Colorectal cancer: angiogenesis and matrix metalloproteinases

Author: Maiborodin I.V., Fursov S.A., Solovenchuk L.L., Gerus P.A., Sheplev B.V., Chernyshova A.L.
Journal: Сибирский онкологический журнал @siboncoj
Section: Обзоры
Article in issue: 2 т.25, 2026.
Free access
Aim: to summarize the available literature data on the role of matrix metalloproteinases (MMPs) in angiogenesis, as well as in the development and progression of colorectal cancer (CRC). Material and Methods. A literature search was conducted in the Medline and eLI BRARY.RU databases from 1999 to the present using the keywords “angiogenesis + cancer + colon + MMP”. Results. Malignant tumors are characterized by the ability to form new vessels, invade surrounding tissues, and metastasize to distant organs, and this aggressiveness depends on the expression of proteolytic enzymes, including MMPs. In CRC, the levels of MMPs are increased in both tumor tissues and blood plasma; these enzymes are required for extracellular matrix remodelling during angiogenesis, tumor invasion and metastasis. The degree of overexpression of certain MMPs correlates with the stage of CRC, vascularization, VEGF expression and/or prognosis; however, not all researchers agree with these findings. MMPs are synthesized both by tumor cells and, more prominently, stromal cells like tumor-associated macrophages. Alterations in the function of immunocompetent and stromal cells that express and produce MMPs play a key role in both the initiation and progression of CRC. Targeting tumor-associated immunocompetent and stromal cells, their reprogramming, neutralization and/or inhibition of their trafficking may delay the growth and invasion of CRC. Inhibition of MMPs may represent one of the stages of antiangiogenic therapy. In addition to suppressing angiogenesis, certain MMP inhibitors stimulate apoptosis, suppress cell proliferation, induce cell cycle arrest, and reduce the expression of antiapoptotic genes of the Bcl family. A wide range of artificial and natural compounds and methods has been proposed as MMP inhibitors. However, despite promising experimental results, clinical trials of MMP inhibitors have been largely disappointing. There is evidence that some MMPs exert antitumor effects, and their blockade may be accompanied by tumor progression. Conclusion. A wide variety of approaches to targeting MMPs, together with the certain inconsistency of available data, may indicate the persisting dissatisfaction of researchers and clinicians with the current solutions to the problem of MMP-mediated angiogenesis in tumor tissues. This also highlights the need for continued investigation into both the effects of MMPs in CRC and the methods for controlling the activity of this enzyme group.
Colorectal cancer, matrix metalloproteinases, tumor angiogenesis, tumor vascularization, tumor-associated macrophages, inhibitors of matrix metalloproteinases, prognostic factors
Short address: https://sciup.org/140314633
IDR: 140314633 | UDC: 616.345-006.6:616.13:577.15 | DOI: 10.21294/1814-4861-2026-25-2-165-174
Рак толстой кишки: ангиогенез и матриксные протеиназы
Цель исследования – обобщить данные о роли матриксных металлопротеиназ (ММП) в ангиогенезе, развитии и прогрессировании рака толстой кишки (РТК). Материал и методы. Проведен поиск публикаций в базах данных Medline и eLI BRARY.RU с 1999 г. по настоящее время по ключевым словам «angiogenesis + cancer + colon + MMP». Результаты. Наиболее распространенным свойством злокачественных опухолей является их способность формировать новые сосуды, проникать в соседние ткани и метастазировать, и эта агрессивность зависит от экспрессии протеолитических ферментов, в том числе ММП. При возникновении РТК в тканях и плазме крови возрастает содержание ММП, которые необходимы для ремоделирования внеклеточного матрикса в процессе ангиогенеза, инвазии и метастазирования опухоли. Степень повышенной экспрессии некоторых ММП коррелирует со стадией РТК, васкуляризацией, экспрессией VEGF и/или прогнозом, но с этим согласны не все исследователи. ММП синтезируются как клеточными элементами опухоли, так и клетками, расположенными в строме, в первую очередь туморассоциированными макрофагами. Изменение функций иммунокомпетентных и стромальных клеток, экспрессирующих и продуцирующих ММП, играет решающую роль как в инициации, так и прогрессировании РТК, в том числе через вновь образованные сосуды. Воздействие на туморассоциированные иммунокомпетентные и стромальные клетки, их перепрограммирование, нейтрализация и/или ингибиция трафика способны задержать васкуляризацию, рост и инвазию РТК. Ингибирование ММП может стать одним из этапов антиангиогенной терапии. Кроме супрессии ангиогенеза, некоторые ингибиторы ММП стимулируют апоптоз, подавляют пролиферацию клеток, индуцируют остановку клеточного цикла и снижают экспрессию антиапоптотических генов семейства Bcl. В качестве ингибиторов ММП предложено использование большого количества различных методик, искусственных и естественных соединений. Однако несмотря на многообещающие экспериментальные результаты, клинические испытания с ингибиторами ММП не оправдали ожиданий. Имеются данные об антиопухолевом эффекте некоторых ММП, блокада которых сопровождается прогрессированием рака. Заключение. Такое значительное число способов воздействия на ММП и определенная противоречивость данных могут служить свидетельством неудовлетворенности исследователей и клиницистов решением проблемы ангиогенеза с участием ММП в тканях опухолей, целесообразности продолжения изучения как эффектов ММП при РТК опухоли, так и методов контроля активности этой группы ферментов, средств управления ими.
Text of the scientific article Colorectal cancer: angiogenesis and matrix metalloproteinases
Angiogenesis is a physiological process involving the growth of new blood vessels from pre-existing ones; however, it also plays a crucial role in tumor progression [1]. Cancer cells require nutrients to support the growth of the primary tumor mass and metastatic dissemination. This is ensured by tumor-associated neovascularization, angiogenesis, which also facilitates the transition from hyperplasia to neoplasia, that is, to a state of uncontrolled cellular proliferation [2, 3]. In chronic inflammatory bowel diseases, the mi-crovascular network expands through intussusceptive angiogenesis, which includes increased blood flow and the splitting of existing capillaries. Such pathological angiogenesis contributes to the initiation and progression of cancer [4].
The most characteristic property of malignant tumors is their ability to invade neighbouring tissues and metastasize, and this aggressiveness depends on the overexpression of proteolytic enzymes. Matrix metalloproteinases (MMPs) constitute a family of zinc-dependent endopeptidases that degrade the extracellular matrix and participate in a wide range of pathophysiological processes, including cardiovascular diseases, arthritis, periodontitis, osteogenesis imperfecta and disorders of the central nervous system. MMPs play a leading role in the initiation and development of many types of cancer, including colorectal cancer (CRC), as well as in tumor invasion and metastasis [5–7]. Even MMPs that lack enzymatic activity towards matrix components (MMP-11, stromelysin-3) are associated with tumor progression in humans and with unfavorable clinical outcomes [8]. The key MMPs involved in pathological processes in CRC include MMP-2, -3, -7, -9, -10, -12 and -13. All MMPs are closely linked to immune function and influence the immune system by modulating the differentiation and activity of immune cells, as well as by promoting the recruitment of macrophages and neutrophils in colitis-associated CRC [7, 9–11].
Increased MMP expression correlates with reduced survival [6, 12]. Members of the MMP family are overexpressed in malignant tumors across many cancer types, including breast cancer, CRC, prostate cancer, oral cancer, and others [13]. M. Buttacavoli et al. [6] demonstrated the role of MMPs in epithelial – mesenchymal transition and immune responses. The activity of multifunctional MMPs is regulated by tissue inhibitors of metalloproteinases (TIMPs), which are critically important in modulating the pro-inflammatory NF-κB pathway (a family of transcription factor proteins that play a key role in the regulation of inflammation, immune responses, apoptosis and proliferation; this signalling pathway is activated in response to various stimuli, resulting in translocation of the NF-κB complex to the cell nucleus and the initiation of gene expression) in the cellular components of various tumors [10].
The aim of this study was to summarise the available literature data on the role of MMPs in angiogenesis, as well as in the development and progression of colorectal cancer.
Material and Methods
A literature search was conducted in the Medline and eLIBRARY.RU databases from 1999 to the present using the keywords “angiogenesis + cancer + colon + MMP”. A total of 159 sources were identified during this period. Full-text articles in Russian and English were included in the analysis, encompassing the following types of publications: basic research studies, experimental studies, clinical studies and case reports. Reviews and articles addressing changes in MMP levels and expression during CRC treatment not specifically targeting angiogenesis were excluded. Of the sources identified, 47 were included in the present review.
Matrix Metalloproteinasesas a Factor in Colorectal Cancer Progression
The expression of MMP-2 and MMP-9 is significantly increased in colorectal cancer (CRC) tissues, predominantly in the tumor stroma, compared with the corresponding non-tumorous colonic tissue. Increased MMP-2 content has been detected by immunohistochemistry in areas where tumor cells invade the muscular layer. Plasma levels of MMP-2 and MMP-9 were also higher in patients with CRC and showed a strong correlation with clinical stage. The concentrations of both enzymes returned to normal values after radical surgery at all stages of the disease [14]. At the same time, H. Shantha Kumara et al. [7] reported that MMP-2 levels remained elevated for 5 weeks after surgery, while MMP-7 levels persisted for 2-6 weeks. This may be attributable to surgical trauma and wound healing; however, it cannot be excluded that such dynamics may also reflect residual tumor growth and metastasis.
-
F. Klupp et al. [5] identified a significant overexpression of MMP-7, -10 and -12 in the serum of 78 patients with CRC compared with a healthy control group. In the presence of vascular invasion, MMP-12 expression was significantly higher than in CRC in situ . Overexpression of MMP-7, -10 and -12 was associated with a marked reduction in overall survival. Multivariate analysis demonstrated that a high serum MMP-10 level is an independent adverse prognostic marker in patients with CRC. Using bioinformatic-based computer modelling, N. Mokgautsi et al. [15] showed that a high level of expression of vascular endothelial growth factor A (VEGF)/β-catenin/MMP-7/CD44 in CRC indicates tumor progression, the presence of cancer stem cells, therapeutic resistance, metastasis, and unfavorable clinical outcomes. Increased MMP-14 expression is also associated with a poor prognosis in cancer patients [16], whereas reduced expression of VEGF and MMP-9 genes is directly associated with the suppression of angiogenesis and metastasis [17]. These findings were confirmed in animal experiments by H. Gungor et al. [18]: during the induction of CRC in rats using 1,2-dimethylhydrazine dihydrochloride, blood samples showed a significant increase in MMP-2 and MMP-9 levels and in the MMP-2/TIMP-2 ratio compared with the control and treatment groups.
-
Y. Takahashi et al. [19] investigated vessel density and the expression of VEGF and MMP-7 in cellular elements from samples of 25 adenomas, 35 in situ mucosal tumors, 29 CRCs invading the submucosa, and 33 CRCs infiltrating the muscularis, using immunohistochemical methods. Pronounced statistically significant differences in vessel density and in the expression of VEGF and MMP-7 were identified as
the tumor progressed from the mucosa to infiltration of deeper layers of the intestinal wall. The angiogenic switch may occur simultaneously with the onset of submucosal invasion at the early stages of CRC development.
The occurrence of an early angiogenic switch in CRC progression is also confirmed by C.A. Staton et al. [20]. A total of 210 surgical specimens of the colon, including adenomas and/or carcinomas, were immunohistochemically stained for endothelial cells (CD31) and VEGF. A significant increase in vessel density was observed in CRC, with a significant correlation with Dukes’ stage and lymph node involvement. The greatest increase in vascularisation was associated with the onset of dysplasia, accompanied by a significant increase in VEGF expression (p<0.0005). No correlation was found between VEGF and vessel density, tumor size, Dukes’ classification, lymph node involvement, or survival.
During the development of induced CRC in Balb/c mice, S. Setia et al. [21] observed an upregulation of the angiogenic pathway accompanied by increased expression of VEGF, MMP-2 and MMP-9. An elevated plasma MMP-9 level in mice lacking integrin-α1 further enhanced the metastasis of CRC cells to the lungs [22]. In mice deficient in the protease MMP-14 in endothelial cells, restricted intussusceptive angiogenesis was noted in the capillary plexus of the colonic mucosa during induced colitis. Endothelial MMP-14 activity is essential for vasodilation and intussuscep-tive angiogenesis [4].
V.K. Bozhenko et al. [11] examined the expression levels of MMP-7, MMP-8, MMP-9 and MMP-11, as well as the proliferation marker Ki67, in 256 tissue samples obtained from 112 patients with CRC: 112 samples of primary tumor tissue, 112 samples of morphologically normal colonic mucosa excised 15–20 cm away from the primary tumor, and 16 samples of CRC liver metastases. In addition, 16 biopsy specimens obtained during colonoscopy from healthy volunteers were analysed. It was found that the mRNA expression of genes encoding MMPs and Ki67 was higher in primary CRC tissue and in metastases compared with normal tissue, whether taken from areas distant from the tumor or from healthy donors. The proliferative activity of primary CRC cells and metastatic cells was similar. The study also showed that normal colonic mucosa from CRC patients had significantly higher levels of MMP-8 mRNA expression compared to healthy donors.
It should be noted that, according to S. Zucker and J. Vacirca [14], increased expression of MMP-12 – unlike other MMPs – is associated with improved survival in CRC, presumably owing to its inhibitory effect on angiogenesis. O.O. Uslukaya et al. [23] analysed 34 fresh CRC samples and paired adjacent non-tumorous tissues. Tumors demonstrated a statistically significant increase in the expression of cyclooxygenase-2 (COX-2), interleukin-8 (IL-8) and VEGF genes.
However, MMP-2 and MMP-9 expression levels did not differ between tumor and normal tissues.
The renin – angiotensin system is activated in patients with CRC and is associated with poor prognosis and chemoresistance. Losartan markedly reduced the growth of CRC xenografts in mice and enhanced tumor cell necrosis. Furthermore, these animals showed inhibition of MMP-2 and MMP-9 activity, and as a consequence, tumor vascularisation was reduced and metastasis was suppressed [24].
Thus, the onset of CRC is accompanied by an increase in MMP levels in both tissues and plasma, and these enzymes are required for extracellular matrix remodelling during tumor angiogenesis, invasion and metastasis. Pronounced statistically significant differences in vessel density, VEGF expression and MMP levels emerge when CRC progresses from the mucosa to infiltration of deeper layers of the intestinal wall. After tumor resection or radical surgery, MMP levels decrease sharply; this may reflect suppression of angiogenesis and invasion, although delayed normalisation may result from surgical trauma as well as residual tumor growth and metastasis of the remaining tumor. The degree of overexpression of certain MMPs correlates with the stage of CRC, vascularisation, VEGF expression and/or prognosis; however, not all researchers agree with these findings.
The Relationship between MMPs and Tumor-Associated Leukocytes and Fibroblasts
Chronic intestinal inflammation is a significant risk factor for colorectal cancer. The development of colitis and CRC is a complex multifactorial process involving chronic inflammation, immune regulation and remodelling of the tumor microenvironment. Macrophages are among the most common tumor-associated cells and play a key role in maintaining intestinal homeostasis and in the transition from colitis to CRC. Macrophages are activated mainly in two ways: classically activated macrophages (pro-inflammatory, M1) and alternatively activated macrophages (anti-inflammatory, M2), with these phagocyte subsets performing distinct roles during inflammation [25].
During the progression of chronic colitis to cancer, the proportion of M2 macrophages gradually increases. M2 macrophages promote inflammation resolution and contribute to tissue repair via angiogenesis at the later stages of inflammation [26]. They are divided into M2a, M2b, M2c and M2d subpopulations. M2a and M2b macrophages participate in immune regulation by stimulating Th2-cell responses, whereas M2c cells are associated with immunosuppression and tissue remodelling [27]. The M2d subtype is activated predominantly by tumor cells, tumor-associated factors or chemotherapeutic agents. This macrophage subset has immunosuppressive and tumor-promoting functions [28] and specifically expresses IL-10, transforming growth factor-β (TGF-β) and VEGF, which stimu- late angiogenesis and matrix remodelling and create favorable conditions for the proliferation, infiltration and migration of cancer cells, contributing to tumor progression and angiogenesis [25, 28, 29]. High levels of M2 macrophage infiltration are associated with poor prognosis in CRC patients [30].
MMPs are synthesized not only by tumor cells themselves, but also by cells located in the stroma, primarily tumor-associated macrophages (TAMs) [31]. Interestingly, M1 and M2 phagocytes exhibit different proteolytic activities, with IL-10 – stimulated TAMs inducing MMP-2 and MMP-9 activity more effectively. Using a broad-spectrum MMP inhibitor, it was demonstrated that proteolysis is required for TAM-mediated cancer cell invasion and angiogenesis [32].
TAMs infiltrating CRC express hypoxia-inducible factor 1α (HIF-1α) as an “angiogenic switch” and themselves respond to hypoxic signals by producing pro-angiogenic cytokines and growth factors such as angiopoietin-2, VEGF, interleukin-8 (IL-8) and fibroblast growth factor-2 (FGF-2). These promote the recruitment, proliferation and maturation of endothelial cells, thereby forming new blood vessels and facilitating tumor growth [33].
The number of TAMs in CRC correlates with both blood and lymphatic vascularization [34]. Perivascular macrophages secrete chemotactic signals guiding the migration of CRC cells along collagen fibres towards blood vessels, enabling their entry into the circulation. TAMs may also activate the epidermal growth factor (EGF)/VEGF receptor-3 (VEGFR-3) axis, which supports the formation of lymphatic vessels and provides a route for tumor cells to penetrate the lymphatic system [35].
Levels of miR-92a-3p are increased in plasma exosomes of CRC patients and positively correlate with lymph-node metastasis. miR-92a-3p promoted the polarization of M0 macrophages towards an M2 phenotype by activating the MAPK/ERK ((mitogen-activated protein kinase/extracellular signal-regulated kinase) signalling pathway (one of the key pathways triggered by extracellular signals such as hormones, growth factors, chemokines and neurotransmitters recognized by receptor tyrosine kinases or G-protein-coupled receptors; ERK signalling ultimately promotes cell survival, proliferation and motility). It also enhanced the migratory and invasive capabilities of tumor cells and increased their angiogenic potential in vitro [29].
Macrophage production is regulated by colonystimulating factor-1 (CSF-1), the expression of which is significantly elevated in CRC xenografts in mice. Inhibition of CSF-1 can suppress tumor growth by slowing macrophage-mediated degradation of the extracellular matrix: the growth of xenografted embryonic tumors was reduced to a quiescent state, and CRC growth decreased by 50 %. Furthermore, tumor vascularization, MMP-2 expression and levels of pro-angiogenic factors were reduced. Six-month survival was observed only in mice with CRC after
CSF-1 blockade, whereas all control animals died by day 65 [31].
Accumulation of neutrophils in tissues is a recognized feature of both ulcerative colitis and CRC. It has been shown that MMP-14 is strongly induced in tumor-associated neutrophils (TANs), where it regulates chemotaxis and endothelial cell proliferation. Clinical samples confirmed increased MMP-14-positive TANs in high-grade CRC, but not in patients with ulcerative colitis. Pharmacological inhibition of TAN trafficking or MMP-14 activity effectively reduced tumor vessel density, leading to CRC regression [36].
According to Y. Xiao et al. [12] and A. Mezhey-euski et al. [30], infiltration of colorectal cancer (CRC) by CD8+ T cells is associated with improved patient survival. However, as reported by M. Gallazzi et al. [37], natural killer cells – effector lymphocytes of the innate immune system – undergo alterations in several types of cancer. In CRC, circulating and tumor-associated/infiltrating natural killer cells displayed a pro-angiogenic phenotype and produced factors capable of sustaining inflammatory angiogenesis in an in vitro model. Secretome analysis revealed that such natural killer cells secrete pro-inflammatory cytokines and chemokines involved in monocyte recruitment and their M2-like polarization.
Cancer-associated fibroblasts, or activated fibroblasts of the tumor stroma, are important modifiers of tumor progression. Using several in vivo experimental models, it was demonstrated that expression of MMP-11 by fibroblasts surrounding malignant epithelial cells stimulates tumor formation in a paracrine manner. This effect was not due to increased neoangiogenesis or enhanced proliferation of cancer cells, but rather to reduced cell death through inhibition of apoptosis and necrosis [8].
Antiangiogenic therapy is widely used in CRC treatment; however, many patients eventually develop resistance and experience disease progression. Vas-culogenic mimicry, i.e., the formation of vessel-like networks by tumor cells is one mechanism contributing to this resistance. In tissue samples from CRC patients (n=157), follistatin-like protein 3 (FSTL3) was identified as a critical factor predominantly expressed in CRC-associated fibroblasts. Its expression strongly correlated with increased vasculogenic mimicry, the presence of intratumoral blood vessels and poor prognosis. Single-cell RNA sequencing of tumors from patients with and without vasculogenic mimicry showed that hypoxia stimulates FSTL3 expression in tumor fibroblasts, leading to extracellular matrix remodelling and enhanced endothelial-like plasticity of tumor cells. Anti-FSTL3 antibodies effectively inhibited angiogenesis in both in vitro and in vivo models, and their combination with bevacizumab produced synergistic suppression of neo-vascular-like structures and distant metastases [38].
Wnt5a, a ligand of the Wnt family (a group of intracellular signalling pathways regulating embryogenesis, cell differentiation and tumor development; disruptions in these pathways may lead to cancers such as CRC), activates the β-catenin-independent signalling cascade and promotes tumor formation. A. Harada et al. [39] showed that Wnt5a is expressed in fibroblasts adjacent to CRC in mice, and that deficiency of this ligand suppresses tumor development. Wnt5a expression is induced in fibroblasts under inflammatory conditions and hypoxia, accompanied by activation of HIF-2. Moreover, fibroblasts under hypoxia produce epiregulin, which also stimulates CRC growth.
Alterations in the functions of immunocompetent and stromal cells play a decisive role in both the initiation and progression of CRC. MMPs are synthesized by tumor cells and stromal cells, primarily TAMs, and proteolysis is required for cancer cell invasion and angiogenesis. Macrophages differentiate into the M2 phenotype during chronic colitis to resolve inflammation and support subsequent tissue regeneration and angiogenesis aimed at restoring blood flow in damaged tissues. However, certain M2 phagocyte subtypes create a favorable microenvironment for CRC development through immune suppression and tissue remodelling, and subsequently contribute to cancer progression and tumor angiogenesis. During the progression of chronic colitis to cancer, the proportion of M2 macrophages gradually increases. The number of TAMs in CRC correlates with blood and lymphatic vascularization. High infiltration by M2 macrophages is associated with poor prognosis in CRC patients. In addition to macrophages, both ulcerative colitis and CRC are characterized by neutrophil infiltration, with TANs containing higher levels of MMPs than neutrophils present in chronic colonic inflammation. Regarding CD8+ T-cell infiltration in CRC, interpretations among researchers differ. Some consider it a favorable indicator associated with improved survival, while other data show that natural killer cells undergo cancer-related changes, acquire a pro-angiogenic phenotype and release factors that support inflammatory angiogenesis and promote monocyte recruitment and their M2-like polarization. CRC-associated fibroblasts stimulate angiogenesis and vasculogenic mimicry with endothelial-like tumor cell transformation via modulation of MMP expression. Targeting tumor-associated immune and stromal cells – through their reprogramming, neutralization and/or inhibition of trafficking – may delay CRC progression and invasion.
Effects of MMP Inhibition in Cancer
Angiogenesis is essential for tumor growth, and its inhibition may serve as an effective approach to controlling cancer cell expansion. MMP inhibition represents an attractive therapeutic target for intervening in multiple aspects of tumor progression. Local MMP production followed by extracellular matrix degradation contributes to sustaining tumor growth, invasion and angiogenesis; therefore, inhibition of proteolysis in cancer tissues, including MMP-mediated proteolysis, should attenuate metastasis, angiogenesis and migration [13].
MMP-9 inhibitors induce apoptosis, suppress cell proliferation, cause cell-cycle arrest and reduce expression of antiapoptotic genes Bcl-2 and Bcl-xl in vitro [40]. Bioinformatic analysis revealed a positive correlation between MMP-2 expression and PD-L1 levels (programmed cell death ligand 1 – a protein that suppresses the immune response by binding to the PD-1 receptor on T cells, thereby reducing their proliferation and cytokine secretion and inducing apoptosis) in CRC. In vitro , inhibition of MMP-2 markedly reduced PD-L1 expression in SW480 cells (a human CRC cell line), indicating a regulatory role of MMP-2 in tumor immune evasion. Moreover, a negative association was identified between MMP-2 expression and the presence of tumor-infiltrating lymphocytes, underscoring the potential involvement of MMP-2 in shaping the immunological landscape of CRC [12].
MMP-2 may serve as a target for cancer immunotherapy. Immunization against self-MMP-2 appears to be a promising strategy for treating MMP-2-positive tumors. Specific autoantibodies to MMP-2 were detected in the serum and on tumor cells of mice immunized with a xenogeneic MMP-2 vaccine. Such treatment prolonged survival in tumor-bearing mice and enhanced CD8+ T-cell activity. Its antitumor efficacy was abolished in vivo by depletion of CD4+ and CD8+ T-cell populations and was augmented by adoptive transfer of T cells from mice immunized against MMP-2 [41, 42].
Y.Y. Kimura et al. [43] examined the antiangiogenic effects of 2,3-, 3,4- and 4,4’-dihydroxystilbenes, which inhibited pro-MMP-9 production in colonic cells and suppressed VEGF-induced migration of human umbilical vein endothelial cells (HUVECs). These stilbenes inhibited CRC neovascularization in mice. The antiangiogenic activity of stilbenes may result from suppression of VEGF-induced endothelial cell migration and angiogenesis through inhibition of VEGFR2 phosphorylation in endothelial cells and downregulation of pro-MMP-9 expression in colonic cells.
TIMP-2 is a natural tissue inhibitor of MMPs that prevents extracellular matrix protein degradation. TIMPs suppress the hydrolytic activity of MMP-2 and MMP-9, which are selective for collagenolysis. TIMP-2 overexpression in vitro inhibits invasion of both tumor and endothelial cells without affecting their proliferation. Pre-infection of tumor cells with an adenovirus carrying the TIMP-2 gene suppressed the growth of human CRC C51 in more than 50 % of mice. A single local injection of this virus into established tumors reduced tumor growth rates by 60–80 % and the tumor angiogenesis index by 25–75 %. Treated mice also exhibited significantly increased survival [44].
Use of non-steroidal anti-inflammatory drugs (NSAIDs) reduces the risk of CRC development. Their anticancer effect involves suppression of β-catenin signalling pathways, reduction of angiogenic markers such as VEGF and MMP-9, and inhibition of cell proliferation. This effect is accompanied by pro-apoptotic activity, evidenced by increased DNA fragmentation and altered Bax/Bcl-2 ratios [2, 45].
P. Ghanghas et al. [2] used 1,2-dimethylhydrazine dihydrochloride as a carcinogenic agent to induce CRC in rats and administered three NSAIDs – celecoxib, etoricoxib or diclofenac – orally as chemopreventive agents. VEGF, MMP-2 and MMP-9 expression was significantly increased in the carcinogenesis group and markedly reduced by each NSAID. NSAID treatment also induced apoptosis. Inhibition of angiogenesis and induction of apoptosis by NSAIDs are likely mechanisms underlying CRC chemoprevention. Additionally, increased expression of VEGF and MMPs during the development and progression of induced CRC in experimental animals can be prevented by celecoxib [21].
It should also be noted that the soluble heat shock protein B1 is released predominantly from endothelial cells and regulates angiogenesis through direct interaction with VEGF. MMP-9 can cleave this heat shock protein and release antiangiogenic fragments that play a key role in tumor progression by inhibiting VEGF-induced endothelial cell activation. This suggests that MMP inhibitors may exert pro-tumorigenic effects by reducing HSPB1 fragmentation [46]. A. Pozzi et al. [47] provided evidence that reduced plasma MMP-9 levels in both wild-type and integrin-α1-deficient mice diminish angiostatin production and consequently enhance tumor growth and vascularization. Conversely, selective upregulation of MMP-9 expression in vivo reduced tumor vascularization. The findings of S.H. Choi et al. [46] and A. Pozzi et al. [47] contradict other studies suggesting a pro-tumorigenic role of MMP-9, but may explain some failures of anti-MMP therapy in cancer treatment.
Inhibition of MMPs with synthetic compounds was once considered a promising strategy for slowing or even halting cancer progression. However, despite promising experimental results, clinical trials of MMP inhibitors have been largely disappointing. However, given the multifunctionality of MMPs and the multifactorial nature of cancer, it is impossible to predict at which stage of tumor development and in which processes they are involved. Therefore, the use of MMP inhibitors in cancer therapy should be approached with caution [9, 14].
MMPs, as proteases capable of degrading the extracellular matrix, participate both in tumor invasion and in neovascularization. Inhibition of MMPs may represent one of the stages of antiangiogenic therapy. In addition to suppressing angiogenesis, certain MMP inhibitors stimulate apoptosis, suppress cell proliferation, induce cell cycle arrest, and reduce the expression of antiapoptotic genes of the Bcl family. A wide range of synthetic and natural compounds has been proposed as MMP inhibitors, including TIMPs, stilbenes and non-steroidal anti-inflammatory drugs (NSAIDs). Particular attention has also been given to immunotherapy and the blockade of MMP genes. However, despite encouraging experimental findings, clinical trials involving MMP inhibitors have demonstrated limited efficacy. It is also important to note that some MMPs exhibit anti-tumor properties, and their blockade is associated with tumor progression. The large number of approaches to modulating MMP activity described in the literature, combined with the inconsistency of available data, may indicate the relative insufficiency of current findings and an ongoing search for new strategies to inhibit these proteases. The clinical use of MMP inhibitors for the treatment of CRC is currently limited to experimental studies conducted in various institutions.
Conclusion
Thus, the most widespread characteristic of malignant tumors is their ability to invade adjacent tissues and metastasize, and this aggressiveness depends on the expression of proteolytic enzymes, including MMPs. In CRC, the levels of MMPs increase in both tissues
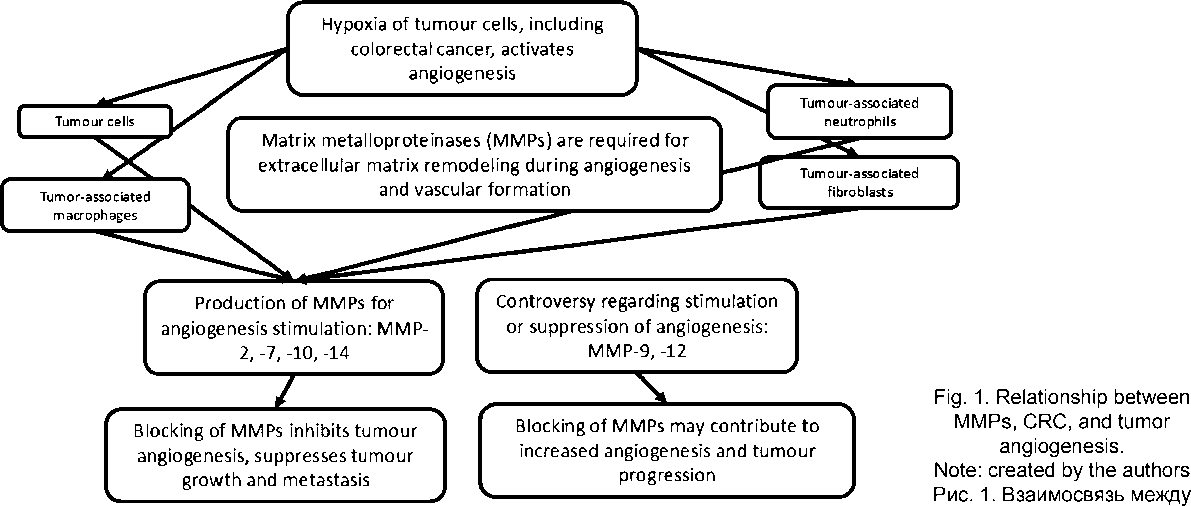
It is advisable to continue studying both the effects of MMPs in tumours and methods for monitoring the activity of this enzyme group матриксными металлопротеиназами, раком толстой кишки и опухолевым ангиогенезом.
Примечание: рисунок выполнен авторами
and blood plasma, and these enzymes are required for extracellular matrix remodelling during tumor invasion and metastasis. The degree of overexpression of certain MMPs correlates with the stage of CRC, vascularization, VEGF expression and/or prognosis; however, not all researchers agree with these findings. MMPs are synthesized both by tumor cells and by stromal cells, primarily tumor-associated macrophages (TAMs). Alterations in the function of immunocompetent and stromal cells that express and produce MMPs play a key role in both the initiation and progression of CRC. Targeting tumor-associated immunocompetent and stromal cells, their reprogramming, neutralization and/or inhibition of their trafficking may delay the growth and invasion of CRC. Inhibition of MMPs may represent one of the stages of antiangiogenic therapy. In addition to suppressing angiogenesis, certain MMP inhibitors stimulate apoptosis, suppress cell prolifera- tion, induce cell cycle arrest, and reduce the expression of antiapoptotic genes of the Bcl family. A wide range of artificial and natural compounds and methods has been proposed as MMP inhibitors. However, despite promising experimental results, clinical trials of MMP inhibitors have been largely disappointing. There is evidence that some MMPs exert antitumor effects, and their blockade may be accompanied by tumor progression. Such a wide variety of approaches to targeting MMPs, together with the certain inconsistency of available data, may indicate the persisting dissatisfaction of researchers and clinicians with the current solutions to the problem of MMP-mediated angiogenesis in tumor tissues. This also highlights the need for continued investigation into both the effects of MMPs in CRC and the methods for controlling the activity of this enzyme group (Fig. 1).
