Ассоциация комплексных генотипов генов DGAT1, CSN3, LGB с хозяйственноценными признаками быков-производителей

Автор: Зиннатов Ф.Ф., Якупов Т.Р., Зиннатова Ф.Ф., Харисова Ч.А.
Статья в выпуске: 4 т.248, 2021 года.
Бесплатный доступ
Проведенные исследования с применением ДНК-анализа среди комплексных генотипов генов DGAT1, CSN3 и LGB показали, что у 63 животных идентифицировано 14 различных комплексных генотипов. Наивысшие показатели продуктивности принадлежат животным с сочетанием генотипов DGAT1ak CSN3bb LGBbb - 9261,5 кг и 3,7 % жира и 4,2 % белка. Их насчитывается 1,6 % из числа исследованных. Коровы с этим же сочетанием генотипов отличаются высоким выходом жира и белка - 342,6 кг и 388,9 кг.
Днк, пцр-пдрф анализ, генотип, быки-производители
Короткий адрес: https://sciup.org/142231414
IDR: 142231414 | УДК: 636. | DOI: 10.31588/2413-4201-1883-248-4-62-66
Текст научной статьи Ассоциация комплексных генотипов генов DGAT1, CSN3, LGB с хозяйственноценными признаками быков-производителей
Важнейшим генетическим резервом дальнейшего улучшения молочного поголовья коров являются быки-производители, полученные от лучших представителей генофонда породы. Улучшение генетики молочного поголовья на 85-90 % зависит от племенной ценности быков-производителей. При интенсивном использовании выдающихся быков и их лучших сыновей, внуков, правнуков в странах США и Европы, в том числе и в Российской Федерации, созданы молочные стада высокопродуктивных коров. Рентабельность отбора определяется степенью подкрепляющего эффекта племенных быков, применяемых в каждом поколении. При этом, как показывают работы отечественных ученых, племенная ценность быков состоит из ряда характеристик и показателей. Отбор и идентификация быков-производителей по их способности передать желаемые экономически значимые признаки обеспечивают положительную динамику селекционной работы [3, 5].
Идентификация аллельных вариантов генов позволяет осуществлять прямой отбор на уровне ДНК в дополнение к традиционному отбору животных. Преимущество анализа ДНК в том, что с помощью ДНК-анализа можно определить генотип биологического объекта независимо от возраста, пола и физиологического состояния, что является важнейшим пунктом в племенной работе [4, 7].
Генетический маркер – ген, полиморфный признак, идентифицируемый методами молекулярной биотехнологии на уровне ДНК [1, 6]. Рассматриваемые нами генетические маркеры это: Ген диацилглицерин-O-ацилтрансферазы (DGAT1), выявлен как генетический маркер, влияющий на качество молока. Этот ген используется в биосинтезе липидов и связан с жирномолочностью коровьего молока. Ген капа-казеина (CSN3) связан с технологическими свойствами молока и молочным белком, указывает на оптимальные технологические свойства молока при производстве сыра. Ген лактоглобулина (LGB), который кодирует белок бета-лактоглобулин в молоке, отвечает за белок молока и идентифицирован как индикатор ценности молока и молочных продуктов [7].
Таким образом, целью работы является ДНК идентификация ассоциации комплексных генотипов генов DGAT1, CSN3, LGB с хозяйственно-ценными признаками быков-производителей и коров основного стада.
Материал и методы исследований. Для исследований были отобраны образцы крови от быков голштино-фризкой породы в количестве 17 голов, черно-пестрой – 46 голов племпредприятий и племрепродукторов Республики Татарстан ГУП ГПП «Элита» ООО АФМ «Лельвиж», ООО «Смаиль». Для проведения генетического мониторинга с целью увеличения этой популяции в племенной работе учитывали их родительский индекс
(РИБ). Показатель РИБ соответственно определяли по молочной продуктивности женских предков.
ДНК выделяли из лейкоцитов крови быков-производителей в количестве 100 мкл с применением стандартного набора ДНК-сорб-В (ООО «ДНК-технологии»), по методике представленной изготовителем. В работе была оптимизирована программа проведения ПЦР с некоторыми поправками в температурных и временных режимах реакции, что обеспечило оптимальную ПЦР апмлификацию фрагментов.
ПЦР-фрагменты генов диациглицерол-О-ацил трансферазы (DGAT1), капа-казеина (CSN3) и лактоглобулина (LGB) были подвергнуты гидролизу с применением ферментов рестриктаз AcoI, Hinf I и Hae III соответственно. Рестрикция проводилась по рекомендациям изготовителя. Молекулярный масса продуктов ПЦР-ПДРФ анализа определяли методом горизонтального электрофореза в 2,4-3 % агарозном геле.
Обработку данных производили с помощью программного обеспечения Microsoft Excel.
Частоту встречаемости генотипов определяли по формуле: p=n/N, где p-частота определения генотипа, n-количество особей, имеющих определенный генотип, N - число особей.
Результат исследований. В процессе ПЦР-ПДРФ анализа ДНК лейкоцитов крови быков-производителей были получены специфические фрагменты гена DGAT1 длиной 411 пар нуклеотидов, выявлено два аллеля – A и K и соответственно три генотипа – DGAT1AA, DGAT1KK, DGAT1AK. Гомозиготному генотипу AA соответствует 208/203 п.н., гетерозиготному генотипу AK – 411/208/203 п.н., гомозиготному генотипу KK – 411 п.н. Частота встречаемости гомозиготного генотипа АА составила 9,5 %, гетерозиготного генотипа АК – 87,3 %, гомозиготного генотипа КК – 3,2 %. Частота встречаемости аллеля А– 0,53, К – 0,47 (Таблица 1).
После проведения ПЦР-ПДРФ анализа ДНК крови быков-производителей, нами были получены специфические фрагменты гена CSN3 длиной 265 пар нуклеотидов, выявлено два аллеля – A и В и три генотипа – CSN3AA, CSN3ВВ , CSN3AВ. Гомозиготному генотипу AA соответствует 134-132/89 п.н., гетерозиготному генотипу AВ – 265/134-132/89 п.н., гомозиготному генотипу ВВ – 265/89 п.н. Частота встречаемости гомозиготного генотипа АА составила 73,1 %, гетерозиготного генотипа АВ – 20,6 %, гомозиготного генотипа ВВ – 6,3 %. Частота встречаемости аллеля А– 0,83, В – 0,17 (Таблица 2).
В результате ПЦР-ПДРФ анализа ДНК лейкоцитов крови быков-производителей с парой праймеров были получены специфические фрагменты гена LGB длиной 153 пар нуклеотидов. Выявлено два аллеля A и B и три генотипа LGBAA, LGBAB, LGBBB. Гомозиготному генотипу АА соответствуют фрагменты 153/109 п.н., гетерозиготному генотипу АВ – 153/109-74/79 п.н., гомозиготному генотипу ВВ – 109-74/79 п.н. (Рисунок 1).
Таблица 1 – Полиморфизм гена DGAT1 у быков-производителей
|
АА |
АК |
КК |
Частота аллелей |
||||
|
n |
% |
n |
% |
n |
% |
А |
К |
|
6 |
9,5 |
55 |
87,3 |
2 |
3,2 |
0,53 |
0,47 |
Таблица 2 – Полиморфизм гена CSN3 у быков-производителей
|
АА |
АВ |
ВВ |
Частота аллелей |
||||
|
n |
% |
n |
% |
n |
% |
А |
В |
|
46 |
73,1 |
13 |
20,6 |
4 |
6,3 |
0,83 |
0,17 |
Таблица 3 – Полиморфизм гена LGB у быков-производителей
|
АА |
АВ |
ВВ |
Частота аллелей |
||||
|
n |
% |
n |
% |
n |
% |
А |
В |
|
10 |
15,9 |
34 |
54 |
19 |
30,1 |
0,43 |
0,57 |
Таблица 4 – РИБ быков-производителей в зависимости от комплексных генотипов генов
DGAT1, CSN3 и LGB
|
Генотип |
Количество |
РИБ быков-производителей |
|||||
|
n |
% |
удой, кг |
массовая доля, % |
количество молочного, кг |
|||
|
жира |
белка |
жира |
белка |
||||
|
DGAT1AACSN3AALGBAA |
1 |
1,6 |
7923,5 |
3,1 |
3,2 |
245,6 |
253,6 |
|
DGAT1AACSN3AALGBAB |
2 |
3,2 |
7424,35 + 940,6 |
3,4 + 0,3 |
3,7 + 0,4 |
255,2 + 54,3 |
278,5 + 64,5 |
|
DGAT1AACSN3AALGBBB |
2 |
3,2 |
7089,6 + 260,1 |
3,5 |
3,95 + 0,15 |
248,1 + 9,1 |
279,6 + 0,36 |
|
DGAT1AACSN3ABLGBBB |
1 |
1,6 |
7336,7 |
3,2 |
3,7 |
234,8 |
271,5 |
|
DGAT1AKCSN3AALGBAA |
6 |
9,5 |
7385,5 + 992,2 |
3,3 + 0,3 |
3,4 + 0,3 |
247,01 + 45,2 |
249,9 + 46,3 |
|
DGAT1AKCSN3AALGBAB |
22 |
34,9 |
7220,4 + 691,2 |
3,5 + 0,3 |
3,6 + 0,2 |
251,5 + 35,6 |
258,5 + 31,2 |
|
DGAT1AKCSN3AALGBBB |
11 |
17,5 |
7224,6 + 248,9 |
3,4 + 0,2 |
3,7 + 0,14 |
243,2 + 18,7 |
268,6 + 13,6 |
|
DGAT1AKCSN3ABLGBAA |
5 |
7,9 |
8039,42 + 365,8 |
3,6 + 0,2 |
3,62 + 0,2 |
288,4 + 25,6 |
290,9 + 21,1 |
|
DGAT1AKCSN3ABLGBAB |
4 |
6,4 |
7269,5 + 614,9 |
3,4 + 0,18 |
3,6 + 0,29 |
246,2 + 12,5 |
262,1 + 14,3 |
|
DGAT1AKCSN3ABLGBBB |
3 |
4,7 |
7845,3 + 1519 |
3,53 + 0,3 |
3,83 + 0,2 |
279,9 + 68,9 |
304 + 73,1 |
|
DGAT1AKCSN3BBLGBAB |
3 |
4,7 |
7075,6 + 500,6 |
3,7 + 0,08 |
4,2 + 0,09 |
262,2 + 23,9 |
295,1 + 25,8 |
|
DGAT1AKCSN3BBLGBBB |
1 |
1,6 |
9261,5 |
3,7 |
4,2 |
342,68 |
388,9 |
|
DGAT1KKCSN3AALGBAB |
1 |
1,6 |
6483,8 |
3,1 |
3,3 |
200,9 |
213,9 |
|
DGAT1KKCSN3AALGBBB |
1 |
1,6 |
9151,3 |
3,9 |
3,7 |
356,9 |
338,6 |
200 п.н. —
100 п.н. —
50 п.н. —
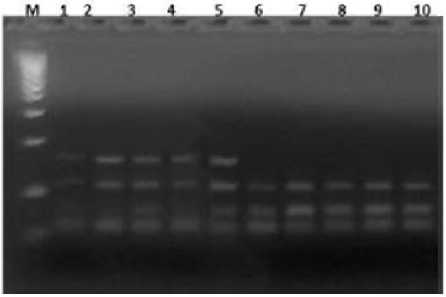
— 79-74 п.н
— 153п.н
— 109п.н.
Рисунок 1 – Электрофореграмма результата ПЦР-ПДРФ анализа гена бета- лактоглобулина (LGB). Обозначения: М-ДНК – маркеры 1500-50 bp (СибЭнзим); 1, 2, 3, 4, 5 – генотип АB (153/109/74-79); 6, 7, 8, 9, 10 – генотип ВВ (109/74-79 bp)
Частота встречаемости гомозиготного генотипа АА составила 15,9 %, гетерозиготного генотипа АВ – 54 %, гомозиготного генотипа ВВ – 30,1 %. Частота встречаемости аллеля А– 0,43, В – 0,57 (Таблица 3).
Особые сочетания генотипов имеют влияние на количество жира и белка в молоке. Поэтому было решено проследить закономерность сочетания генотипов генов DGAT1, CSN3 и LGB. Всего выявлено у 63 животных 14 различных комплексных генотипов (Таблица 4).
Наивысшие показатели продуктивности по РИБ принадлежат животным с сочетанием генотипов DGAT1AKCSN3BBLGBBB – 9261,5 кг и 3,7 % жира и 4,2 % белка. Коровы с этим же сочетанием генотипов отличаются высоким выходом жира и белка – 342,6 кг и 388,9 кг.
Заключене. Для увеличения в стадах животных с наивысшими показателями хозяйственно-ценных признаков следует проводить ДНК тестирование поголовья коров и быков-производителей по генам липидного и белкового обмена в животноводческих предприятиях Российской Федерации, что позволит повысить экономическую эффективность, молочную продуктивность и улучшить качество молока.
Резюме
Проведенные исследования с применением ДНК-анализа среди комплексных генотипов генов DGAT1, CSN3 и LGB показали, что у 63 животных идентифицировано 14 различных комплексных генотипов. Наивысшие показатели продуктивности принадлежат животным с сочетанием генотипов DGAT1AK CSN3BB LGBBB – 9261,5 кг и 3,7 % жира и 4,2 % белка. Их насчитывается 1,6 % из числа исследованных. Коровы с этим же сочетанием генотипов отличаются высоким выходом жира и белка – 342,6 кг и 388,9 кг.
Список литературы Ассоциация комплексных генотипов генов DGAT1, CSN3, LGB с хозяйственноценными признаками быков-производителей
- Ахметов, Т.М. Использование методов маркер-вспомогательной селекции в молочном скотоводстве Республике Татарстан / Т.М. Ахметов// Автореф. дис. докт. биол. наук. – Казань, 2009. – 50 с.
- Амерханов, Х.А. Нормы оценки племенных качеств крупного рогатого скота мясного направления продуктивности / Х.А. Амерханов, И.М. Дунин, В.И. Шаркаев [и др.]. – Москва, 2010. – 36 с.
- Габидулин, В.М. Русская комолая и аберин-ангусская породы в России и методы их совершенствования / В.М. Габидулин, М.В. Тарасов // Вестник мясного скотоводства. – 2010. – № 63 (2). – С. 7-11.
- Зиннатова, Ф.Ф. Анализ родительского индекса ремонтных-бычков и генотипирование их по генам каппа-казеина (CSN3), жирномолочности (DGAT) и BLAD / Ф.Ф. Зиннатова, A.M. Алимов, Ф.Ф. Зиннатов //Ученые записки КГАВМ. – Казань. – 2011. – Т. 206. – С. 81-85.
- Зиннатова, Ф.Ф. Изучение влияния комплексных генотипов генов CSN3, DGAT1, TG5, PRL, LGB на показатели родительского индекса быков / Ф.Ф. Зиннатова, А.М. Алимов, Ш.К. Шакиров, Ф.Ф. Зиннатов // Ученые записки Казанской ГАВМ им. Н. Э. Баумана. – 2013. – Т. 215. – С. 126-129.
- Идиатулин, Р.И. Совершенствование мер обеспечения безопасности продовольствия / Р.И. Идиатулин, Н.М. Калишин, Д.А. Орехов //Вопросы нормативно правового регулирования в ветеринарии. – 2012. – № 3. – С. 7-15.
- Орлова, Д.А. Изучение показателей качества сыров, фальсифицированных компонентами немолочного происхождения. / Д.А. Орлова, Т.В. Калюжная, А.С. Смолькина, А.Н. Токарев, А.В. Дрозд // Международный вестник ветеринарии. – 2018. – № 2. – С. 82-86.
- Рахматов, Л.А. Химический состав молока свиноматок разного генотипа / Л.А. Рахматов, М.А. Сушенцова // Актуальные проблемы животноводства, ветеринарной медицины, переработки сельскохозяйственной продукции и товароведения. Материалы международной научно-практической конференции ФГОУ ВПО ВГАУ. – Воронеж. – 2010. – С. 65-66.
- Рахматов, Л.А. Экстерьерные особенности поросят, полученных от свиноматок различных продуктивных типов / Л.А. Рахматов, М.А. Сушенцова // Ученые записки Казанской государственной академии ветеринарной медицины им. Н.Э. Баумана. – 2013. – № 216. – С. 280-283.
- Рахматов, Л.А. Взаимосвязь и развитие поросят с молочностью и химическим составом молока свиноматок // Ученые записки Казанской государственной академии ветеринарной медицины имени Н.Э. Баумана. – 2011. – № 205. – С. 177-184.
- Роженцов, А.Л. Биохимический статус крови коров в зависимости от качества кормов / А.Л. Роженцов, С.Ю. Смоленцев // Зоотехния. – 2009. – № 12. – С. 9-10.
- Хайруллин, Д.Д. Влияние углеводно-витаминно-минерального концентрата на морфологрический состав крови дойных коров / Д.Д. Хайруллин, Ш.К. Шакиров, А.Р. Кашаева // Вестник АПК Ставрополья. – 2019. – № 4 (36). – С.36-39.
