Адаптационные стратегии гомининов и хищников на территории Западной Европы в раннем плейстоцене

Автор: Гарсиа Гаррига Х., Мартинес К., Ираведра Х.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 2 т.44, 2016 года.
Бесплатный доступ
Многие материалы, обнаруженные в последние годы, свидетельствуют о том, что заселение Европы гомининами в эпоху раннего плейстоцена происходило на протяжении более миллиона лет. По некоторым вопросам первоначального заселения европейского континента, в частности о том, был ли этот процесс последовательным и постоянным или прерывался, возникают споры среди специалистов. В статье дается обзор доступных зооархеологических материалов раннего плейстоцена, открытых на территории Европы, и обсуждается вопрос о влиянии добычи мясных ресурсов на поведенческие особенности гомининов. Наличие фаунистических остатков на европейских стоянках указывает на то, что гоминины имели доступ к разнообразным источникам пищи животного происхождения: от мелких животных (птицы, рептилии) до крупных млекопитающих (гиппопотамы, копытные), а также побеждали в конкурентной борьбе с хищниками, такими как гиены и кошачьи за одни и те же природные ресурсы. Климатические колебания, характерные для исследуемого периода, а также разнообразие экосистем, выявленных в Средиземноморском регионе и на континенте в целом, сделали потребление мяса основным условием жизнедеятельности в рамках адаптационных возможностей местных гомининов. Таким образом, непрерывность обитания гомининов в Европе в течение раннего плейстоцена могла зависеть от социальной сплоченности групп и их способности регулярно обеспечивать себя запасами мяса.
Ранний плейстоцен, западная европа, непрерывность популяции, стратегии жизнеобеспечения гоминин, потребление мяса
Короткий адрес: https://sciup.org/145145755
IDR: 145145755 | УДК: 903.4 | DOI: 10.17746/1563-0102.2016.44.2.019-029
Текст научной статьи Адаптационные стратегии гомининов и хищников на территории Западной Европы в раннем плейстоцене
Первоначальное заселение Европы человеком является предметом активных дискуссий на протяжении многих лет. Специалисты обсуждают хронологию этого процесса [Muttoni, Scardia, Kent, 2013; Garcia et al., 2014], маршруты передвижения популяций [Rook, Martínez-Navarro, Howell, 2004], факторы, обусловившие их расселение по Европе и Азии [Turner, 1992; Arribas, Palmqvist, 1999], в частности значимость климатических условий и окружающей среды [Rodrigues-Gómez et al., 2014], характер заселения – постоянный или прерывистый [Garcia, Martínez, Carbonell, 2011; Bermúdez de Castro et al., 2013]. По мнению ученых, в раннем плейстоцене расселение гомининов по Европе, вероятно, было спорадическим, в виде волн миграции, в периоды между которыми континент характеризовался малонаселенностью и даже опустением [MacDonald et al., 2012]. Данная гипотеза подтверждается малочисленностью останков гомининов, относящихся к 1,4–0,7 млн л.н. Кроме того, предполагается, что гоминины были неспособны заселять территории севернее 50° с.ш. (хотя этому противоречат свидетельства деятельности гомининов, обнаруженные на местонахождениях Хэпписбург 3 в Англии [Parfitt et al., 2010] и Унтермассфельд в Германии [Landeck, 2010; Garcia et al., 2013; Landeck, Garcia Garriga, 2015; Landeck, Garcia Garriga, in press]). Стабильная палеоэкологическая обстановка в эпоху раннего и среднего плейстоцена, которую отражают материалы памятника Сьерра-де-Атапуэрка, и археологические технокомплексы, последовательно перекрывающие друг друга на стоянках Барранко Леон D и Фуэнтэ Нуэва 3 (Орсе), Вальпарадис (Барселона), Сима дель Элефанте и Гран Долина TD6 (Атапуэрка), указывают на постоянное обитание гомининов в Южной Европе.
Объем археологических данных, отно сящих-ся к эпохе раннего плейстоцена в Европе, постоян- но увеличивается, т.е. расширяется документальная база для изучения причин, основ и динамики первоначального заселения континента. Каменные индустрии и/или антропологические останки периода, предшествующего субхрону Харамильо, которые обнаружены на стоянках Барранко Леон D и Фуэн-тэ Нуэва 3, находки из Сима дель Элефанте, Пирро Норд, пещеры Козарника, Унтермассфельд свидетельствуют о появлении в Европе групп гомининов 1,4–1,1 млн л.н. Следы присутствия гомининов, соответствующие приблизительно субхрону Харамильо, зафиксированы на стоянках Унтермассфельд, Ка’Бельведере ди Монте Поджиоло, Понт де Лавад, Лунери-Росьерес и Ле Валлонне. Период после эпизода Харамильо (0,98–0,78 млн л.н.) представлен значительно большим количеством стоянок: Вальпарадис, Гран Долина TD6, Пакфилд и Хэпписбург 3, Солейяк и Королево (см. рисунок и таблицу). Материалы всех этих стоянок подтверждают предположение о постоянном заселении региона гомининами, вероятно, с 1,4 млн л.н. до границы Матуяма–Брюнес [Garcia, Martínez, Carbonell, 2011; Garcia et al., 2014]. По мнению Муттони и соавторов [Muttoni, Scardia, Kent, 2013], самое раннее заселение Европы гомининами должно относиться к 0,85-0,78 млн л.н.
Согласно зооархеологическим и тафономическим данным, гоминины, обитавшие в средних и низких широтах, имели доступ к большому разнообразию животных, и это могло способствовать их адаптации к европейским экосистемам [Martínez et al., 2010; Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013; Landeck, Garcia Garriga, in press]. Широко известно, что до выхода за пределы Африки ок. 1,8 млн л.н. гоминины имели первичный доступ к тушам травоядных, однако исследователи пока не выработали единого мнения о том, какое место гоминины занимали в трофической цепи относительно крупных хищников ( Pachycrocuta brevirostris ,
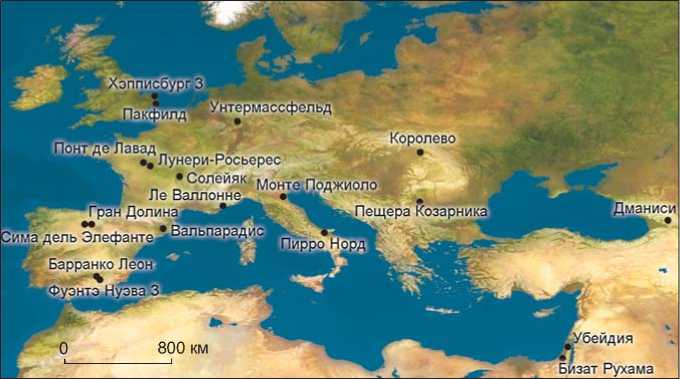
Рис. 1. Расположение стоянок, упоминаемых в статье.
Panthera gombaszoegensis или Homotherium latidens ). В настоящей статье мы, опираясь на тафономические свидетельства, которые приводятся в дискуссиях по теме «охотники или падальщики?», попытаемся обсудить предпосылки конкуренции между гоминина-ми и хищными животными и ее возможное влияние на последовательность или прерывистость заселения средних и низких широт Европы первыми человеческими популяциями.
Материалы и методы
Заселение Европы гомининами в период раннего плейстоцена рассматривается в статье с учетом зоо-морфологических и тафономических данных, собранных на наиболее важных стоянках в юго-западной части континента (см. рисунок и таблицу ). Наша цель – найти подтверждение того, что стратегии жизнеобеспечения первых поселенцев Европы, связанные с получением пищевых ресурсов животного происхождения, были достаточно надежными, чтобы поддерживать непрерывное воспроизводство популяции. Поскольку в центре нашего исследования – деятельность гомининов в отношении фауны, мы будем рассматривать только стоянки, на которых обнаружены зооархеологические и тафономические свидетельства: именно эти данные позволят оценить степень участия гомининов в формировании местонахождений фаунистических остатков (см. таблицу ).
Пиренейский полуостров в раннем плейстоцене
Местонахождения Фуэнтэ Нуэва 3 и Барранко Леон D (Орсе), Сима дель Элефанте и Гран Долина TD6 (Ата- пуэрка), а также Вальпарадис можно отнести к памятникам типа С по классификации Исаака [Isaac, 1983], т.к. на них представлена каменная индустрия в сочетании с останками нескольких особей травоядных животных. Однако эта связь не обязательно указывает на прямую (функциональную) зависимость между различными останками. Материалы многочисленных памятников свидетельствуют о случайности связей между фаунистическими останками и каменными артефактами. Таким образом, чтобы соотнести фаунистические материалы, найденные на стоянках, с деятельностью гомининов, необходимо изучить процессы осадконакопления и тафономии, способствовавшие формированию археологического объекта. Следовательно, изучение внешнего вида костей – насечек и следов ударов на них, а также их анатомического распределения позволяет получить убедительные аргументы в пользу предположения о воздействии гомининов на тушу животного. Анализ признаков этого воздействия, а также информации, например, о механизмах излома или кривых смертности помогает определить очередность получения гомининами доступа к туше животного и их род деятельности.
Бассейн Гвадикс-Баса
В бассейне Гвадикс-Баса на юге Испании было найдено несколько палеонтологических и археологических стоянок разного возраста. Стоянки, демонстрирующие несомненную связь между каменными артефактами и фаунистическими останками – Фуэнтэ Нуэва 3 и Барранко Леон D, датируются 1,4–1,2 млн л.н. [Toro-Moyano et al., 2009; Espigares et al., 2013]. На стоянке Барранко Леон D обнаружены фрагменты костей травоядных, на которых определены насечки, спира-
Характеристики фаунистических остатков с основных археологических стоянок эпохи раннего плейстоцена в Западной Европе
|
т О 1— О X |
со , ” 8 СО I Й S со g со - £ 2 о 2 s 2 ° 5 ” Б g о 8 а 5 2 2 „ 2 ” Б Б § В 2™ 2 О'- о го го„ ф „СМ СМ1 о 5 - Ф 2 О со 5 о 2 — CD СО СО _ И 1 о СО Д О А О Го ^ СО CO - & ” О Р О О Р О — СО С О го СМ СМ го СО СО CD О СО CM CD О = О LL СО СМ 2 2 о и I о ш > со о |
|
Ф ш $ о я 5 ° 5 5 О т Ш |
^ § ^8 i ^ 8 i g ^ ^ СП ° о го S £ О s О 5 5 ГО $ ° ° т £ ГО 2 го CD О О 5 я Го ’5 5 Го о 5 -В >5 5 х m 1 S Р 5 2 ГО х ’^ ГО X >Х £Q £Q £QQ) £0 X О-ХШО-^ФОХО. т О 0) I S 1_ск1— о CDO Ф ^ л о 1= FC ^ ^ С С |
|
3 5 Х ° CL ПС л 9 Р F ш Н m ™ е X о СО |
-го 3 S S X ГО SS X S Я ГО о О Т Ш о ГО х А н О dc;x А о фх о ГО о х ф х о о^ффхф Т НО- ш хт н шт т |
|
ш 2 и X |
и + + + + + + + + + О Т + с; х |
|
ш .2 СО X О и; ^ х о 5 х О СЕ И 0) X ГО ГО X Т О |
2 - ? го ш о F ^ со ^ + + с; ГО + сч + + ф + + + + со Ф ° О |
|
5 х -0 со g S со а 1 с |
о _ о Ш СО ^ с; с О О s ю 0Q |
|
О |
g го 8 о го ° X О ° Ю S X S X 2 X - ^ X СЕ S х х СЕ (П х СЕ Го т т X Е X ф Го О о х о. ГО с; х о Е^ ОО О е^ ш о О Е^ |
|
ГО X и го с |
■S 3 ’S 3 2 а н ф ф Ос О с: |
|
го СО X |
°ч °ч ^ Т "Г CN ООО ” о СО 4 4 ^ О О о о о т-" т-" О |
|
го с: |
Q о о СО е Ш О С" С" CQ R 20 С" |
левидные или винтообразные изломы, а также точки нанесения ударов и негативы сколов, и костяные от-щепы. Например, на фрагменте ребра Hippopotamus antiquus имеется насечка, появившаяся в процессе потрошения туши [Toro-Moyano et al., 2013]. В верхнем археологическом слое стоянки Фуэнтэ Нуэва 3 найден неполный скелет слона в связи с 34 копролитами Pachycrocuta и 17 каменными орудиями [Espigares et al., 2013]. Наличие таких свидетельств предполагает, что гоминины и хищники могли поочередно иметь доступ к туше этого животного. Однако прямых доказательств того, что этот представитель отряда хоботных был съеден гомининами или хищниками, нет. Кости слона не имеют каких-либо насечек, следов ударов или изломов. Тем не менее их залегание в контексте с каменными орудиями может указывать на воздействие гомининов на тушу животного.
В верхнем и нижнем археологических слоях памятника Фуэнтэ Нуэва 3 выявлены два достаточно сходных комплекса. Материалы верхнего слоя почти не несут следов деятельности гомининов, а нижнего – содержат олдовайские орудия и несколько большеберцовых и лучевых костей крупных млекопитающих (от 340 до 907 кг) со следами свежевания и расчленения [Ibid.]. По малочисленным костным останкам со следами антропогенного воздействия из нижних слоев этой стоянки невозможно определить, с какой стратегией они связаны – охоты или падальщичества, хотя некоторые насечки на подвздошных костях таза, ребрах и бедренных костях могут означать, что первыми доступ к туше получили все-таки гоминины и только потом – хищники [Ibid.]. Кроме того, следы ударов на диафизах некоторых лучевых, бедренных и большеберцовых костей указывают на извлечение костного мозга. Следовательно, сначала было съедено мясо, а затем и костный мозг. В нижних слоях Фуэнтэ Нуэва 3 отражением воздействия хищников на кости являются немногочисленные ямообразные отметины и бороздки от зубов. В верхних слоях были найдены кости лишь с отметинами зубов и копролиты гиены Pachycrocuta .
Сьерра-де-Атапуэрка
Раннеплейстоценовые свидетельства присутствия го-мининов в Сьерра-де-Атапуэрке (Бургос, Северная Испания) были зафиксированы на стоянках Сима дель Элефанте и Гран Долина [Bermúdez de Castro et al., 1997; Carbonell et al., 2008]. Здесь, помимо останков гомининов, были найдены в сочетании с олдовайски-ми орудиями фаунистические материалы [Carbonell et al., 1995, 2008]. Выявленные насечки, в частности на костях гомининов, были о ставлены людьми [Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013, in press]. Разные слои стоянки Сима дель
Элефанте подвергались различным тафономическим процессам [Rodríguez et al., 2011; Huguet et al., 2013, in press]. Насечки на нескольких костях оленя, бизона и черепахи, залегавших в слое ТЕ9, свидетельствуют о таких процессах, как снятие шкуры, разделка и свежевание туши [Ibid.]. Наличие насечек на костях осевого скелета и конечностей этих животных предполагает, что гоминины первыми получили доступ к тушам. Однако ввиду ограниченности остеологического материала невозможно сделать надежный вывод о характере этого воздействия. К этому следует добавить, что кости, подвергшиеся воздействию хищников, найдены во всех слоях памятника. И несмотря на малочисленность таких находок, можно предположить, что гоминины и хищники конкурировали между собой за доступ к тушам животных. Небольшой объем материала не позволяет определить, какая из сторон в большей степени способствовала аккумуляции костей, обнаруженных в Сима дель Элефанте. Однако, судя по насечкам, типичным для процесса отделения мяса, можно предположить, что по меньшей мере часть животной добычи была принесена на стоянку гомининами.
В Гран Долине слои различаются по объему свидетельств деятельности гомининов. В TDW4 накопление костных останков происходило естественным путем: это место медведи использовали для зимней спячки, и их кости были погребены в результате разрушения каменных глыб. Однако на костях мясных частей были зафиксированы следы потрошения и свежевания, которые можно считать признаками того, что гоминины первыми получили доступ к туше. На данной стоянке, как и на других рассматриваемых объектах, кости со следами деятельности хищников составляют 9,2 % выборки [Huguet et al., 2013]. Судя по распределению костей, обнаруженных на стоянке Гран Долина TD6, частям скелета и следам воздействия на них, гоми-нины, несмотря на конкуренцию с хищниками, имели доступ к туше, именно они до ставляли на стоянку основную часть добычи [Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013; Rodríguez-Gómez et al., in press]. Это предположение подтверждается данными о доле костей с насечками (13 %) и наличием следов зубов поверх насечек [Huguet et al., 2013]. Различные следы свидетельствуют о том, что доступ к тушам был неограниченным, с них снимали шкуру, потрошили и отделяли мясо и т.д. Как отмечают Диез с соавторами [Díez et al., 1999], транспортировка добычи происходила по-разному, в зависимости от размеров животных: тушу небольших и средних размеров приносили на стоянки целиком. По мнению Салади и его коллег [Saladié et al., 2011], туши некоторых крупных животных, возможно, доставлялись целиком, что предполагало сотрудничество гомининов и совместное использование ресурсов.
Среди находок со стоянки Гран Долина TD6 определены кости со следами зубов хищников. Поскольку следы находятся поверх насечек, можно предположить, что животные подбирали падаль, оставленную гомининами [Díez et al., 1999; Saladié et al., 2011; Huguet et al., 2013]. Согласно материалам слоя TD6-2, хищники получали значительную часть туш копытных, после того как гоминины съедали бóльшую часть мяса [Saladié et al., 2014]. В слое TD6 обнаружены многочисленные ко сти с насечками, принадлежавшие Homo antecessor . Эти находки считаются самыми ранними свидетельствами каннибализма в эволюции человека. Такая форма поведения, как постоянный каннибализм обусловлена дефицитом пищи и воды [Carbonell et al., 2010]. Однако, как показали материалы стоянки Гран Долина TD6, гоминины были способны первыми получить доступ к тушам животных. Исследованиями установлено, что по возрастному составу группы гомининов, останки которых найдены в этом слое, аналогичны популяциям шимпанзе, у которых каннибализм был результатом межгрупповой агрессии [Saladié et al., 2012]. Каннибализм среди го-мининов, возможно, являлся частью стратегии адаптации к стрессовым ситуациям, связанным с нехваткой еды; актом, призванным предотвратить поедание хищниками тел гомининов после того, как их сородичи покидали стоянку, или же результатом межгрупповой конкуренции. В любом случае важно, что охота людей на крупные стада копытных сопровождалась актами каннибализма. Возможно, эта практика не была связана с заботой о пропитании и экономии ресурсов.
Вальпарадис
Сектор EVT7 (археологические слои 10 и 10с) памятника Вальпарадис (Барселона, Северо-Восточная Испания) является наиболее насыщенным в археологическом отношении. Он был раскопан на площади более 780 м2 и глубину 3 м. Возраст отложений в этом секторе определен с помощью биостратиграфического анализа остатков мелких и крупных млекопитающих, а также методами палеомагнитного, ЭПР- и ОСЛ-датирования. Выявленная отрицательная полярность указала возраст памятника – между эпизодом Харамильо и границей Матуяма–Брюнес. С учетом найденных остатков грызунов Mimomys savini и Iberomys huescarensis Вальпарадис был отнесен к биозоне Allophayomis chalinei [Martínez et al., 2010]. По результатам сравнения нижних первых моляров отдельных взрослых особей полевок Mimomys savini, обнаруженных в секторе EVT7, и экземпляров из соответствующих стратиграфических уровней памятников Гран Долина, Фуэнтэ Нуэва 3 и Барранко Леон D, Вальпарадис датирован периодом ок. 0,98–0,95 млн л.н. (МИС 27) [Martínez et al., 2014; Garcia, Martínez, 2015]. Это определение дополняют даты 0,83 ± 0,13 и 0,849 ± 0,48 млн л.н. для археологического слоя 10 сектора EVT7 и 0,858 ± ± 0,87 млн л.н. для сектора EVT8, установленные ЭПР-и ОСЛ-методами по зубам лошадиных и зернам кварцита [Martínez et al., 2010; Duval et al., 2011, 2015].
Свидетельства активности гомининов на стоянке и исходное положение части археологического материала были рассмотрены в статье Гарсиа и его соавторов [Garcia et al., 2012]. Богато представленный олдовайский комплекс, основанный на биполярном расщеплении с применением наковальни, с остатками травоядных, на которых фиксируются насечки и следы дробления, о ставленные гомининами, был обнаружен в секторе EVT7 [Garcia, Martínez, Carbonell, 2013]. Анализ распределения археологических материалов в этом секторе по вертикали позволяет выделить три разных комплекса каменных и фаунистических материалов и сделать предположение о нескольких эпизодах седиментации на местонахождении. Поскольку методом ремонтажа удалось восстановить четыре каменных изделия, есть надежда, что изучение последовательности отложений поможет определить исходное положение части материалов. Макрофаунистический комплекс сектора EVT7 может быть определен как эпивиллафранкский. Он представлен, как показал тафономический анализ, ко стными остатками, связанными с жизнедеятельностью гиены Pachycrocuta brevirostris – падальщика: большая часть костей имеет следы обгладывания и много отметин цилиндрической формы [Madurell-Malapeira et al., 2012], обнаружены также фрагменты срыгнутых костей и копролиты [Martínez et al., 2010; Garcia et al., 2012].
Следы разделки туш травоядных гомининами зафиксированы на нескольких костях очень крупных животных, таких как представители семейств носороговых Rhinocerotidae и бегемотовых Hippopotamidae, а также на остатках травоядных средних размеров – представителей семейств лошадиных Equidae и оленевых Cervidae. Процессы разделки, освежевания и снятия шкур получили отражение на 12 элементах костей верхней части передних и задних конечностей и позвоночника; признаки дробления, произведенного гомининами, прослежены на 5 фрагментах [Martínez et al., 2010; Garcia et al., 2012]. Половина костей с насечками также имеет отметины от зубов. Следовательно, и гоминины, и крупные хищники имели доступ к одной добыче. Отметины, оставленные на одних и тех же костях, могут свидетельствовать о конкуренции между представителями обеих групп за обладание добычей, а также о том, что падальщики употребляли остатки пищи, брошенные хищниками. До сих пор не найдены перекрывающие друг друга следы от зубов и орудий, которые позволили бы опре- делить последовательность доступа представителей обеих групп к туше животного. Поскольку на данной стоянке ко стей со следами антропогенного воздействия немного, а остатки с признаками воздействия хищников, напротив, представлены широко, предполагается, что деятельность гомининов была незначительной, здесь обитали в основном гиены Pachycro-cuta brevirostris.
Памятники на территории Италии и Франции
К памятникам позднего этапа раннего плейстоцена, позволяющим получить информацию о первоначальном заселении Европы, относится стоянка Солейяк во Франции. Ее отложения залегали вместе с фаунистическими остатками (более 3 тыс. экз., по которым выделены 29 особей) и каменными артефактами [Bonifay, Bonifay, 1981]. Гоминины могли иметь отношение к накоплению большей части этого комплекса костей, но плохая сохранность фаунистических остатков затрудняет их идентификацию. Убедительным выглядит только определение костей оленя [Fosse, 1994]. На стоянке Ле Валлонне во Франции также были обнаружены фрагменты костей, относящиеся к раннему этапу среднего плейстоцена. Их связь с каменной индустрией сомнительна; многочисленные фаунистические материалы позволяют сделать вывод о низкой степени воздействия на них гомининов: следы человеческой деятельности имеются исключительно на костях оленевых [Echassoux, 2009].
В Италии древнейшие свидетельства присутствия человека обнаружены на памятнике Пирро Норд, но зооархеологические остатки здесь не выявлены. Материалы других памятников, например, Ка’Бельведере ди Монте Поджиоло могут дать представление о некоторой контекстуальной связи фауны и каменных артефактов [Villa, Lenoir, 2009]. Обнаруженные на этих стоянках каменные орудия и кости с насечками являются отражением деятельности гоми-нинов, а фаунистические остатки – деятельности хищников. На стоянках Ле Валлонне [Echassoux, 2009] и Пирро Норд [Arzarello et al., 2012] были представлены фрагменты костей гиены Pachycrocuta , что позволяет сделать предположение о конкуренции между гомининами и хищниками за доступ к добыче или же об ограничении деятельности гомининов вмешательством хищников.
Обсуждение
Зооархеологические и тафономические материалы раннего плейстоцена, зафиксированные на Пиреней- ском полуострове, свидетельствуют о том, что го-минины имели возможно сть добывать разных животных – от мелких животных (птицы и рептилии) до крупных млекопитающих (бегемоты и копытные различных размеров, в частности оленевые и полорогие) [Martínez et al., 2010; Rodríguez et al., 2011; Saladié et al., 2011, 2014; Huguet et al., 2013, in press] (см. рисунок и таблицу). Прослеженные по материалам стоянок Сима дель Элефанте, Гран Долина, Орсе и Вальпарадис следы деятельности хищников чаще всего синхронны свидетельствам воздействия гомининов. Поскольку на одной стоянке сосуществование гомининов и хищников было невозможно, то наличие на ней следов деятельно сти и тех, и других, возможно, следует связывать с нехваткой пищевых ресурсов и конкуренцией за них или заселением этими группами одних зон в ранний период плейстоцена. Некоторые исследователи, учитывая эти отношения между хищниками и гомининами, считают, что первое население Европы зависело от наличия или отсутствия хищников. Согласно этой точке зрения, гоминины, мигрировавшие из Африки, были падальщиками; они подбирали добычу, оставленную хищниками [Turner, 1992; Arribas, Palmqvist, 1999]. Вместе с тем ввиду вымирания гиен вида Pachycrocuta сложились более благоприятные условия для распро странения гомининов в Европе: уменьшилась пищевая конкуренция, и гоминины получили доступ к большему количеству добычи [Turner, 1992]. Однако, по мнению ряда специалистов, трофическое давление хищников не оказывало какого-либо влияния на поведенческие особенности гомининов и их адаптацию к местным природно-климатическим условиям [Echassoux, 2009].
На территории Европы хищники обитали в течение всего плейстоцена. В начале раннего плейстоцена они по доле в общей численности и количеству видов с узкой специализацией превосходили травоядных. Видовое разнообразие увеличивалось за счет мигрировавших в Европу представителей новых видов, лучше адаптированных к открытым сухим ландшафтам, таких как волк Lycaon, гиена Pachycrocuta brevirostris, этрусский волк Canis etruscus или этрусский медведь Ursus etruscus [Croitor, Brugal, 2010]. Переход от раннего плейстоцена к среднему ознаменовался вымиранием одиночных крупных хищников: гепарда Acynonix, саблезубых кошек родов Megantereon, Dinofelix и Homotherium, древней рыси Lynx issidorensis. Освободившуюся нишу заполнили более мелкие плотоядные, чье поведение предполагало жизнь в стае с более широким спектром потребляемых ресурсов [Ibid.]. Среди находок со стоянки Гран Долина TD6 велика доля костей с насечками; кости представляют животных нескольких видов и все отделы скелета. Установлено, что зафиксированные на ко стных о статках следы применения орудий появились в процессе свежевания, потрошения, снятия шкуры и разделки туши. Составленные по этим материалам возрастные профили выявили преобладание молодых особей. Эти свидетельства позволили сделать предположение о том, что аккумуляция костных остатков происходила под влиянием в основном антропогенного фактора. Оно подтвердилось, когда были обнаружены на костях отметины от зубов хищников поверх следов от орудий, т.е. воздействие животных было вторичным [Saladié et al., 2011, 2014; Huguet et al., 2013]. Все это позволяет заключить, что в этот период охотничьи стратегии могли носить регулярный характер, а принесенная на стоянку добыча делилась между всеми членами социальной группы.
Необходимо добавить, что обитатели других стоянок охотились на животных на берегах рек и различных водоемов, где конкуренция с хищниками была максимально высокой. Это доказывают материалы памятников Фуэнтэ Нуэва 3, Барранко Леон D и Вальпа-радис. Ранний или даже первичный доступ гомининов к туше может быть идентифицирован при наличии насечек на костях мясных частей туш животных средних размеров, например, оленевых. Как показали экспериментальные исследования, подобные следы не часто встречаются на костях наиболее питательных частей туши после воздействия на нее хищников [Gidna, Yaravedra, Domínguez-Rodrigo, 2012]. Первичный доступ гомининов к тушам, примеры которого мы наблюдаем в нижнем плейстоцене Европы на стоянке Гран Долина TD6, связан с высоким трофическим давлением в некоторых ареалах обитания. Так, следы от орудий, идентифицированные Эспигаресом и соавторами [Espigares et al., 2013] на ребрах, подвздошной кости таза и диафизе бедренной кости из нижних слоев стоянки Фуэнтэ Нуэва 3, как и кости позвоночника, передних и задних конечностей со следами разделки орудиями с памятника Вальпарадис, которые были исследованы Мартинесом и его соавторами [Martínez et al., 2010], не соответствуют поведению гомининов как падальщиков и, соответственно, свидетельствуют об их первичном воздействии на добычу.
Что касается порядка, в котором гоминины и/или хищники получали доступ к тушам на стоянках Валь-парадис и Фуэнтэ Нуэва 3, то такую последовательность установить невозможно ввиду отсутствия перекрывающих друг друга следов воздействия. Различия, выявленные по тафономическим данным объектов, могут быть связаны с местом расположения или специфическими функциями памятников. Так, стоянки Орсе и Вальпарадис находятся на открытых пространствах вблизи водоемов, тогда как Гран Долина TD6 является пещерным памятником закрытого типа с признаками интенсивной поселенческой деятельности [Saladié et al., 2011, 2014; Rodríguez-Gómez et al., in press]. Среди находок из этой пещеры наибольшую долю составили кости со следами от орудий и наименьшую – со следами воздействия хищников. Кости со стоянок Орсе и Вальпарадис, расположенных у водоемов, имеют повреждения, оставленные в основном крупными хищниками; воздействие последних могло уничтожить свидетельства деятельности гомининов, которые также представлены на памятнике.
Заключение
Археологическая последовательность, прослеженная по материалам памятников Орсе, Атапуэрка и Валь-парадис, отно сится к периоду от 1,4–1,2 млн л.н. до границы Матуяма–Брюнес (0,78 млн л.н.). Следовательно, процесс заселения Пиренейского полуострова гомининами был продолжительным [Garcia et al., 2011, 2014]. Тафономическое и зооархеологи-ческое изучение показало, что материалы всех этих пиренейских памятников, по аналогии с евразийскими, соответствуют типам основных адаптационных стратегий, которые предполагают наличие сплоченных социальных коллективов, спо собных регулярно обеспечивать себя мясом [Martínez et al., 2010; Saladié et al., 2011, 2014; Huguet et al., 2013, in press]. Кроме того, на всех этих стоянках обнаружены комплексы каменных орудий, близкие к олдовайскому технокомплексу, при некоторой вариабельности [Garcia et al., 2013]. Такие адаптационные и технологические характеристики предполагают приспособление гоми-нинов к различным эко системам и климатическим колебаниям, зафиксированным в течение рассматриваемого периода.
Фаунистический материал европейских стоянок представляет много разнообразных таксонов копытных, что позволяет сделать вывод об обеспеченности гомининов животной пищей. При этом к концу раннего плейстоцена соотношение хищников и травоядных изменяется в пользу последних [Croitor, Brugal, 2010]. Влияние этих факторов могло способствовать расселению гомининов в Европе в течение нижнего плейстоцена. Обсуждаемые здесь материалы пиренейских памятников, как и свидетельства из других частей Европы, позволяют рассматривать охоту как довольно раннюю стратегию. На таких евразийских памятниках, как Гран Долина TD6 [Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013; Rodríguez-Gómez et al., in press], Фуэнтэ Нуэва 3 [Espigares et al., 2013; Huguet et al., 2013], а также в некоторых слоях Убейдии [Belmaker, 2006] и Би-зат Рухамы [Zaidner, Yeshurun, Mallol, 2010] обнаружены явные свидетельства деятельности гомининов. Находки со стоянки Гран Долина TD6 доказывают, что гоминины первыми получали до ступ к тушам животных (и телам гомининов) весом от 10 до 400 кг с целью потребления мяса посредством либо охоты либо подбирания туш крупных млекопитающих, таких как слоны или бегемоты [Díez et al., 1999; Saladié et al., 2011, 2014; Huguet et al., 2013].
Материалы памятников Сима дель Элефанте и Барранко Леон D [Huguet et al., 2013], Вальпарадис [Martínez et al., 2010; Garcia et al., 2011, 2012, 2014], Унтермассфельд [Landeck, Garcia Garriga, in press] или Дманиси [Tappen et al., 2007] также содержат свидетельства потребления мяса гомининами, а кроме того, и хищниками. В основном гиеновые [Ibid.; Madurell-Malapeira et al., 2012] и кошачьи [Arribas, Palmqvist, 1999] были причастны к накоплению фаунистических остатков на стоянках. Вероятно, гоми-нины и хищники были вынуждены конкурировать в борьбе за одни и те же природные ресурсы, во время расселения по Европе и Азии гоминины сталкивались с такими серьезными проблемами, как пищевой стресс и вынужденная адаптация к новым условиям окружающей среды.
Поедание крупных животных, например бегемотов, жителями стоянок Вальпарадис, Убейдия, Унтермассфельд или Орсе можно рассматривать как следствие спорадического освоения ими ресурсов. Наличие скелетов в местах обитания соответствующих животных можно объяснить естественной смертью последних. Возможно также, что сначала на них охотились крупные кошачьи, а затем доступ к добыче получили гоминины. Даже если гоминины являлись вторичными потребителями, они все равно напрямую конкурировали с крупными представителями гиеновых за большие туши; в этом случае у го-мининов была более активная роль падальщиков, чем та, которую описывает Тёрнер [Turner, 1992]. Данные о стабильных климатических условиях в южных рефугиумах, каким являлся и Пиренейский полуостров, а также следы антропогенного воздействия на костях в наиболее мясных частях туш крупных травоядных могут указывать на существование долговременных поселений ранних людей на полуострове и расселение гомининов в северных районах Европы в более теплые периоды.
Список литературы Адаптационные стратегии гомининов и хищников на территории Западной Европы в раннем плейстоцене
- Arribas A., Palmqvist P. On ecological connection between sabre-tooths and hominids: faunal dispersal events in the Lower Pleistocene and a review of the evidence for the first human arrival in Europe // J. of Archaeol. Sci. - 1999. - Vol. 26, N 5. -P. 571-585.
- Arzarello M., Pavia G., Peretto C., Petronio C., Sardella R. Evidence of an Early Pleistocene hominin presence at Pirro Nord (Apricena, Foggia, southern Italy): P13 site // Quaternary Intern. - 2012. - Vol. 267. - P. 56-61.
- Belmaker M. Community structure through time: ‘Ubeidiya, a Lower Pleistocene site as a case study: Ph.D. dissertation, Hebrew Univ. - Jerusalem, 2006. - 284 p.
- Bermudez de Castro J.M., Arsuaga J.L., Carbonell E., Rosas A., Martinez I., Mosquera M. A hominid from the Lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and Modern Humans // Science. - 1997. -Vol. 276. - P. 1392-1395.
- Bermudez de Castro J.M., Martinön-Torres M., Blasco R., Rosell J., Carbonell E. Continuity or discontinuity in the European Early Pleistocene human settlement: the Atapuerca evidence // Quaternary Sci. Rev. - 2013. - Vol. 76. -P. 53-65.
