skeletal morphology) |
mtDNA HVR I haplotype |
mtDNA coding region markers |
mtDNA haplogroup (subgroup) |
Genetically determined sex |
|
1 |
Child, 8–9 years |
16145A - 16224C-16311C |
12308G |
K (K1a24a) |
Male |
|
2 |
Male, 30–35 years |
16223T-16287T-16298C-16327T |
13262G |
C |
" |
haplogroup K. A substitution in the 16145 position suggests that this variant belongs to the K1a24a subgroup.
These variants (as well as the haplogroups they belong to) are different from a phylogeographic point of view. Haplogroup C is widely spread in modern indigenous populations of Eastern Eurasia. It is most typical of Siberian populations, but is also found in Central Asia and Altai. The 16223T-16287T - 16298C-16327T haplotype is rare, and shows a mosaic distribution. According to previous publications, it is found in Central Asia in the Kyrgyz (Comas et al., 1998), but also in the north of Western Siberia, in the Khanty (Gubina, Osipova, Villems, 2005). Because of such a mosaic distribution, this variant of the haplogroup cannot be associated with the population of any particular region. We can only deduce that it undoubtedly belongs to the Eastern Eurasian mtDNA cluster and likely reflects a genetic impact from some local Southern Siberian population (or a population from adjacent regions). This hypothesis is also confirmed by the omnipresence of several variants of haplogroup C in the autochthonous Eastern Eurasian component of the mtDNA gene pool of ancient populations of adjacent regions of the south of Western and Eastern Siberia (spanning from the Cis-Baikal region to the forest-steppe zone of Western Eurasia), from the Neolithic on (Molodin et al., 2012; Trapezov, Pilipenko, Molodin, 2014). According to our unpublished results, haplogroup C is also present in the gene pool of the Okunev population of Minusink Basin and in the Bronze Age Karakol population of the Altai Mountains (though in this case, different variants of this haplogroup are found). Owing to the lack of data on the mtDNA gene pool of the populations from the Altai-Sayan and adjacent regions of Southern Siberia, it is impossible to find an association with ancient groups of the region known from archaeological data. The above-mentioned variant likely represents an autochthonous genetic substrate, rather than arising from genetically contrasting groups from other regions of Eurasia.
Unlike haplogroup C, haplogroup K is typical of the gene-pool of Western Eurasian populations. Interestingly, variants identical to the one observed in the boy from Bertek-56 are only found in rare cases in the populations of Southwestern and Central Europe:
the Spaniards (Larruga et al., 2001) and the French (Richards et al., 2000). Similar, but not identical, variants were found in the Basques (Corte-Real et al., 1996) and the Germans (Pfeiffer et al., 1999); and outside Europe, in the Iranians (Metspalu et al., 2004). Subgroup K1a is found mostly in haplogroups of Near Eastern origin brought to Europe by Neolithic farmers and stockbreeders. The source of the later spread of this subgroup into Eastern Eurasia could have been either Eastern Europe (and adjacent areas) or the region of the Near East/Southwest Asia. Certainly, it is impossible to choose between these two possibilities on the basis of just one specimen. We can only state that presence of haplogroup K undoubtedly reflects the westward direction of the genetic relationships of the Bertek-56 population. The Western Eurasian gene-flow might have taken place not only in the Middle Bronze Age, but in previous times as well. Another structural variant of haplogroup K was found in, among others, a representative of the Afanasievo culture from the Altai-Sayan (Trapezov, 2014). One of its variants (an unrelated subgroup) was found in a representative of the Andronovo culture from Minusinsk Basin (Keyser et al., 2009), though haplogroup K, to our knowledge, was not as typical of the gene pool of populations of Andronovo historical-cultural community.
Thus, the two variants of mtDNA found in the Bertek-56 samples are phylogenetically and phylogeographically in contrast. Their simultaneous presence in the genepool of the same ancient population points towards that population’s mixed origin. The population emerged as a result of an admixture of groups of local (or Eastern Eurasian) and Western Eurasian origins. Such a structure is typical of the Middle Bronze Age population in the vast contact zone between Western and Eastern Eurasian populations that includes Central Asia, Western Siberia, and the Altai Mountains.
The availability of a full allelic profile of the Y-chromosome of the man (see Table 2) made it possible to assign it, using the predictor program, to haplogroup Q, which is most typical of modern native population of Siberia and Central Asia. The presence of this haplogroup (with a low frequency) in some populations of Western Eurasia is usually explained by a geneflow from the East (from Central Asia). Thus, this structural variant of the Y-chromosome is also a marker of an Eastern Eurasian component of the Bertek-56 population’s gene-pool. At present, Y-chromosome variation in ancient populations is very poorly understood: there are only data on single specimens or very small samples. According to the results of study of a small sample of the Andronovo culture (Keyser et al., 2009; Allentoft et al., 2015), as well as of few specimens of the Afanasievo culture (Allentoft et al., 2015), haplogroup Q was not typical of those two archaeological cultures. Reference data on the Bronze Age Southern Siberia and Central Asia population are rather scarce: we can only note the presence of a variant of haplogroup Q in an individual of the Karasuk culture (Late Bronze Age) from Minusinsk Basin (Ibid.). Later, in the middle of the 1st millennium BC, this haplogroup was represented in the gene-pool of early nomads from the south of Inner Mongolia (China) (Zhao et al., 2010). In Altai, it was still present in the Early Iron Age (few specimens, see: (Allentoft et al., 2015)). A search for completely identical allelic profiles of the 17 Y-chromosome STR-loci in the “Y-Chromosome STR Haplotype Reference Database” has revealed that such variants are present in some modern individuals from Northeast China and South Korea (though it should be noted that populations from Northern Eurasia are poorly represented in this database, which can bias the results).
Thus, variants of mtDNA and Y-chromosome found in the man are markers of a component of Bertek’s genepool that is either autochthonous for Southern Siberia, or originates from adjacent regions of Eastern Eurasia. This finding concurs with the prevalence of the “Eastern stock” markers in his dental morphology (Chikisheva, 2012: 130). We have not found any genetic evidence of individual admixture effect (e.g. very strong genetic difference between his parents), and our data do not support the hypothesis of mixed ancestry for him. At the same time, the presence of phylogenetically and phylogeographically contrasting variants of mtDNA in the gene-pool of the group suggests a mixed origin for the population. The “mosaic” cranial and dental morphology of the Bertek-56 individuals, noted in previous studies, was probably not a result of admixture in the previous generation. Rather, it points towards the mixed origin of the population in general, as a result of admixture between strongly morphologically and genetically diverse groups of people. “Eastern” elements could have originated from people of Karakol and/or Okunev cultures of Southern Siberia (which is in line with findings of physical anthropologists), while “Western” elements cannot yet be associated with any particular ancient population of Western origin.
Notably, the ceramics found at the site are unusual for the Altai Mountains region (Molodin et al., 2004). Very similar vessels were found during the excavation of a ritual complex on Kucherla River (Molodin, Efremova, 2001), where the Bronze Age cultural layer was not identified at all (Molodin, Efremova, 2010). In the light of our results, it is important that very similar burial structures and ceramics were found at the Bronze Age Akbats burial ground in the Mangyshlak Peninsula (Balandina, Astafieva, 2000). This region might have been a source of migratory influx arriving in the Ukok Plateau. Importantly, the burial rite of Bertek-56 differs from that of the northerly Karakol culture (Kubarev, 1988; Molodin, 2006), which is in all respects closer to Okunev culture, as well as from the southerly Chemurchek culture (Drevneishiye evropeitsy…, 2014). Probably, we are dealing with a distinct ethnocultural phenomenon that combines an autochthonous base with a Western “coloring”. This was previously shown by archaeological and anthropological data, and has been further confirmed by our paleogenetic data.
Further reconstruction of complex ethnogeneses of South Siberian populations of the Bronze Age and later periods will largely depend on the accumulation of new genetic data on numerous ancient groups. Not only the study of various polymorphisms (mtDNA, Y-chromosome, and others), but correct interpretation of the obtained results in the light of archaeological and anthropological data, will be crucial. This approach is being employed by our interdisciplinary team in several ongoing research projects.
