Фенотипическое разнообразие чужеродных видов при разных механизмах вселения в бассейн р. Вычегда

Автор: Рафиков Р.Р.
Журнал: Принципы экологии @ecopri
Рубрика: Оригинальные исследования
Статья в выпуске: 3 (53), 2024 года.
Бесплатный доступ
В работе представлено описание морфологического разнообразия чужеродных для бассейна р. Вычегда популяций белоглазки и ротана-головешки. Их вселение произошло в результате разных механизмов инвазии - самостоятельного расселения и акклиматизации. Показано, что белоглазка широко распространилась по магистральному руслу р. Вычегда, вплоть до верхнего течения, а акклиматизированный ротан обитает в нескольких пожарных водоемах дачных обществ в районе среднего течения реки. Сравнение морфологических описаний выборки белоглазки с ее номинативной формой позволило установить более широкий размах изменчивости количества лучей в анальном плавнике и тычинок на первой жаберной дуге. Для ротана также отмечено большее число лучей в грудном и спинном плавниках. Данные изменения можно расценивать как результат адаптации исследованных популяций чужеродных видов к обитанию в условиях приобретенной части ареала. С помощью показателя µ Животовского выявлен сходный уровень внутрипопуляционного морфологического разнообразия выборок ротана и белоглазки. Это может указывать на общность протекающих процессов при вселении вида на новую территорию. Однако частота асимметричного проявления на признак в выборке ротана оказалась высокой (4 балла) и соответствовала уровню значительного отклонения от нормы. На основании проделанной работы выдвинута гипотеза о том, что популяция, формирующаяся из небольшого числа особей в водоеме малой площади, испытывает большее воздействие экологических факторов на период раннего онтогенеза по сравнению с самостоятельно расселяющимися по руслам рек видами. Предполагается, что в основе данного процесса лежат эффекты «основателя» и «горлышка бутылки».
Белоглазка, ротан-головешка, вселенцы, фенопитическое разнообразие, европейский северо-восток России, вычегда
Короткий адрес: https://sciup.org/147251105
IDR: 147251105 | УДК: 597.2/.5:574.91(282.247.133) | DOI: 10.15393/j1.art.2024.15142
Phenotypic diversity of alien species under different mechanisms of introduction into the Vychegda river basin
This paper presents a description of the morphological diversity of populations of white-eye bream and Chinese sleeper, which are alien to the Vychegda River basin. Their introduction occurred as a result of different mechanisms of invasion - self-settlement and acclimatization. It is shown that the white-eye bream has widely spread along the main channel of the Vychegda River, up to the upper stream. The acclimatized Chinese sleeper inhabits several fire ponds of dacha communities in the middle stream of the Vychegda River. Comparison of morphological descriptions of the white-eye bream sample with its nominative form allowed establishing a wider range of variability in the number of rays in the anal fin and rakers on the first gill arch. For the Chinese sleeper, there are also a greater number of rays in the pectoral and dorsal fins. These changes can be regarded as the result of the adaptation of the studied populations of alien species to inhabit the conditions of the acquired part of the range. Using the Zhivotovsky µ indicator, a similar level of intrapopulation morphological diversity of Chinese sleeper and white-eye bream samples was revealed. This may indicate the similarity of ongoing processes when a species moves into a new territory. However, the frequency of asymmetric manifestation per trait in the Chinese sleeper sample turned out to be high (4 points) and corresponded to the level of significant deviation from the norm. Based on the work done, a hypothesis was put forward that a population formed from a small number of individuals in a small reservoir area is more influenced by environmental factors during the period of early ontogenesis compared with species independently settle along riverbeds. It is assumed that the "founder" and "bottle neck" effects are the basis of this process.
Текст научной статьи Фенотипическое разнообразие чужеродных видов при разных механизмах вселения в бассейн р. Вычегда
Активное хозяйственное освоение водных экологических систем в XX в. привело к становлению проблемы биологических инвазий, т.е. миграции и вселению гидробионтов за пределы своих «материнских» ареалов (Биологические инвазии…, 2004; Стерлигова, Ильмаст, 2009). К антропогенно-обусловленным причинам расселения видов относятся преобразование среды обитания или изменение ее экологических характери- стик, целенаправленная интродукция и реинтродукции видов, а также случайное вселение (Биологические инвазии..., 2004).
Для водных экологических систем данная проблема наиболее актуальна, поскольку при их трансформации устраняются географические барьеры для распространения обитающих в них гидробионтов. В результате данного процесса возможно изменение состава и структуры рассматриваемых сообществ, что приводит к экологическим и экономическим последствиям (Lodge, 1993).
Одним из интересных аспектов исследования инвазионного процесса является изучение фенотипического разнообразия чужеродных видов. Известно, что внутрипо-пуляционная изменчивость количественных признаков является результатом сложной системы взаимодействия генофонда группы и условий конкретной ландшафтно-географической зоны (Кирпичников, 1987; Павлов, 2007). Сильно различающаяся численность стартовых группировок (от нескольких единиц до сотен тысяч) при разных путях инвазии, а также воздействие естественного отбора в конечном итоге формируют определенное фенотипическое разнообразие популяций чужеродных видов, которое представляет основу для их возможной дифференциации.
В бассейне реки Вычегда, являющейся одной из крупнейших рек Республики Коми, в XX–XXI вв. произошла натурализация белоглазки Ballerus sapa (Pallas, 1814). Основными источниками вселения здесь явились гидротехнические сооружения, среди которых наибольшую роль в процессе расселения сыграли каналы, соединяющие бассейны крупнейших речных систем (Новоселов, 2000).
Иной коридор инвазий представлен различными искусственными водоемами, среди которых, помимо водохранилищ, есть даже незначительные по площади пожарные водоемы и карьеры (Захаров, Бознак, 2011; Рафиков 2016). Так, например, в пожарных водоемах дачных обществ в нескольких километрах от г. Сыктывкара обнаружен ротан-головешка Perccottus glenii Dybowski, 1877, вселенный туда аквариумистами или рыбаками-любителями в 90-х годах прошлого столетия (Бознак, 2004).
Неоднородность условий формирования популяций рассматриваемых видов и разная численность их стартовых группировок позволяют нам предположить, что показатели морфологического разнообразия и стабильность развития особей в популяциях ротана-головешки и белоглазки могут отличаться.
Наблюдения за процессами инвазии дают возможность выявить происходящие изменения рыбной части сообщества крупных речных систем европейского северо-востока России, что необходимо для прогнозирования возможных экологических и экономи- ческих последствий. Изучение биологических особенностей популяций чужеродных видов, характеризующихся разными механизмами (способами) инвазии, позволит оценить процесс адаптации вида к новым условиям обитания, внести вклад в понимание причин и направленности микроэволю-ционного процесса при расселении видов.
Цель данной работы – описать фенотипическое разнообразие популяций белоглазки и ротана-головешки в пределах бассейна р. Вычегда.
Материалы
Материал, использованный для данной работы, собирался крючковыми и сетными орудиями лова в нескольких малых прудах (район расположения – N 61°40’56.8» E 50°45’48.0») на территории дачных массивов с 2012 по 2014 г., а также русловой части р. Вычегда в районе г. Сыктывкара с 2022 по 2024 г. Объем обработанного материала составил: две выборки по 30 экз. ротана-голо-вешки и 31 экз. белоглазки. Также для сравнительного анализа использована выборка леща в количестве 34 экз. из того же района отлова (рис. 1).
Площадь малых прудов, где обитает ротан, составляет от 3 до 6 га, грунт песчан-ный с наилком. Вдоль берегов произрастают лиственные и хвойные породы деревьев, а также отмечены заросли рогоза и различных видов осок. Вода имеет коричневатую окраску с прозрачностью около 0.9 м по диску Секки. По результатам гидрохимического анализа, проведенного на базе лаборатории «Экоаналит» Института биологии Коми НЦ УрО РАН, вода относится к гидрокарбонат-но-кальциевому классу 1-го типа с повышенным содержанием ионов натрия и калия. Минерализация составила до 300 мг/л, что в два раза выше, чем в пойменных водоемах бассейна среднего течения р. Вычегда. В летний период отмечено большое количество нитчатых водорослей, покрывающих собой дно водоема, а также зарастание поверхности водного зеркала исследованных прудов ряской малой Lemna minor L.
Более высокая минерализация, содержание азота и фосфора, а также интенсивное развитие водорослей свидетельствуют о происходящем антропогенном эвтрофиро-вании указанных водных объектов (Рафиков, 2018).
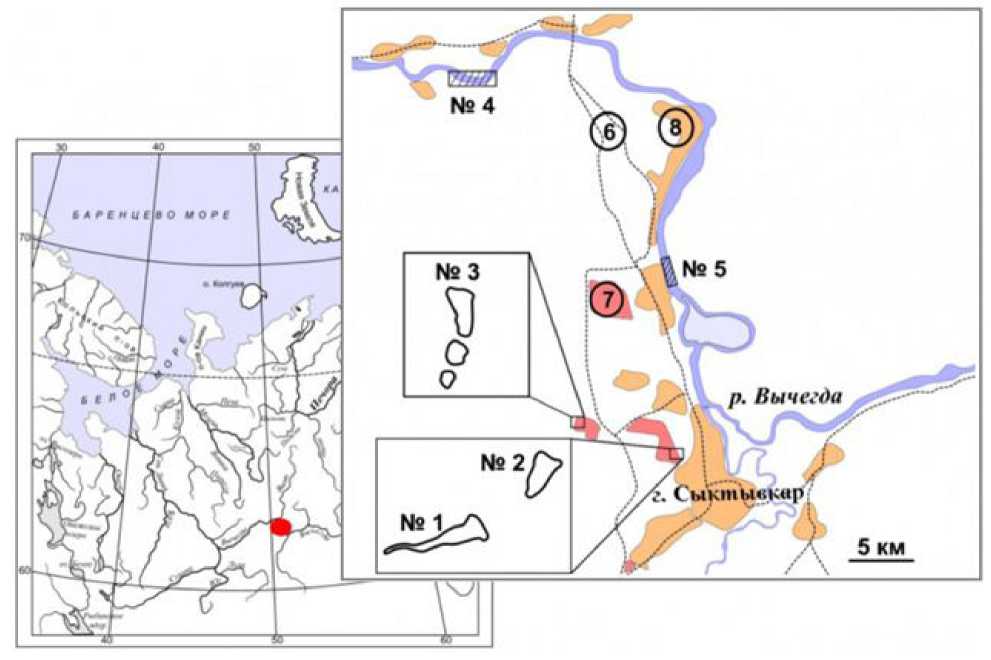
Рис. 1. Карта-схема района исследования в бассейне р. Вычегда. Условные обозначения: № 1 и 2 – места отлова ротана в дачном массиве «Дырнос», № 3 – место поимки ротана в дачном массиве «Находка», № 4–5 – места отлова белоглазки и леща в русле р. Вычегда, 6 – автодороги, 7 – дачи, 8 – населенные пункты
Fig. 1. Scematic map of the research area in the Vychegda River basin. Symbols: 1 and 2 – ponds, places of catching Chinese sleeper in the dacha community "Dyrnos", 3 – pond, place of catching Chinese sleeper in the dacha community “Nakhodka”, 4–5 – places of catching white-eye bream and eastern bream in the Vychegda River, 6 – highways, 7 – dachas, 8 – settlements
Методы
Обработку материала проводили в лабораторных условиях по стандартным методикам (Правдин, 1966), использовав для подготовки данной работы лишь группу стабильных в онтогенезе меристических признаков: ll – число прободенных чешуй в боковой линии (для ротана squma – число поперечных рядов чешуй в боку тела), D или D2, A, V, P – число ветвистых лучей соответственно в спинном (для ротана во втором спинном), анальном, брюшном и грудном плавнике, sp. br. – число тычинок на 1-й жаберной дуге, vert. – общее число позвонков. Для более точного определения количества лучей в плавниках и тычинок на первой жаберной дуге указанные структуры окрашивали ализариновым красным (Якубовский, 1970). Значения парных морфологических признаков представлены по левой стороне тела. При наличии сросшихся позвонков их количество было определено по остаткам сочленовых поверхностей и остистым отросткам, произведен перерасчет их общего количества.
Статистическая обработка данных проведена с использованием программного пакета PAST 3.25. Вследствие отсутствия полового диморфизма по всем исследованным меристическим признакам выборки представлены смешанным по половому составу материалом. Значение признаков номинативных форм представлено из монографии «Атлас пресноводных рыб России» (Атлас…, 2003).
Для оценки стабильности развития чужеродных видов проведено определение частоты асимметричного проявления на признак (Захаров и др., 2000) в билатеральных морфологических признаках ( squma или ll, P, V, sp. br. ), определен уровень морфологического разнообразия с применением коэффициента вариации, показателя ( µ ) Животов-ского и доли ( h ) редких морф (Животовский, 1980) на основании семи признаков, указанных в табл. 1 и 3.
Результаты
На фоне достаточно интенсивного рыболовства, сосредоточенного в рассматриваемом водотоке, хозяйственно ценных (сиг обыкновенный, хариус европейский) и крупных частиковых (щука, лещ) видов рыб, увеличивается промысловое значение менее ценных представителей ихтиофауны, одним из которых является расселяющаяся белоглазка. В бассейне р. Северная Двина она стала отмечаться в 1960-х гг., а в р. Вычегда – в начале 1970-х гг. (Соловкина, 1975; Новоселов, 2000). На сегодняшний день данный вид освоил русловые биотопы достаточно обширной территории бассейна р. Северная Двина, в т.ч. нижнее и среднее течения (до района с. Усть-Кулом) самого крупного притока – р. Вычегда.
Нам неизвестно, как именно (посредством аквариумистов или рыбаков-любителей) ротан-головешка был акклиматизирован в пруды дачного общества возле г. Сыктывкара, однако необходимо отметить, что по прошествии двух десятилетий спектр водоемов, где он был обнаружен, увеличился. Помимо озер № 1 и 2, где отобран материал, обнаружены особи ротана в других пожарных водоемах (№ 3) на удалении 10 км от первого места обнаружения (см. рис. 1). В иных водных объектах бассейна р. Вычегда данный вид пока не зафиксирован.
Величина и диапазон изменчивости меристических признаков белоглазки и ротана-головешки из бассейна р. Вычегда (см. табл. 1) соответствуют их видовым стандартам по большей части показателей (Атлас..., 2003). Однако необходимо отметить, что белоглазка характеризуется большим количеством лучей в анальном плавнике и тычинок на жаберной дуге. Ротан, в свою очередь, имеет большее число лучей в грудном и спинном плавниках. При анализе опубликованных данных обнаружены разнонаправленные значимые отличия с указанными выборками. Например, ротан из водоемов Республики Коми значимо отличается ( Tst = 4 при p < 0.05) меньшим количеством ветвистых лучей во втором спинном и анальном ( Tst = 7.5 при p < 0.05) плавниках по сравнению с нативной популяцией из района г. Хабаровска. Однако какая-либо географическая обусловленность (зависимость от широты местности) этих изменений не прослеживается.
Результаты, полученные на основании морфологических описаний одних и тех же признаков (указаны в табл. 1 и 3), свидетельствуют о том, что популяционное разнообразие белоглазки (µ = 3.61 ± 0.11 и h = 0.14 ± 0.02) и ротана (µ = 3.44 ± 0.12 и 3.45 ± 0.09; h = 0.15 ± 0.03 и 0.09 ± 0.02) достоверно не различаются. В то же время показатель частоты асимметричного проявления на признак и его балльная оценка для ротана-головешки указывают на существенные отклонения стабильности индивидуального развития его выборок от нормы (табл. 2).
Одним из факторов успешной натурализации вселенцев является их филогенетическое сходство с аборигенными видами рыб (Xu et al., 2024). Так, например, для белоглазки, активно расселяющейся по магистральному руслу р. Вычегда, можно указать близкородственный вид – лещ Abramis brama (Linnaeus, 1758). Это довольно широко распространенный и массовый вид в бассейне изучаемой реки. Морфологическое разнообразие его выборки оказалось выше (3.93 ± 0.11, Tst = 2.1 при p < 0.05), чем у белоглазки, при отсутствии значимых различий в уровне асимметрии.
Наибольший вклад в показатель общей морфологической изменчивости леща и белоглазки внесли одни и те же морфологические структуры, среди которых: число пробо-денных чешуй в боку тела, ветвистых лучей в анальном плавнике и количество жаберных тычинок (см. табл. 3).
Обсуждение
Рассматриваемые виды рыб очень отличаются по своим предпочтениям к главным экологическим характеристикам, что подтверждается принадлежностью к разным фаунистическим комплексам рыб (Никольский, 1980). Так, например, белоглазка относится к понтокаспийскому ихтиокомплексу, а ротан-головешка – к китайскому равнинному. Однако, несмотря на то, что основная черта представителей понтокаспиского их-тиокомплекса – это теплолюбивость, близкое филогенетическое сходство белоглазки с широко распространенным в данном бассейне лещом может свидетельствовать о ее предрасположенности к расселению в условиях Севера (Xu et al., 2024). На сегодняшний день белоглазка расселилась по руслу Северной Двины и вплоть до верхнего течения одного из крупных притоков – р. Вычегда.
Представители китайского равнинного комплекса имеют высокий инвазивный потенциал, что позволило им расселиться по всей России, сопредельным странам и не только (Решетникав, 2009; Интересова, 2016;
Таблица 2. Значения показателей морфологического разнообразия и флуктуирующей асимметрии исследованных выборок белоглазки, леща и ротана-головешки
|
Вид |
Число особей |
ЧАП/П |
Балл |
Показатель Животовского ( µ ) |
Доля редких морф ( h ) |
|
Белоглазка |
31 |
0.26 ± 0.03 |
1 |
3.61 ± 0.11 |
0.14 ± 0.02 |
|
Лещ |
34 |
0.30 ± 0.04 |
2 |
3.93 ± 0.11 |
0.13 ± 0.02 |
|
Ротан озеро № 1 |
30 |
0.36 ± 0.04 |
3 |
3.45 ± 0.09 |
0.09 ± 0.02 |
|
Ротан озеро № 2 |
30 |
0.40 ± 0.04 |
4 |
3.44 ± 0.12 |
0.15 ± 0.03 |
Примечание. ЧАП/П - частота асимметричного проявления на признак, балл - балльная оценка показателя асимметрии.
Таблица 3. Основные показатели изменчивости меристических признаков белоглазки и леща из бассейна р. Вычегда
|
Признак |
Белоглазка (n = 31) |
Лещ ( n = 34) |
||||||||
|
min–max |
X ± x |
СV |
m |
µ |
min–max |
X ± x |
СV |
m |
µ |
|
|
ll |
49–54 |
51.8 ± 0.2 |
2.3 |
6 |
5.3 ± 0.4 |
52–60 |
55.4 ± 0.4 |
3.1 |
9 |
8.2 ± 0.5 |
|
D |
8 |
8 |
0 |
1 |
1.0 ± 0.0 |
9–10 |
9.1 ± 0.1 |
3.2 |
2 |
1.6 ± 0.1 |
|
A |
36–43 |
40.0 ± 0.3 |
4.5 |
8 |
7.3 ± 0.4 |
23–28 |
25.6 ± 0.2 |
5.3 |
6 |
5.7 ± 0.2 |
|
V |
7–8 |
8.0 ± 0.1 |
2.3 |
2 |
1.4 ± 0.2 |
8–9 |
8.0 ± 0.1 |
2.2 |
2 |
1.3 ± 0.2 |
|
P |
16–18 |
16.7 ± 0.1 |
3.4 |
3 |
2.6 ± 0.2 |
16–17 |
16.5 ± 0.1 |
3.1 |
2 |
2.0 ± 0.0 |
|
sp. br . |
20–25 |
22.2 ± 0.2 |
5.9 |
6 |
5.4 ± 0.3 |
21–27 |
24.0 ± 0.3 |
6.2 |
7 |
6.1 ± 0.4 |
|
vert . |
45–47 |
46.4 ± 0.1 |
1.2 |
3 |
2.5 ± 0.2 |
43–45 |
44.1 ± 0.1 |
1.3 |
3 |
2.7 ± 0.2 |
|
Уровень морфологического разнообразия, рассчитанный по выборке ( µ ) |
3.6 ± 0.1 |
3.9 ± 0.1 |
||||||||
Примечание. min–max – пределы варьирования, x ± xm – среднее значение признака ± его ошибка, СV – коэффициент вариации, m – количество вариантов (морф), µ – показатель Животовского.
ских факторов, что затрудняет поддержание гомеостаза развития особей. Данное предположение может объяснять высокие значения асимметрии в выборках ротана-голо-вешки (4 балла по 5-балльной шкале – значительный уровень отклонений от нормы). Ранее были описаны различные фенодиви-ации (аномалии позвоночного столба и лучей в плавниках) у исследованных выборок ротана. Так, например, у особей из пруда № 2 до 57 % (17 особей из 30 экз.) от выборки имели компрессии позвонков, включающее срастание позвонков в количестве от 1 до 3 случаев на 1 особь. Количество аномалий позвоночника в выборке из пруда № 1 составила до 20 %, или 6 особей из 30 экз. (Рафиков, 2018). Для чужеродных популяций ротана уже описаны случаи большого числа аномалий (до 40 % выборки) в строении плавников (Шемонаев, Кириленко, 2013). Показан высокий уровень флуктуирующей асимметрии (0.35–0.42, что соответствует 3–4 баллам), рассчитанной по единой методике с применением тех же признаков для популяций ротана из некоторых водоемов Тюменской области (Бойченко, 2020).
В качестве альтернативной причины качественных и количественных изменений фенотипических характеристик указанных видов можно предположить неравнозначный характер отбора в популяциях видов-вселенцев, находящихся в разных условиях. При расселении в естественном водном объекте чужеродный вид встраивается в систему отношений в сообществе, как и аборигенные виды, в частности отношения по типу «хищник – жертва». В изолированных водных объектах может не быть естественных хищников, которые бы нивелировали особей с дефектами развития.
После накопления морфологических описаний других чужеродных видов рыб, оби-
