Биохимическая характеристика генотипов галеги восточной ( Galega orientalis Lam.), произрастающей в условиях Беларуси
, произрастающей в условиях Беларуси")
Автор: Домаш В.И., Прохоров В.Н., Канделинская О.Л., Шарпио Т.П., Забрейко С.А., Грищенко Е.Р.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биологические основы кормопроизводства
Статья в выпуске: 6 т.48, 2013 года.
Бесплатный доступ
Галега восточная ( Galega orientalis Lam. ) — кормовая культура с комплексом ценных свойств, включая раннее отрастание, быстрый рост, высокую продуктивность и питательную ценность. Активная симбиотическая азотфиксация позволяет снизить нормы минерального азота, вносимого в почву, и содержание в кормах нитратов, негативно влияющих на продуктивность и качество животноводческой продукции. Ингибиторы протеиназ и лектины способны отрицательно повлиять на биологическую ценность кормов, выступая в качестве антипитательных факторов. Однако распределение и активность компонентов системы протеолиза и лектинов в органах и тканях галеги восточной остаются недостаточно изученными. Мы провели биохимические исследования листьев и семян у 19 сортов и образцов галеги (в основном, восточно-европейского происхождения) с разными сроками весеннего отрастания, оценив активность нейтральных и щелочных протеиназ, ингибиторов трипсина, фитогеагглютинирующую активность (ФГА) лектинов, содержание, состав и переваримость белков. В целом показатели у сортов значительно варьировали. Установлена высокая кормовая ценность культуры. Для галеги восточной характерно высокое содержание белка и незаменимых аминокислот. У образцов выявлены представляющие интерес показатели по сочетанию аминокислот, важных для развития молодняка, и особенности электрофоретических спектров белков семян. Показано, что для культуры характерно незначительное количество антипитательных веществ — ингибиторов трипсина (ИТ) и лектинов (активность ИТ выше при позднем сроке начала вегетации, ФГА — у образов с ранним и средним сроком).
Галега восточная, козлятник восточный, ингибиторы трипсина, лектины, белки, аминокислоты
Короткий адрес: https://sciup.org/142134822
IDR: 142134822 | УДК: 636.086.2/.3:577.112(476)
Biochemical peculiarities of Galega orientalis Lam. genotypes in Byelorussia
Galega orientalis Lam. is a fodder crop with a number of important traits, including the early beginning of vegetation, fast growth, high productivity and nutritiousness. For its cultivation, the doses of mineral N are reduced due to active symbiotic N-fixation, resulting in the decrease of nitrogen content in the fodder and livestock production. Lectins and proteinase inhibitors, as anti-nutrients, affect the fodder quality negatively. But the distribution and activity of proteolytic factors and lectins in Galega orientalis are not still clarified enough. We conducted the chemical investigation of leaves and seeds in 19 varieties and samples of Galega orientalis, originated mainly from the Eastern Europe. The activity of neutral and alkaline proteinases, tripsin inhibitors, lectin phytohemagglutination (PHA), as well as the protein content, composition and digestibility were examined. In total, the parameters varied significantly. The tested Galega varieties are the highly valuable fodder crop. Particularly, the content of proteins and essential amino acids in plants was high. The amino acid composition, necessary for the young growing animals, was revealed. In contrary, the content of tripsin inhibitors and lectins was not sufficient. The inhibitors were more active in the plants which began their vegetation later, and PHA was higher in the plants with early and intermediate beginning of vegetation. The characteristic electrophoretic patterns of seed proteins were also shown.
Текст научной статьи Биохимическая характеристика генотипов галеги восточной ( Galega orientalis Lam.), произрастающей в условиях Беларуси
Галега восточная ( Galega orientalis Lam . ) — многолетняя кормовая культура семейства Бобовые ( Fabaceae ), которая обладает комплексом ценных свойств: зимостойкостью, засухоустойчивостью, высокой эффективностью использования весенних запасов влаги; ранним отрастанием весной и быстрым ростом, значительной облиственностью (60-70 %) и стабильностью семеноводства (урожайность семян до 6 ц/га и более); долголетием (10-15 лет и более); высокой продуктивностью (за 2 укоса можно получить зеленой массы до 60-70, сена — 10-15 т/га) и питательной ценностью (в 1 корм. ед. содержится 150-270 переваримого протеина), сочетающейся с низкой себестоимостью кормовой единицы и высокой переваримостью корма. Активная симбиотическая азотфиксация позволяет снизить нормы минерального азота, вносимые в почву, и содержание в кормах нитратов, оказывающих негативное влияние на продуктивность и качество животноводческой продукции (1-3).
По основным биохимическим показателям галега восточная не уступает клеверу и люцерне. Она характеризуется высоким содержанием белка, сбалансированного по аминокислотному составу, с полным набором незаменимых аминокислот. В цветках содержатся флавоноиды, в семенах — сахароза, стахиоза, стероиды, сапонины, алкалоиды, масла и высшие жирные кислоты (пальмитиновая, стеариновая, линоленовая). В листьях обна-
Работа выполнена при финансовой поддержке ГПОФИ «Ресурсы растительного и животного мира», задание 20/3.
ружены дубильные вещества, фенолкарбоновые кислоты, витамин С, гале-гин, пеганин, рутин, каротин, танин, горькие вещества, свободные сахара, а также пектины (4-8).
Вместе с тем остаются недостаточно изученными распределение и активность в органах и тканях галеги восточной компонентов системы протеолиза и лектинов, принимающих участие в регуляции метаболизма в целом, в том числе в механизмах конститутивной и индуцированной устойчивости растений. Система протеолиза, функционирование которой обеспечивается согласованным взаимодействием ингибиторов и протеолитических ферментов, участвует в процессах обмена белков (9, 10). Следует указать и на присутствие в галеге белков лектинов (гемагглютининов), которые избирательно и обратимо взаимодействуют как с углеводами в составе полисахаридов, гликолипидов и гликопротеидов, так и со свободными моно- и олигосахаридами, включая рецепторы плазматических мембран, что обеспечивает их участие в регуляции интегральных процессов в растительных клетках (11).
Ингибиторы протеиназ и лектины способны отрицательно влиять на биологическую ценность кормовой продукции, выступая в качестве ан-типитательных факторов, что необходимо учитывать при составлении сбалансированного рациона для сельскохозяйственных животных (12). Большую ценность для исследования функционального состояния протеиназно-ингибиторной системы и метаболизма лектинов приобретают сведения, полученные при анализе электрофоретического спектра этих белков, которые позволяют выявлять маркеры высокопродуктивных и устойчивых форм в процессе селекции.
Целью настоящей работы стала биохимическая характеристика сортов и форм галеги восточной разных сроков весеннего отрастания.
Методика. Объектом исследований были сорта и образцы галеги восточной, собранные в различных эколого-географических зонах. Коллекция заложена в 2004 году на полевом стационаре РУП «НПЦ НАН Беларуси по земледелию» (г. Жодино). Почва участка — дерново-подзолистая легкосуглинистая, средне окультуренная (степень насыщенности основаниями 52,3-59,5 %), развивающаяся на легком песчанистом суглинке, подстилаемом с глубины 30-50 см рыхлым песком. Содержание гумуса в пахотном слое 2,2-2,4 %, фосфора и калия — соответственно 183-190 и 210220 мг/кг почвы, рН 6,0-6,2. Гидролитическая кислотность составляла 1,68-1,85, сумма поглощенных оснований — 10,0 мг-экв/100 г почвы.
Для биохимического анализа использовали семена и листья сортов и образцов галеги восточной, различающихся по срокам начала весеннего отрастания. Группу ранних сроков составляли сорта Widmantai, Магистр, образец № 5 (отобран из популяции сорта Гале в Минской области), Спутник, Ялгинский, Еля-ты, Горноалтайский; средних — БГСХА 3, Гале, Донецкий 90, Нестерка, образец № 2 (из популяции сорта Гале в Ленинградской области), образец № 4 (из популяции сорта Гале на территории Центрального ботанического сада НАН Беларуси), Полесская, Салют; поздних — Нальчик 2, Нальчик 3, Кавказ. Кроме того, в группу позднеспелых был включен образец галеги лекарственной к-24864, полученный из коллекции Всероссийского НИИ растениеводства им. Н.И. Вавилова (ВИР, г. Санкт-Петербург).
Активность белков — ингибиторов трипсина определяли по уменьшению скорости гидролиза субстрата ферментом (трипсином) в их присутствии (13). В качестве субстрата использовали БАПА (На-бензоил-ПЕ-аргинин-4-нитроанилид). Активность нейтральных протеиназ оценивали по методу Ансона (14). За единицу активности принимали количество фермента, вызывающего образование 1 мкмоль тирозина за 1 мин инкубации. Активность БАПАазы определяли по методу Эрлангера (15).
Общую фракцию лектинов выделяли посредством экстракции этанолом (16). Тестирование полученных препаратов лектинов выполняли с помощью реакции агглютинации эритроцитов человека I(0), Rh + (17). Фи-тогемагглютинирующую активность (ФГА) лектинов определяли посредством микротитрования на иммунологических планшетах с U-образными лунками с последующим добавлением в них 2,5 % суспензии эритроцитов. Реакцию проводили при комнатной температуре, гемагглютинацию регистрировали через 2 ч после начала титрования. Активность лектинов выражали в величинах, обратных минимальной концентрации белка (мкг белка/мл), при которой отмечали реакцию гемагглютинации, и переводили в Ед/г сырой массы.
Переваримость белка оценивали in vitro с использованием фермента панкреатина (18). Содержание белка определяли по методу Кьельдаля и методу Бредфорда (19). В последнем случае в качестве стандарта служил бычий сывороточный альбумин. Электрофорез белков из семян галеги проводили на приборе S-250 («Amercham Bioscience», Великобритания). Белки экстрагировали буфером Трис-Hd (рН 7,0), содержащим сахарозу, EDTA и 0,01 % аскорбат. Электрофорез осуществляли в 15 % ПААГ в денатурирующих условиях по методу U.K. Laemmli (20). В качестве метчиков использовали в -галактозидазу (молекулярная масса 116 кД), альбумин бычий (66 кД), овальбумин (45 кД), лактатдегидрогеназу (35 кД), РНКазу (25 кД), в -лактатглобулин (18,4 кД), лизоцим (14,4 кД). Гели фиксировали 20 % ТХУ. Электрофореграммы обрабатывали с использованием компьютерной программы LAB, Control Centre v. 2.1. Аминокислотный состав белков определяли на автоматическом анализаторе Т339 (Чехия).
Опыты проводили в 2 биологических и 3-4 аналитических повторностях. Статистическую обработку данных осуществляли с использованием компьютерной программы Microsoft Excel.
Результаты. Активность нейтральных протеиназ (в расчете на абсолютно сухую массу) у исследуемых сортов и форм находилась в пределах
1. Активность протеиназ и ингибиторов трипсина в листьях у галеги восточной Galega orientalis Lam. разных сроков весеннего отрастания в фазу цветения ( X ± х , полевой стационар РУП «НПЦ НАН Беларуси по земледелию», г. Жо-дино, 2009-2010 годы)
|
Сорт, образец |
Нейтральные протеиназы, ЕА/г |
Щелочная протеиназа (БАПАаза), ЕА/г |
Ингибиторы трипсина, ИЕ/г |
|||
|
с.м. |
абс.с.м |
с.м. |
абс.с.м |
с.м. |
абс.с.м |
|
|
Образец № 5 |
5,62±0,00 |
21,86±0,12 |
Р а н н и е 18,52±0,01 |
72,04±0,38 |
1,07±0,00 |
4,16±0,00 |
|
Спутник |
5,96±0,00 |
24,22±0,00 |
20,84±0,01 |
84,68±0,25 |
0,85±0,00 |
3,48±0,00 |
|
Widmantai |
7,46±0,00 |
25,15±0,13 |
18,60±0,00 |
72,20±0,00 |
1,04±0,00 |
4,05±0,00 |
|
Магистр |
7,50±0,00 |
30,12±0,00 |
21,36±0,00 |
85,44±0,17 |
0,84±0,00 |
3,36±0,00 |
|
Донецкий 90 |
5,45±0,00 |
25,70±0,17 |
С р е д н и е 18,24±0,01 |
86,00±0,41 |
1,34±0,00 |
5,32±0,00 |
|
БГСХА 3 |
5,47±0,00 |
25,79±0,15 |
21,60±0,00 |
93,42±0,21 |
0,73±0,00 |
3,17±0,00 |
|
Образец № 2 |
5,93±0,00 |
22,54±0,12 |
18,68±0,00 |
71,00±0,22 |
1,34±0,00 |
5,09±0,00 |
|
Нестерка |
6,58±0,00 |
27,42±0,01 |
19,54±0,00 |
81,42±0,18 |
0,74±0,00 |
3,10±0,00 |
|
Гале |
7,34±0,00 |
28,68±0,20 |
21,00±0,00 |
82,06±0,17 |
0,73±0,00 |
2,86±0,01 |
|
Салют |
7,65±0,00 |
30,38±0,00 |
21,60±0,00 |
93,42±0,00 |
0,82±0,01 |
3,26±0,00 |
|
Нальчик 2 |
6,68±0,00 |
26,00±0,22 |
П о з д н и е 18,72±0,00 |
72,87±0,00 |
1,07±0,00 |
4,16±0,00 |
|
Кавказ |
5,22±0,01 |
25,82±0,32 |
18,00±0,15 |
88,99±0,75 |
1,08±0,01 |
5,36±0,00 |
|
к-24864 |
6,58±0,00 |
25,54±0,05 |
18,06±0,19 |
70,16±0,19 |
1,34±0,00 |
5,19±0,00 |
|
Примечание. БАПА - |
- N a -бензоил-DL-аргинин-4-нитроанилид, |
с.м. - сырая |
масса, абс.с.м. - |
|||
|
абсолютно сухая масса. |
||||||
21,86-30,38 ЕА/г, щелочной протеиназы — 70,16-93,42 ЕА/г, ингибиторов трипсина — 2,9-5,3 ИЕ/г (табл. 1). При этом наибольшие значения активности нейтральных и щелочных протеиназ были характерны для растений средних сроков весеннего отрастания, ингибиторов трипсина — для представителей группы поздних сроков. Тесная корреляционная связь была обнаружена между активностью нейтральных протеиназ и ингибито ров трипсина в листьях у галеги среднего срока начала весеннего отраста ния (r = -0,71), а также между активностью БАПАазы и ингибиторов трипсина (r = -0,85). Содержание ингибиторов трипсина в листьях оказалось незначительным и составляло 0,3-0,5 мг/г абсолютно сухой массы, то есть ниже предельно допустимого количества (5 мг/г абсолютно сухой массы), что свидетельствует о высокой кормовой ценности этой культуры.
Известно, что протеолитические ферменты играют важную роль не только в обмене белков в прорастающих семенах, но и в реутилизации азотистых соединений из вегетативных органов в репродуктивные (21). Белки-ингибиторы протеиназ — биологически активные вещества, образующие с ферментами неактивные (обратимые) комплексы, в которых ферменты лишены каталитической активности. Эти белки выполняют функции регуляторов активности протеолитических ферментов, а также принимают участие в формировании защитных механизмов при действии фи топатогенов и насекомых-вредителей (22). Активность ингибиторов трипсина в зрелых семенах галеги разных сроков вегетации составляла около 7 ИЕ (0,65 мг/г абсолютно сухой массы).
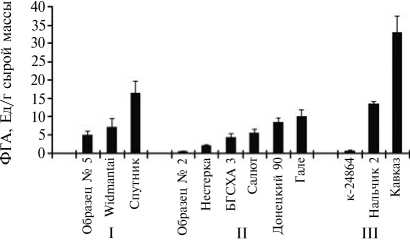
Наши исследования продемонстрировали наличие в листьях галеги восточной фитогемагглютинирующей активности (ФГА) (рис. 1). Величина ФГА лектинов в листьях у изученных образцов значительно варьировала. Установлено, что у форм с ранним сроком начала весеннего отрастания показатель ФГА (Ед/г сырой массы) находился в пределах от 4,9 (образец № 5) до 16,4 (сорт Спутник), у среднеспелых сортообразцов — от 0,4 (образец № 2) до 9,9 (сорт Гале), у представителей поздних сроков вегетации — изменялся от 0,8 (сортообразец К-24864) до 3 3,0 (сорт Кавказ).
В семенах активность лектинов варьировала от 149,2 (сорт Ялгин-ский) до 297,0 Ед/г сырой массы (сорт Магистр) у представителей ранних сроков весеннего отрастания и от 168,0 (сорт Горноалтайский) до 320,0 Ед/г сырой массы (сортообразец № 2) у генотипов средних сроков. По-види- мому, аккумуляция лектинов в галеге восточной осуществляется в боль шей степени в семенах, как это свойственно бобовым растениям (17).
Сорт
Рис. 1. Фитогемагглютинирующая активность (ФГА) лектинов в листьях у сортов галеги восточной Galega orientalis Lam. с ранним (I) , средним (II) и поздним (III) сроком весеннего отрастания (РУП «НПЦ НАН Беларуси по земледелию», г. Жодино, 2009-2010 годы).
Максимальные значения фитогемагглютинирующей активности лектинов в листьях отмечали у растений ранних и средних сроков вегетации, однако значения этого показателя были невысокими. В доступной литературе мы не обнаружили данных о предельно допустимых количествах лектинов и их активности в кормах для сельскохозяйственных животных, поэтому делать какие-либо выводы об антипитально-сти лектинов галеги преждевременно.
В листьях галеги восточной содержалось от 15,37 (сорт Гале) до 19,79 % (образец № 5) белка. Показатель переваримости белка in vitro варьировал от 43 до 55 % (табл. 2).
Содержание белка у образцов галеги ранних сроков отрастания составляло в среднем 18,1 %, у представителей группы со средним сроком — 16,4 %; переваримость белка — соответственно 55,1 и 48,1 %. В 1 кг сухой массы у образцов галеги с ранним сроком отрастания содержалось 91 г, со средним сроком — 79 г переваримого белка, что согласуется с данными других авторов (2).
-
2. Содержание белка в листьях у образцов галеги восточной Galega orientalis Lam. разных сроков весеннего отрастания в фазу цветения и его переваримость ( X ± х , РУП «НПЦ НАН Беларуси по земледелию», г. Жодино, 20092010 годы)
-
3. Аминокислотный состав (мг/100 мг белка) в листьях у галеги восточной Galega orientalis Lam. (РУП «НПЦ НАН Беларуси по земледелию», г. Жодино, 2009-2010 годы)
Сорт, образец Содержание белка в расчете на абсолютно сухую массу, % | Переваримость белка, %
Р а н н и е
|
Магистр |
17,10±0,01 |
47,37±0,34 |
|
Спутник |
18,90±0,01 |
52,44±0,69 |
|
Widmantai |
16,50±0,01 |
47,03±0,16 |
|
Образец № 5 |
19,79±0,01 |
55,27±0,26 |
|
С р е д н и е |
||
|
Гале |
15,37±0,01 |
50,00±0,58 |
|
БГСХА 3 |
15,51±0,01 |
49,59±0,28 |
|
Нестерка |
17,20±0,16 |
43,61±0,78 |
|
к-24864 |
15,42±0,01 |
46,91±0,26 |
|
Салют |
17,39±0,01 |
49,16±0,34 |
|
Донецкий 90 |
17,68±0,16 |
49,23±0,42 |
У исследуемых образцов галеги было установлено высокое содержание незаменимых аминокислот (табл. 3).
|
Аминокислота |
Сорт, образец |
|
№ 5 Widmantai Спутник | Нестерка | Гале Донецкий Салют | БГСХА к-24864 |
Незаменимые аминокислоты:
|
Вал |
5,13 |
4,89 |
4,95 |
4,94 |
4,90 |
5,02 |
5,05 |
4,84 |
4,98 |
|
Иле |
3,85 |
3,62 |
3,69 |
3,65 |
3,56 |
3,68 |
3,78 |
3,62 |
3,71 |
|
Лей |
8,56 |
8,23 |
8,47 |
8,39 |
8,17 |
8,61 |
8,50 |
8,28 |
8,54 |
|
Лиз |
5,79 |
6,23 |
5,97 |
6,16 |
5,90 |
6,05 |
6,14 |
6,20 |
6,04 |
|
Мет |
1,14 |
1,03 |
1,10 |
1,09 |
1,04 |
0,94 |
1,07 |
1,12 |
1,14 |
|
Тре |
5,20 |
4,69 |
4,72 |
4,85 |
4,70 |
4,78 |
4,56 |
4,52 |
4,89 |
|
Фен |
5,59 |
5,96 |
5,71 |
5,66 |
5,56 |
5,65 |
5,62 |
5,91 |
5,79 |
|
Заменимые аминокислоты: Глу 2,42 12,00 |
12,13 |
12,35 |
12,08 |
12,34 |
12,34 |
11,83 |
12,58 |
||
|
Арг |
5,76 |
5,89 |
5,91 |
5,94 |
5,65 |
5,74 |
5,59 |
5,57 |
6,07 |
|
Сер |
5,14 |
4,93 |
5,14 |
5,03 |
5,30 |
5,14 |
4,90 |
5,15 |
5,01 |
|
Про |
6,60 |
5,54 |
6,58 |
6,21 |
6,83 |
6,13 |
7,02 |
6,65 |
5,69 |
|
Цис |
0,51 |
0,72 |
0,74 |
0,74 |
0,62 |
0,61 |
0,74 |
0,65 |
0,76 |
|
Гли |
5,06 |
5,19 |
5,23 |
5,15 |
5,23 |
5,28 |
5,16 |
4,86 |
5,17 |
|
Ала |
6,27 |
6,27 |
6,03 |
6,20 |
5,97 |
6,11 |
6,04 |
5,63 |
6,20 |
|
Тир |
5,15 |
5,71 |
5,01 |
5,31 |
5,22 |
5,46 |
5,02 |
5,29 |
5,28 |
|
Гис |
3,01 |
3,56 |
2,99 |
3,32 |
3,02 |
3,09 |
3,17 |
3,39 |
3,03 |
|
Сумма: общая |
95,33 |
94,91 |
95,00 |
95,16 |
95,03 |
94,94 |
94,95 |
94,37 |
95,06 |
|
незаменимых аминокислот |
35,26 |
34,65 |
34,61 |
34,74 |
33,83 |
34,73 |
34,72 |
34,49 |
35,09 |
|
Арг+ Гис |
8,77 |
9,45 |
8,90 |
9,26 |
8,67 |
8,83 |
8,76 |
8,96 |
9,10 |
|
Мет + Цис |
1,65 |
1,75 |
1,84 |
1,83 |
1,66 |
1,55 |
1,81 |
1,77 |
1,90 |
|
Фен + Тир |
10,74 |
11,67 |
10,72 |
10,79 |
10,78 |
11,11 |
10,64 |
11,20 |
11,07 |
В группах с ранним и средним сроком весеннего отрастания сумма незаменимых аминокислот варьировала незначительно. Заслуживает внимания вариабельность содержания аминокислот пролина и цистеина, принимающих участие в формировании неспецифической устойчивости рас- тений. Так, наиболее высокие значения этого показателя отмечались у сортов Гале и Салют со средним сроком начала вегетации. Представляются интересными интегральные показатели суммы аминокислот аргинина и гистидина, серосодержащих аминокислот метионина и цистеина, фенилаланина и тирозина, имеющих значение для молодняка сельскохозяйственных животных (23). Сорт Widmantai характеризовался наибольшими значениями суммы по аргинину и гистидину, а также фенилаланину и тирозину.
Электрофоретический спектр белков семян у галеги восточной был представлен 18-26 компонентами, основная часть которых оказалась сосредоточенной в области молекулярных масс 35-124 кДа. Сорт Нальчик 2 характеризовался отсутствием ярко выраженного полипептида в области
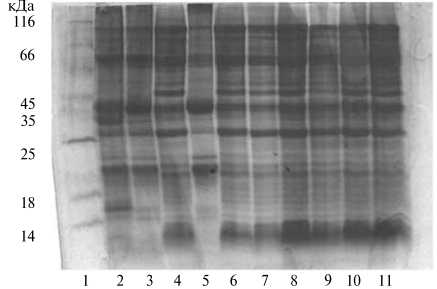
Рис. 2. Электрофоретический спектр белков семян у галеги восточной Galega orientalis Lam. с разным сроком весеннего отрастания: 1 — белки-маркеры; 2 — Нальчик 3; 3 — Нальчик 2; 4 — Полесская; 5 — Нестерка; 6 — Донецкий 90; 7 — Ялтинский; 8 — образец № 4; 9 — Горноалтайский; 10 — Магистр; 11 — Еля-ты (РУП «НПЦ НАН Беларуси по земледелию», г. Жодино, 2009-2010 годы).
14 кДа, сорт Нестерка — отсутствием полипептидов в области 14-25 и 35 кДа. Отмеченные у галеги особенности электрофоретического спектра белков семян могут быть использованы в селекции этой важной кормовой культуры.
Таким образом, у различных сортов и образцов галеги восточной листья характеризуются высоким содержа- нием белка, в состав которого входят все незаменимые аминокислоты. По набору аминокислот у исследованных форм не наблюдается высокой внутривидовой вариабельности. Не выявлены особенности ами нокислотного состава белка в связи со сроками весеннего отрастания. Из образцов с ранним сроком наиболее перспективными по содержанию белка оказались сорт Спутник и образец № 5, со средним — сорта Салют и Донецкий 90. Культура содержит незначительное количество антипитательных белков — ингибиторов трипсина и лектинов.
Л И Т Е Р А Т У Р А
-
1. Л а м а н НА., П р о х о р о в В.Н., М о р о з о в а И.М., С а к о в с к а я А.Г. Галега восточная — многолетняя высокобелковая кормовая культура: биологические особенности, технология возделывания, хозяйственное использование. Минск, 2008.
-
2. Я р о ш е в и ч М.И., К у х а р е в а Л.В., Б о р е й ш а Н.С. Галега восточная — перспективная кормовая культура. Биология, кормовая ценность, требования к условиям произрастания, особенности возделывания. Минск, 1991.
-
3. К ш н и к а т к и н а А.Н., Г у щ и н а В.А., Г а л и у л л и н А.А. Продукционный процесс агрофитоценоза козлятника восточного в условиях Среднего Поволжья. Сельскохозяйственная биология, 2003, 2: 101-107.
-
4. С т р е л ь ц и н а С.А., Ю д к е в и ч Е.В., Ж у к о в а М.А, К о н а р е в А.В., Д з юб е н к о Н.И. Биохимическая характеристика козлятника восточного ( Galega orientalis Lam). Аграрная Россия, 2006, 6: 46-52.
-
5. Т о к а р ь Н.А. Галега восточная. В сб.: Новые и малораспространенные кормосилосные растения. Киев, 1969: 149-152.
-
6. 3 е в а х и н а Ю.А., О ф и ц е р о в Е.Н. Сравнительное содержание пектиновых ве
ществ в листьях и стеблях Galega orientalis. Химия растительного сырья, 2003, 2: 33-38.
-
7. С т р е л ь ц и н а С.А., Ж у к о в а М.А., Ч а ч к о Е.В., Д з ю б е н к о Н.И., К он а р е в А.В. Сравнительный анализ внутрипопуляционной изменчивости люцерны посевной Medicago sativa L. и козлятника восточного Galega orientalis L. по биохимическим
признакам качества. Сельскохозяйственная биология, 2001, 5: 37-47.
-
8. С а г и р о в а Р.А. Онтогенетический морфогенез галеги восточной ( Galega orientalis Lam.) как перспективного кормового растения. Сельскохозяйственная биология, 2009, 4: 75-80.
-
9. М о с о л о в В.В., Г р и г о р ь е в а Л.И., В а л у е в а Т.А. Участие протеолитических ферментов и их ингибиторов в защите растений. Прикладная биохимия и микробиология, 2001, 37(7): 131-140.
-
10. Д о м а ш В.И., Ш а р п и о Т.П., 3 а б р е й к о С.А., С о с н о в с к а я Т.Ф. Протеолитические ферменты и ингибиторы трипсина высших растений в условиях стресса. Биоорганическая химия, 2008, 34(3): 353-357.
-
11. Ш а к и р о в а Ф.М., Б е з р у к о в а М.В. Современные представления о предполагаемых функциях лектинов растений. Журнал общей биологии, 2007, 68(2): 109-125.
-
12. П о н о м а р е н к о Ю.А. Питательные и антипитательные вещества в кормах. Минск, 2007.
-
13. Г о ф м а н Ю.Я., В а й с б л а й И.М. Определение активности ингибиторов трипсина в семенах гороха. Прикладная биохимия и микробиологя, 1975, 11(5): 777-787.
-
14. A n s o n M.Z. The estimation of pepsin, trypsin, papain and catepsin with hemoglobin. J. Genet. Physiol., 1938, 22: 79-89.
-
15. E r l a n g e r F., K o k o w s k y N., C o h e n W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys., 1961, 96: 271-278.
-
16. С ы т н и к о в Д.М., К о ц ь С.Я., М а л и ч е н к о С.М. Лектиновая активность различных органов сои в условиях эффективного и неэффективного симбиоза. Физиология и биохимия культурных растений, 2006, 38, 1: 53-60.
-
17. Б а б о ш а А.В., Л а д ы г и н а М.Е. Определение фитогемагглютининов в связи с вирусоустойчивостью картофеля. В кн.: Физиолого-биохимические и биофизические методы диагностики степени устойчивости растений к патогенам и другим факторам /Под ред. М.Е. Ладыгиной. М., 1992: 43-52.
-
18. Л е в и ц к и й А.П., П о х о м е н к о Л.И., В о в ч у к С.В. Биохимический метод определения переваримости зерновых белков: Метод. реком. Одесса, 1985.
-
19. B r a d f o r d M.M. Rapid and sensitive method for quantitation of microgramm quantities of protein utilizing the principle of protein dye binding. Anal. Biochem., 1976, 8: 248-254.
-
20. L a e m m l i U.K. Cleavage of structure protein during the assembly of the head of bacteriophage. Nature, 1970, 227(5259): 680-685.
-
21. Л о к ш и н а Л.А. Регуляторная роль протеолитических ферментов. Молекул. биология, 1979, 13(6): 1205-1229.
-
22. М о с о л о в В.В., В а л у е в а Т.А. Ингибиторы протеиназ и их функции у растений (обзор). Прикладная биохимия и микробиология, 2005, 41(3): 261-283.
-
23. Л е н и н д ж е р А. Основы биохимии. М., 1985, т. 2: 653-681.
ГНУ Институт экспериментальной ботаники Поступила в редакцию им. В.Ф. Купревича НАН Беларуси, 6 февраля 2012 года
BIOCHEMICAL PECULIARITIES OF Galega orientalis Lam. GENOTYPES IN BELORUSSIA
V.I. Domash, V.N. Prokhorov, O.L. Kandelinskaya, T.P. Sharpio, S.A. Zabreiko, E.R. Grischenko
