Биология сои: возможности оптимизации отдельных продукционных процессов

Бесплатный доступ
С целью увеличения зерновой продуктивности сои следует продолжить изучение биологии культуры и её биологических особенностей. К таким особенностям относятся фотопериодические реакции, продолжительность и интенсивность прохождения этапов органогенеза и эффективность симбиотической азотфиксации. Изучали сорта амурской селекции МК 1, Амурская 310, Янтарная, ВНИИС 1, Смена (выведенные и частично используемые в период с 1975 по 2005 гг.). Относительно высокая продуктивность фотосинтеза отмечена у сортов: Мк 1 (5,2 г/см2), ВНИИС 2 (4,78 г/см ), Янтарная (3,46 г/см2). Максимальный показатель получен для дикорастущей сои (7,10 г/см2). В ряде случаев выявлены несоответствия масштабов нарастания вегетативной массы и урожайностью растений. Объясняется это сортовыми особенностями перераспределения пластических веществ между органами растений в различные фазы развития. Количество ассимилятов в расчете на 1 боб в течение фазы налива бобов изменяется от 0,64 до 10,0 мг. Во второй половине вегетации наблюдается явление абортивности цветков и бобов. В значительной мере это обусловлено дефицитом питательных веществ. У сортов Смена, ВНИИС 1, Амурская 310 количество почек сформировавших бобы составляет соответственно 39, 32, 31 % (от общего количества). У дикорастущей сои этот же показатель превышает 67 %. Деятельность симбиотической системы в первой половине вегетационного периода (количество и масса клубеньков) у сортов МК 1 и Смена, различающихся по периоду вегетации более чем на 7 дней, оказалась практически одинаковой. В изучаемых почвах отмечен высокий уровень соответствия (детерминированности) в системе «сорт-штамм». Затраты на формирование клубеньков составляют от 1,8 до 2,3 % от надземной массы в фазе налива бобов. В течение отдельных этапов органогенеза и фаз развития сои имеет место «неэкономичное» использование ассимилятов, обостряемое явлениями абортивности.
Соя, биология, пластические вещества, органогенез, ризобии, азотфиксация, урожай
Короткий адрес: https://sciup.org/142151321
IDR: 142151321 | УДК: 633.853.52:581.1
Biology of soybean: possibilities to optimize the separate production processes
To enlarge seed productivity of soybean it is necessary to prolong studying of this crop biology and its biological features. Such features are: photoperiodic reactions, duration and intensiveness of organogenesis stages, effectiveness of symbiotic nitrogen fixing. The cultivars of Amur breeding were studied: MK 1, Amurskaya 310, Yantarnaya, VNIIS 1, Smena (developed and partially used in 1975-2005). The relatively high productivity of photosynthesis was recorded at cultivars: МК 1 (5.2 g per cm2), VNIIS 1 (4.78 g per cm2), Yantarnaya (3.46 g per cm2). The maximal meaning was noted at wild soybean (7.10 g per cm2). In some cases there were observed discrepancies between level of vegetative mass growth and plant yield. It is explained by features of a cultivar on redistribution of plastic substances between plant organs in the different phases of its development. Number of assimilates per a bean during the phase of pods formation varies from 0.64 to 10.0 mg. A phenomenon of flowers and beans abortion is observed at the second half of vegetative period. This is caused considerably by a deficit of nutrients. Number of buds forming pods was 39, 32, 31% (of their total number), respectively, for cultivars Smena, VNIIS 1, Amurskaya 310. This trait of wild soybean exceeds 67%. Activity of symbiotic system in the first half of vegetative period (quantity and weight of tubercles) of cultivars MK 1 and Smena (which are differed with vegetative period duration more than seven days) was almost equal. In studying soils a high level of correspondence (determinacy) was noted in system cultivar-strain. Expenses for tubercles formation are from 1.8 to 2.3% of aboveground mass in phase of pods ripening. ‘Uneconomic’ utilization of assimilates aggravated by abortion is observed during the some stages of organogenesis and development phases.
Текст научной статьи Биология сои: возможности оптимизации отдельных продукционных процессов
To enlarge seed productivity of soybean it is necessary to prolong studying of this crop biology and its biological features. Such features are: photoperiodic reactions, duration and intensiveness of organogenesis stages, effectiveness of symbiotic nitrogen fixing. The cultivars of Amur breeding were studied: MK 1, Amurskaya 310, Yantarnaya, VNIIS 1, Smena (developed and partially used in 1975–2005). The relatively high productivity of photosynthesis was recorded at cultivars: МК 1 (5.2 g per сm2), VNIIS 1 (4.78 g per cm2), Yantarnaya (3.46 g per сm2). The maximal meaning was noted at wild soybean (7.10 g per cm2). In some cases there were observed discrepancies between level of vegetative mass growth and plant yield. It is explained by features of a cultivar on redistribution of plastic substances between plant organs in the different phases of its development. Number of assimilates per a bean during the phase of pods formation varies from 0.64 to 10.0 mg. A phenomenon of flowers and beans abortion is observed at the second half of vegetative period. This is caused considerably by a deficit of nutrients. Number of buds forming pods was 39, 32, 31% (of their total number), respectively, for cultivars Smena, VNIIS 1, Amurskaya 310. This trait of wild soybean exceeds 67%. Activity of symbiotic system in the first half of vegetative period (quantity and weight of tubercles) of cultivars MK 1 and Smena (which are differed with vegetative period duration more than seven days) was almost equal. In studying soils a high level of correspondence (determinacy) was noted in system cultivar-strain. Expenses for tubercles formation are from 1.8 to 2.3% of aboveground mass in phase of pods ripening. ‘Uneconomic’ utilization of assimilates aggravated by abortion is observed during the some stages of organogenesis and development phases.
В мировом сельскохозяйственном производстве соя является одной из основных бобовых культур, с помощью которой решена проблема производства полноценного растительного белка. В России основные центры соеводства сложились на Дальнем Востоке, на Северном Кавказе и в Белгородской области. Значительное внимание уделяется этой культуре в Центральной России, в Поволжье и в южной Сибири.
На указанных территориях испытываются и возделываются сотни полиморфных сортов, обладающих достаточно разнообразными хозяйственно ценными свойствами. Условия развития соеводства в настоящее время во многом определяются агротехнологическими факторами мезо - и микрозон.
Увеличение хозяйственной потребности в продуктах соеводства предполагает расширение посевов сои в стране до 6 млн га [7; 15]. В дальнейшем, в зависимости от ситуации, можно ожидать расширение посевов до 12–14 млн га, при условии освоения ранее не используемых земель и замены соевыми бобами малопродуктивных культур. Это , в свою очередь , потребует создания дополнитель ного набора сортов, адаптированных к условиям вновь осваиваемых под указанную культуру территорий. В системном порядке продолжится работа по увеличению продуктивности соевых полей.
Вместе с тем в процессе многолетнего селекционно-генетического совершенствования растений сои, как важнейшего объекта мирового сельского хозяйства, урожайность культуры приблизилась к своему биолого-ценотическому порогу. Преодоление трудностей с ростом воспроизводительных свойств растений непосредственно зависит от уровня изученности биологии культуры и возможностей комбинации физиологобиохимических реакций и фенотипических проявлений.
В этом отношении представляет интерес повышение «экономичности» использования растениями энергетических и пластических соединений на формирование систем развития и жизнеобеспечения. Биологические особенности сои предопределяются (помимо гидротермических ресурсов) условиями освещённости (фотопериодизм), оптимальными условиями прохождения этапов органогенеза и стабильным режимом азотного питания. Последний фактор, необходимый для биосинтеза высокобелковых соединений, в определённой мере регулируется самим растением за счет симбиотических взаимоотношений. Затраты на образование клубеньков, поддержание симбиоза на максимально выгодном для растений уровне, потребление за счет этого атмосферного азота осуществляются растениями в течение всего вегетационного периода.
Начинаются указанные процессы после набухания семян, когда реализуется генетическая программа использования запасных веществ. При разрыве семенной оболочки и при поступлении кислорода усиливаются процессы дыхания. В этот период прорастает первичный корень и начинается движение семядолей вверх. В семенах осуществляются сложные биохимические процессы усвоения запасных веществ. Полное использование углеводов отмечается на 3 - й день прорастания, потребление митохондриями кислорода увеличивается, в связи с перестройкой субстрата и в целом запасного энергетического материала. На 4 - й день возрастает, а затем резко снижается активность липооксидазы, которая способствует быстрой деградации жиров (Mc Alister et at.,
1951; Kamata, 1952; Switzer et at., 1957, Howell, 1961) [ цит . по : Хауэл Р . В .] [16]. На 5 - й день после прорастания потребление кислорода на единицу азота заметно возрастает ( Staniforth, 1956) [ цит . по : Хауэл Р . В .] [16]. Большинство указанных реакций происходят с участием митохондриальных ферментов (липооксидазы, пероксидазы, каталазы) и других соединений, соответствующих ферментативному процессу глиоксалатного цикла. Темпы использования белков и масла имеют сходную динамику. В прорастающих семенах появляются аскорбиновая кислота, тиамин, рибофлавин, цистин, цистеин и тиамин ( T. Sugavara, 1953) [ цит . по : Хауэл Р . В .] [16]. По данным Пе-тибской В.С. [12], кратковременно в семядолях имеет место трипсинингиби-рующая активность. Появлению ряда соединений способствует освещённость при выходе семядолей на дневную поверхность. Для использования запасных веществ необходимы динамические структуры перемещения веществ и энергетический материал. По - видимому, в данном случае постоянно осуществляются реакции переаминирования.
Указанные процессы осуществляются в рамках I- го и II- го этапов органогенеза [6; 15] по трёхзначному коду ВВСН – 0,01 –0,09. Из конуса нарастания почки после выхода семядолей на поверхность закладываются вегетативные элементы растения. В этот же отрезок времени в позеленевших семядолях и в разворачивающихся листьях усиливается фотосинтез. Завершаются указанные этапы органогенеза появлением одного - двух настоящих листьев [9]. Характерно, что большую часть рассматриваемого периода для всех процессов основным источником энергии и материалов для конституционного строительства являются семядоли, содержащие белки, молекулы масла, нуклеиновые кислоты, фосфолипиды и другие соединения .
На 5 – 8- й день прорастания семян на корнях образуются визуально наблюдаемые клубеньки.
Следовательно, весь анализируемый материал свидетельствует о том, что в 80
первую неделю роста, в условиях высокой нуждаемости в факторах внешней среды, растения тратят часть биологических ресурсов для создания предпосылок формирования симбиотического аппарата. Даже в этих условиях корни уже выделяют соединения, привлекающие почвенные микроорганизмы.
В дальнейшем , вне зависимости от ресурсного обеспечения (и особенно при недостатке в почве подвижных соединений азота) , соя в течение большей части вегетационного периода затрачивает пластические вещества для образования клубеньков при наличии в почве комплиментарных штаммов клубеньковых бактерий.
У сои ген, способствующий образованию клубеньков, обозначается как No (n о) [4]. Кроме того , выделен ряд мендели-рующих генов, контролирующих количество клубеньков, сорто - штаммовую детерминированность, реакцию на нитраты ( Caetono-Anoies, Zresshof, 1991) [ цит . по : Вэнс К. ] [3].
Уже в начале прорастания корни бобовых выделяют сигнальные молекулы, на которые реагируют ризобии. В состав корневых выделений входят важные для сигналинга соединения [14]. Изофлава-ноиды (диадзеин, генистеин, глицетин) стимулируют активизацию Nod- фактора ризобий. Взаимодействуя с клубеньковыми бактериями, соя изначально затрачивает определённые биологические ресурсы на микробно - растительный контакт.
В течение вегетационного периода такое взаимодействие усиливается и выражается в количестве и массе клубеньков, а также , опосредованно, в интенсивности продукционных процессов.
При возделывании сои на обширных территориях во времени происходит взаимная адаптация партнеров симбиоза и осуществляется масштабная симбиотическая азотфиксация. Названные явления проанализированы в соесеющих регионах Дальнего Востока, где посевы сои приблизились к 1 млн га. Такая ситуация способствует коэволюции [5] растения-хозяина и микросимбионта, учитывая широкое распространение в изучаемом регионе диких форм сои и аборигенных специфичных ризобий.
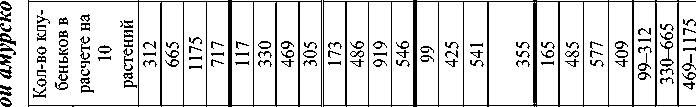
Исследования, проводившиеся Лопаткиной Э.Ф . [7; 8; 9] на лугово - чернозёмо видных почвах Приамурья с амурскими сортами сои (второго–четвёртого этапов селекционных работ) , достаточно отсе-лектированными за многие годы возделывания (1975–2005 гг.), продемонстри ровали значительную изменчивость признаков симбиотрофии.
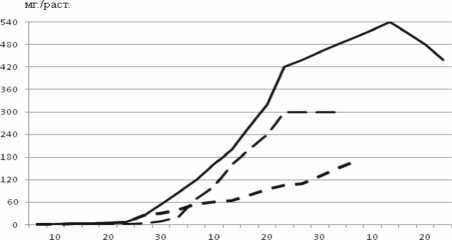
Установлено, что после первого контакта растения - хозяина и клубеньковых бактерий на всех этапах органогенеза (до бобообразования) количество клубеньков возрастает. Максимальные показатели их числа и массы (в расчете на 1 растение) во времени не совпадают (рис. 1).

Количество клубней на корнях сои
■ общее количество ^— 'Набоковых корнях * • на главном корне
а
Количество клубней на корнях сои общая масса — ■ на главном корне — • на боковых корнях
б
Рисунок 1 – Динамика количества и массы клубеньков у сои, сорт Амурская 310 : а – шт./раст.; б – мг/раст.
Доминирующее значение для азотфик-сации имеют, по - видимому, клубеньки, образовавшиеся на боковых корнях. К фазе налива бобов количество клубеньков уменьшается вследствие приостановления роста корневой системы. В это время нарастание вегетативной массы достигает своего максимума (рис. 2, 3). Обращает на себя внимание существенная аналогия темпов прироста массы клубеньков и вегетативной части растений.
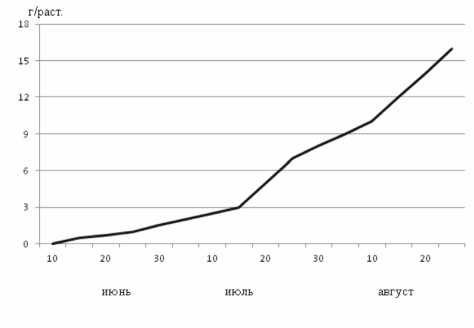
Рисунок 2 – Динамика нарастания надземной части сои, сорт Амурская 310

июнь июль август
Рисунок 3 – Динамика нарастания корневой системы сои, сорт Амурская 3 10
По данным Лопаткиной Э.Ф. [7] , развитие надземной вегетативной массы у изучаемых сортов сои по показателям площади листьев различаются в 1,5–2 ра за (табл. 1).
Таблица 1
Площадь листьев и продуктивность фотосинтеза у сортов сои амурской селекции [9]
|
Сорт |
Площадь листьев в период окончание цветения – начало образования бобов, см 2 |
Продуктивность фотосинтеза, г/см 2 |
|
МК 1 |
3949 + 630 |
5,20 |
|
Амурская 310 |
2818 + 188 |
3,00 |
|
Янтарная |
3565 + 291 |
3,46 |
|
ВНИИС 2 |
2119 + 152 |
4,8 |
|
Смена |
3571 + 305 |
- |
|
Glycine ussuriensis |
18161 + 99 |
7,10 |
В этой связи следует ожидать большие различия у растений в снабжении продуктами фотосинтеза. Но многократно отмечено, что продуктивность фотосинтеза часто не соответствует площади листьев [2]. Большая часть ассимилятов тратится на построение конституционной структуры и поддержание жизнедеятельности растительного организма. Следовательно, в зависимости от сорта функционирование листовой поверхности часто оказывается «экономически» нецелесообразным с точки зрения получения конечного продукта (семян). Так , сорта МК 1 и Янтарная, при близких значениях площади листьев, по продуктивности фотосинтеза различались почти в 1,5 раза. Но самый высокий показатель этого признака свойственен дикорастущей сое ( Glycine ussuriensis ) при минимальном значении площади листовой поверхности.
Во многом указанные закономерности объясняются сортовыми различиями и определяются особенностями использования ассимилятов по фазам развития. До VII этапа органогенеза идёт формирование мужского и женского гаметофитов и затем развитие всех элементов цветка в фазе бутонизации. Образование репродуктивных морфоэлементов (генеративных точек) в этот период мало зависит от продуктов ассимиляции, хотя и контролируется внешними условиями. Поступающие в органы растений ассимиляты приобретают основное значение для дальнейших продукционных процессов после появления цветков и, особенно, за- вязей и бобов. В данный период возникает недостаток пластических веществ по причине образования большого количества генетических органов (почек, цветков). Вследствие этого развивается явление абор-тивности цветков и бобов [7]. У культурных сортов из общего количества цветочных почек только 21–39 % образовали бобы (табл. 2).
Остальные завязи цветков теряются в связи с недостатком питательных веществ и неблагоприятными внешними условиями. По данному признаку также выявлены значительные сортовые различия. У сорта Смена количество почек, сформировавших бобы, составило 40 %, тогда как у сорта Янтарная – 25 %. Однако у сортов МК 1 и Янтарная зерновая продуктивность оказалась одной из самых высоких при максимальных показателях абортив-ности. Вне конкуренции по анализируемым признакам находится дикая соя. Указанная форма, видимо за счет сохранения 67 % репродуктивных органов, воспроизвела максимальную массу семян в расчете на одно растение.
Таблица 2
Количество цветочных почек, способных формировать бобы, у сортов сои амурской селекции
|
Сорт |
Общее количество цветочных почек, в расчете на 1 растение, шт. |
Количество почек, сформировавших бобы, в расчёте на 1 растение, шт. |
Процент почек, образовавших бобы, % от общего количества |
Масса семян в расчете на 1 растение, г |
|
МК 1 |
143 |
41 |
27 |
21,8 |
|
Амурская 310 |
129 |
41 |
31 |
19,7 |
|
Янтарная |
170 |
44 |
25 |
20,9 |
|
ВНИИС 2 |
158 |
33 |
32 |
16,1 |
|
Смена |
110 |
44 |
39 |
19,6 |
|
Glycine ussuriensis |
551 |
373 |
67 |
27,2 |
|
НСР 05 |
3,4 |
Можно заключить, что изучавшаяся дикорастущая форма сои по использованию пластических веществ при формировании семян обладает высокой степенью «экономичности». Среди культурных форм также имеются сортообразцы с достаточно оптимизированной системой реализации энергетических соединений. Объясняется это, по-видимому, различной направленностью и напряженностью использования ассимилятов у изучаемых сортов по периодам развития.
При последовательном прохождении генеративного и продуктивного развития закономерности перераспределения пластических веществ у растений могут быть недостаточно скоординированы. Заметно различаются эти процессы у детерми-нантных и недетерминантных сортовых групп сои, особенно при поступлении ас-симилятов из листьев в бобы каждого конкретного яруса. По данным Белико ва И.Ф. [1] , в первой половине вегетационного периода продукты фотосинтеза поступают в бобы близлежащего яруса. Впоследствии из - за неблагоприятных условий и абортивности поступление ас-симилятов в бобы разных ярусов приобретает конкурентный характер. Но даже на повышенном агрофоне абортивность цветков и бобов не снижается.
В таблице 3 приведены сведения о динамике изменения массы бобов в течение фазы бобообразования и по итоговой массе семян.
Таблица 3
Динамика накопления сухого вещества репродуктивными органами сои за период начало образования бобов – созревание, сорт сои Амурская 310 [7]
|
Вариант |
Продолжительность периода от первого учёта до созревания (по годам) |
|||
|
дата |
19 дней |
дата |
16 дней |
|
|
Сухая масса одного боба, мг |
||||
|
Контроль |
17 августа |
3,50 |
8 августа |
9,20 |
|
24 августа |
1,74 |
16 августа |
2,90 |
|
|
4 сентября |
0,61 |
4 сентября |
2,20 |
|
|
N 30 Р 60 К 30 |
17 августа |
3,20 |
8 августа |
10, 0 0 |
|
24 августа |
1,65 |
16 августа |
3,60 |
|
|
4 сентября |
0,64 |
4 сентября |
2,70 |
|
|
Сухая масса одного семени, мг (среднее из трёх повторений) |
||||
|
Контроль |
0,80 |
0,93 |
||
|
N 30 Р 60 К 30 |
0,80 |
1, 0 0 |
||
|
НСР 05 мг |
0,04 |
0,06 |
||
Наблюдается постепенное снижение количества поступивших ассимилятов в расчете на один боб в связи с увеличением репродуктивных единиц и частичной их утраты (опадение). При этом часть питательных веществ теряется безвозвратно. В этот же период приостанавливается образование новых клубеньков. Различия по накоплению массы бобов и массы семян удобренных и неудобренных растений выражены слабо. В целом у сои доля генеративных органов, реализовавших свой потенциал, невысока и определяется в основном фенотипическими факторами [1; 7]. Однако существенные сортовые различия по этому признаку свидетельствуют о возможности использования генетических механизмов для рационализа ции процессов перераспределения пластических веществ в пользу репродуктивных органов, способных завершить развитие. Установлено, что при несколько меньшей продолжительности периода образования бобов и более рациональном использовании симбиотической системы каждый отдельный боб и каждое отдельное семя получают увеличенное количество пластических веществ.
В определённой мере это подтверждается данными, приведёнными в таблице 4.
Изучаемые сорта сои амурской селекции заметно различаются длиной вегетационного периода. По региональной производственной классификации формы, созревающие более чем за 105–115 дней , считаются позднеспелыми, за 102– 112 дней – среднеспелыми, за 95–101 день – раннеспелыми. Выявлены в динамике определённые пределы изменчивости нарастания массы растения - хозяина, числа и массы клубеньков. Фенотипические проявления сортовых особенностей также в основном определяются продолжительно стью вегетирования. Самый позднеспелый (среди изучавшихся форм) сорт МК 1 сформировал максимальные вегетативную массу и урожай семян. Однако зерновая продуктивность не всегда соответствует масштабам нарастания вегетативной массы .
s

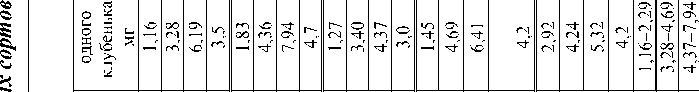
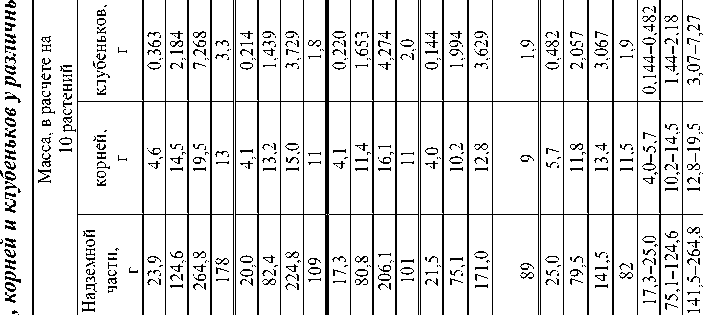
Так , у сорта Смена с периодом вегетации 96 дней урожай семян сформировался такой же, как у сорта Янтарная с периодом вегетации 107 дней. В вариантах с МК 1 и ВНИИС 1 этот показатель оказался практически одинаковым при разнице в веге-тировании в восемь дней.
В соответствии с масштабами накопления фитомассы сорт МК 1 превосходил другие варианты по итоговой массе и количеству клубеньков. Это свидетельствует о достаточно активном функционировании симбиотической системы при наличии значительных ресурсов ассими-лятов. Следует отметить у всех испытывавшихся сортов высокую степень соответствия системы «сорт – штамм». В лугово - чернозёмовидной почве доминирует сообщество штаммов ( Bradyrhi-zobium japonicum ), относящееся к серологической группе 617. Кроме того , встречаются формы , относящиеся к серогруппам 646 и 203, а также некоторое количество штаммов , серологически не идентифицированных [11]. В зависимости от условий активность симбиотической азотфиксации может нарушаться под воздействием другой аборигенной популяции ризобий.
При формировании фитомассы растения сои определённые ресурсы затрачивают на построение и функционирование симбиотического аппарата. Усреднённые за вегетационный период показатели массы клубеньков в процентах от массы надземной части составляют от 1,8 до 2 (табл. 5). Кроме того, значительное количество энергетических веществ затрачивается на транспортировку веществ при обменных процессах между клубеньками и другими органами растений. Количество белка и масла, затрачиваемое на такие реакции, выраженное в углеродных эквивалентах, обозначается как «рабочая энергия» [16].
Таблица 5
Показатель массы клубеньков (среднее за вегетационный период) в процентах от массы надземной части растений сои
|
Показатель |
Сорт |
||||
|
МК 1 |
Амурская 310 |
Янтарная |
ВНИИС 1 |
Смена |
|
|
Масса клубеньков |
1,8 + 0,2 |
1,65 + 0,15 |
2,0 + 0,2 |
2,2 + 0,21 |
2,3 + 0,35 |
Таким образом, динамика нарастания надземной части, корней, массы и числа клубеньков у сои представляет собой высокодинамичный процесс, требующий дополнительного изучения. Доминирующая закономерность заключается в том, что масштабы накопления органического вещества в значительной мере соответствуют показателям развития микросимбионта. В пределах этой закономерности распределение пластических веществ между органами растений определяется сортовой спецификой, которая в свою очередь складывается из достаточно сложных реакций массообмена между репродуктивными и вегетативными органами по этапам и фазам развития культуры и может изменяться под воздействием генетических и фенотипических проявлений.
Таким образом , расширение посевов сои в России (по вероятностным вариантам от 6 до 14 млн га) будет способствовать созданию новых высокоадаптиро ванных сортов на базе уже имеющегося сортового набора, обладающего большим разнообразием хозяйстственно ценных свойств. При этом потребуется преодоление биолого - ценотического порога урожайности.
-
2. Для преодоления определённого порога урожайности целесообразно использовать биологические особенности сои как бобовой культуры, включая изменчивость некоторых реакций. Заметной изменчивостью, при напряженном биосинтезе высокобелковых соединений , характеризуются фотопериодические реакции, наступление – прохождение этапов органогенеза (и их фенотипическая
-
3. С первых дней роста корни сои создают предпосылки для возникновения симбиоза, выделяя разнообразные корневые эксудаты (включающие изофлава-ноиды), которые взаимодействуют с бактериями, обладающими Nod- факто ром. С этого момента растения в течение всей вегетации затрачивают определённые ресурсы на формирование симбиотического аппарата. На 5– 7- й день после прорастания корня могут визуально наблюдаться клубеньки.
-
4. У изучаемых дальневосточных сортов сои (МК 1, Амурская 310, Янтарная, ВНИИС 1, Смена) выявлены существенные различия по масштабам нарастания листовой поверхности и продуктивности фотосинтеза. Наиболее эффективными по последнему признаку являются сорта МК 1, ВНИИС 2 и дикорастущая форма сои (соответственно 5,20 ; 4,87; 7,10 г/см 2 ).
-
5. Особенности развития у разных сортов сои связаны с недостаточно эффективными (непроизводительными) затра тами (на различных этапах органогенеза) пластических веществ, направленных на формирование репродуктивных органов. Во многом это обусловлено явлениями абортивности цветков и бобов и неполной вызреваемостью семян.
-
6. Количество почек, образовавших бобы , у изучаемых сортов колеблется от 25 до 67 % от общего числа этих органов. У сортов Смена, ВНИИС 2 и Амурская 310 данный показатель составляет соответственно 39 , 32 и 31 %. У дикорастущей формы сои продуктивных почек оказалось 67 %.
-
7. В зависимости от продолжительности периода бобообразования масштабы перераспределения ассимилятов могут колебаться (в расчете на единицу) у бобов от 0,64 до 3,5 мг и от 2,2 до 10,0 мг , у семян – соответственно 0,04–0,80 и 0, 6 –0,93 мг. Данные закономерности могут регулироваться генетическими и фенотипическими комбинациями.
-
8. Сопоставление динамики развития надземной части, корней и клубеньков у нескольких сортов сои свидетельствует о высоком уровне соответствия в системе «сорт – штамм». Затраты на формирование симбиотического аппарата составляют от 1,8 до 2,3 % от надземной массы растений в фазе налива бобов. Эффективность функционирования симбиотической системы также определяется «экономичностью» использования пластических веществ и свойствами микросимбионта.
зависимость) и процессы симбиотической азотфиксации.
Список литературы Биология сои: возможности оптимизации отдельных продукционных процессов
- Беликов И.Ф. Взаимоотношения между листовым аппаратом и органами плодоношения у сои: автореф.. д-ра биол. наук. -Владивосток, 1963. -36 с.
- Ващенко А.П., Мудрик Н.В., Фисенко П.П., Дега Л.А., Чайка Н.В., Капустин Ю.С. Соя на Дальнем Востоке. -Владивосток: Дальнаука, 2010. -435 с.
- Вэнс К. Симбиотическая фиксация у бобовых: сельскохозяйственные аспекты//Rhizobiaceae, молекулярная биология бактерий, взаимодействующих с растениями. -СПб., 2002. -С. 541-563.
- Джонсон Г.В., Бернард Р.Л. Генетика и селекция сои//Соя/Под ред. В.Б. Енкена. -М.: Колос, 1970. -С. 11-98.
- Жученко А.А. Экологическая генетика культурных растений. -Самара, 2003. -275 с.
- Куперман Ф.М., Ржанова Е.И. Биология развития растений. -М.: Высш. шк.,1963. -424 с.
- Лопаткина Э.Ф. Распределение пластических веществ по органам растений сои в зависимости от разных уровней почвенного питания//Науч.-тех. бюл. ВНИИ сои: Частные вопросы генетики, биологии и физиологии сои. -1977. -Вып. 7-8. -С. 10-19.
- Лопаткина Э.Ф. Методика количественного учета репродуктивных органов сои//Науч.-тех. бюл. ВНИИ сои: Частные вопросы генетики, биологии и физиологии сои. -1977. -Вып. 7-8. -С. 34-42.
- Лопаткина Э.Ф. Использование морфофизиологического метода при оценке исходного материала сои: методические рекомендации. -Новосибирск, 1991. -27 с.
- Лукомец В.М., Зеленцов С.В., Кривошлыков К.М. Перспективы и резервы расширения производства масличных культур в Российской Федерации//Масличные культуры. Науч.-тех. бюл. ВНИИМК. -2015. -Вып. 4 (164). -С. 81-102.
- Новикова А.Т., Тильба В.А., Бегун С.А. Определение серологической общности клубеньков на корнях сои в почвах Приамурья//Науч.-тех. бюл. ВНИИ сои: Соя и нитрагин. -1976. -Вып. 1. -С. 35-38.
- Петибская В.С. Соя: химический состав и использование. -Майкоп, 2012. -432 с.
- Тильба В.А., Бегун С.А., Дуденко Н.Г. Сравнительная динамика образования клубеньков у некоторых амурских сортов сои//Биология, селекция и генетика сои. -Новосибирск, 1986.-С. 83-92.
- Титова Л.В., Леонова Н.О., Антипчук А.Ф. Азотфиксирующие микроорганизмы в микробно-растительных системах//Биорегуляция микробно-растительных систем/Под ред. А.И. Иутинской, С.П. Пономаренко. -Киев: Ничлава, 2010. -С. 99-194.
- Федотов В.А., Гончаров С.В., Столяров О.В., Ващенко Т.Г., Шевченко Н.С. Соя в России/Под ред. В.А. Федотова, С.В. Гончарова -М.: Агролига России, 2013. -430 с.
- Хауэл Р.В. Физиология сои//Соя/Под ред. В.Б. Енкена. -М.: Колос, 1970. -С. 99-164.
