Генетический состав носителей тагарской культуры: современное состояние и перспективы исследований

Автор: Черданцев С.В., Трапезов Р.О., Пристяжнюк М.С., Томилин М.А., Журавлев А.А., Пилипенко А.С.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология эпохи палеометалла средневековья и нового времени
Статья в выпуске: т.XXVII, 2021 года.
Бесплатный доступ
Несмотря на высокую степень исследованности тагарской культуры и ее носителей методами археологии и физической антропологии, многие направления изучения этой группы населения Южной Сибири раннего железного века остаются актуальными. Данная работа посвящена рассмотрению результатов исследования генетического состава носителей тагарской культуры методами палеогенетики, полученных к настоящему моменту. Проведенное авторами исследование репрезентативной серии образцов мтДНК от представителей разновременных групп тагарского населения (N=79) впервые позволило сформировать объективные представления о составе митохондриального генофонда этой популяции. Полученные результаты хорошо иллюстрируют необходимость исследования численно репрезентативных серий образцов при выполнении палеогенетических реконструкций. Для тагарской популяции в целом характерно присутствие в генофонде как западно-евразийских (H, HV6, HV*, I, K, T, U2e, U4, U5a и U*), так и восточно-евразийских (A*, A8, C*, C5, D, G2a, F1b) гаплогрупп мтДНК. В отличие от других популяций Южной Сибири скифского времени, в тагарском генофонде доминируют западно-евразийские варианты. Среди предшествующих популяций тагарская популяция демонстрирует близость с популяциями Минусинской котловины эпохи развитой бронзы. При сравнении с группами ранних кочевников Евразии тагарское население проявляет близость в отношении структуры генофонда мтДНК с носителями других культур скифского круга, занимая промежуточное положение между географически удаленными скифами Северного Причерноморья и населением южносибирских культур - пазырыкской и алды-бельской. При переходе от подгорновского к сарагашенскому этапам происходит сближение тагарской популяции с другими группами ранних кочевников Южной Сибири. В тагарском генофонде Y-хромосомы выявлено доминирование вариантов гаплогруппы R1a1a, которая была впервые привнесена в Минусинскую котловину носителями андроновской (фёдоровской) культуры. Однако для уточнения происхождения тагарских вариантов Y-хромосомы требуется их углубленный анализ. В работе приведены наиболее перспективные направления дальнейшего исследования тагарской популяциями методами палеогенетики.
Тагарская культура, ранний железный век, палеогенетика, митохондриальная днк, y-хромосома, генетическая история популяций
Короткий адрес: https://sciup.org/145146097
IDR: 145146097 | УДК: 903.5 | DOI: 10.17746/2658-6193.2021.27.0723-0729
Genetic composition of the tagar culture population: current state and research prospects
Despite the high degree of archaeological and anthropological research of the Tagar culture and its carriers, many directions of study of this Sothern Siberian Iron Age population remain relevant. This work is devoted to the consideration of the data on the genetic composition of the Tagar culture population, obtained to date by paleogenetic methods. The study of representative mtDNA sample series from chronologically different groups of the Tagar population (N = 79), carried out by the authors of this work, allow us to obtain the first objective view on the composition of its mitochondrial gene pool. The obtained results illustrate well the significance of analysis of representative sample series when performing paleogenetic reconstructions. The general Tagar population is characterized by the presence in the gene pool of both Western Eurasian (H, HV6, HV*, I, K, T, U2e, U4, U5a, and U*) and Eastern Eurasian (A*, A8, C*, C5, D, G2a, Fib) mtDNA haplogroups. Unlike other populations of southern Siberia of the Scythian time, Western Eurasian variants dominate in the Tagar gene pool. Among the previous populations, the Tagar population demonstrates affinity with the populations of the Minusinsk Basin of the Middle Bronze Age. When compared with the groups of early nomads of Eurasia, the Tagar population shows a similarity in terms of the mtDNA variability with the carriers of other cultures of the Scythian circle, occupying an intermediate position between the geographically distant Scythians of the Northern Black Sea region and the population of the south Siberian cultures, Pazyryk and Aldy-Bel. During the transition from the Podgorny to the Saragashen stages, the Tagar population converges with other groups of early nomads of southern Siberia. In the Tagar gene pool of the Y chromosome, the dominance of variants of the Riaia haplogroup was revealed. This genetic cluster was first introduced into the Minusinsk Basin by the carriers of the Andronovo (Fedorovo) culture. However, additional in-depth analysis of the Tagar Y chromosome lineages is required to clarify their origin. The paper presents the most promising directions for further research of the Tagar populations using paleogenetic methods.
Текст научной статьи Генетический состав носителей тагарской культуры: современное состояние и перспективы исследований
Тагарская культура Минусинской котловины относится к числу наиболее хорошо исследованных археологических культур раннего железного века на территории Южной Сибири. Археологами проведены раскопки тысяч погребальных комплексов, накоплен обширный объем данных об особенностях различных компонентов материальной культуры тагарского населения, предложены варианты периодизации этапов развития культуры [Грязнов и др., 1979; Вадецкая, 1986]. Параллельно с этим сформированы репрезентативные палеоантропологические коллекции, анализ которых позволил охарактеризовать тагарскую популяцию (по крайней мере, некоторые из локальных групп населения) методами палеоантропологии [Козинцев, 1977; Учанева и др., 2017]. Все эти направления исследований (включая раскопки новых памятников, анализ инвентаря, уточнение хронологии и периодизации тагарской культуры, характеристика популяций методами антропологии) сохраняют свою актуальность и на данный момент. На фоне традиционных направлений уровень анализа генофонда носителей тагарской культуры методами палеогенетики оставался недостаточным. Хотя население Алтае-Саянской горной системы раннего железного века стало одним из первых объектов применения методов палеогенетики на территории России, все ранние работы были сфокусированы на генетическом анализе носителей пазырыкской культуры Горного Алтая (см., напр.: [Молодин и др., 2003]). Небольшая серия носителей тагарской культуры была включена в состав низкорепрезентативной диахронной выборки населения Минусинской котловины эпохи развитой бронзы и раннего железного века, что стало первым этапом исследования генофонда тагарской популяции [Keyser et al., 2009]. Позже были опубликованы результаты анализа мтДНК еще нескольких тагарских образцов, происходивших из памятника Барсучий лог [Unterländer et al., 2017], но и эта работа не была сфокусирована на собственно тагарской популяции: тагарские образцы были включены в состав обширной по географическому охвату и разнообразной в отношении культурной атрибуции выборки населения степной полосы Евразии скифского времени. Подробный анализ тагарских материалов в этой работе отсутствует, а среди южносибирских материалов фокус оставался на анализе носителей пазырыкской культуры [Там же].
Целенаправленные исследования структуры генофонда носителей тагарской культуры впервые были выполнены авторами данной работы. Их первым этапом стал анализ репрезентативной серии образцов мтДНК представителей разновременных групп тагарского населения [Pilipenko et al., 2018]. Для работы были использованы палеоантропологические материалы, полученные Красноярской археологической экспедицией Ленинградского отделения Института археологии АН СССР в 1955–1970 гг. под руководством М.П. Грязнова и находящиеся в настоящее время в составе палеоантропологической коллекции ИАЭТ СО РАН (г. Новосибирск). Всего в работе были использованы палеоантропологические образцы от 95 носителей тагарской культуры из более чем десяти могильников, расположенных относительно компактно в среднем течении Енисея вблизи современной границы Красноярского края и Хакасии (Барсучиха-1, -6, -7, Кичик-Кюзюр, Новая черная-1, Перевозин-ский Чаатас, Подгорное озеро, Советская Хакасия, Тепсей-8, -9, Улуг-Кюзюр).
Работы были выполнены на базе палеогене-тической инфраструктуры ИЦиГ СО РАН (г. Новосибирск). Методы предварительной обработки материала, получения образцов ДНК, анализа структуры мтДНК и статистиче ской обработки полученных данных подробно описаны в нашей статье [Там же].
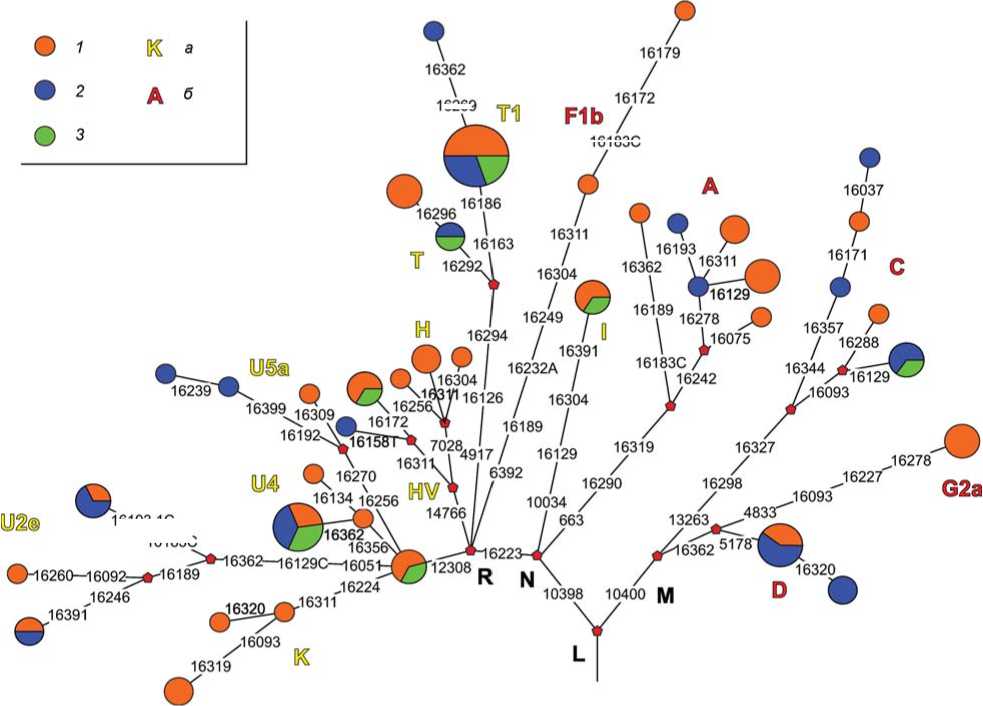
Для 79 из 95 образцов первоначальной серии были получены достоверные данные по структуре мтДНК. В составе этой суммарной выборки присутствовали представители всех основных этапов тагарской культуры (здесь и далее мы использовали периодизацию этапов, предложенную М.П. Грязновым), включая «раннетагарскую» серию численностью 42 образца (почти вся серия – под-горновские материалы), сарагашенский этап (24 образца) и наименее численно репрезентативный тесинский этап (9 образцов). Для всех 79 образцов было определено филогенетическое положение, что позволило объединить их в дерево, отражающее разнообразие и филогенетические отношения образцов из состава исследованной серии (рис. 1).
Дальнейший анализ полученных результатов выполнен как при рассмотрении серии в целом, так и подгрупп образцов, представляющих различные этапы существования тагарской культуры. Среди 79 успешно исследованных образцов мы выявили 39 структурных вариантов мтДНК. Для тагарского населения в целом, как и для всех исследованных к настоящему моменту популяций раннего железного века Южной Сибири, было характерно присутствие в генофонде как западно-евразийских (H, HV6, HV*, I, K, T, U2e, U4, U5a и U*), так и восточно-евразийских (A*, A8, C*, C5, D, G2a, F1b) гапло-групп мтДНК. При этом тагарское население отличается от других групп ранних кочевников Южной Сибири (носители пазырыкской и алды-бельской культур) доминированием в генофонде западно-евразийских вариантов мтДНК (64.6 %), в то время как для других скифо-сибирских групп характерно присутствие в генофонде мтДНК не менее 50 % восточно-евразийских вариантов.
Сравнительное распределение вариантов мтДНК по гаплогруппам в ранее исследованной суммарной серии (16 образцов суммарно из работ [Keyser et al., 2009; Unterländer et al., 2017] и в серии, исследованной в нашей работе (79 образцов) показано на рис. 2. Более 74 % выявленных нами вариантов мтДНК отсутствовали в серии ранее
16183С
Рис. 1. Филогенетическое дерево вариантов мтДНК, выявленных в исследованной нами тагарской серии [Pilipenko et al., 2018].
Принадлежность образцов к этапам тагарской культуры: 1 – раннетагарские (преимущественно подгорновский этап); 2 – сарагашенский этап; 3 – тесинский этап.
Гаплогруппы мтДНК: а – западно-евразийские; б – восточно-евразийские.
16193ЛС
16183С
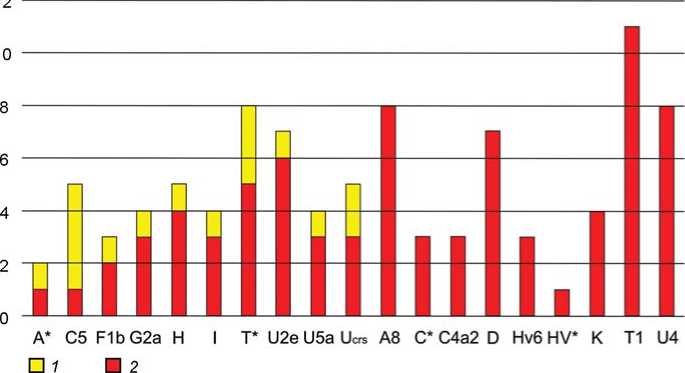
Рис. 2. Состав и соотношение гаплогрупп мтДНК в сериях тагарских образцов, опубликованных к настоящему моменту.
1 – образцы (N=16) из [Keyser et al., 2009; Unterländer et al., 2017]; 2 – образцы, исследованные авторами (N=79) [Pilipenko et al., 2018].
исследованных образцов. Более того, в ранее опубликованных данных полностью отсутствовали гаплогруппы мтДНК, охватывающие почти 60 % исследованной нами репрезентативной выборки. Это еще одна иллюстрация необходимости анализа репрезентативных по численности серий образцов при исследовании структуры генофонда древних популяций, особенно на уровне маркеров с однородительским типом наследования (мтДНК и Y-хромосома). В то же время, численность исследованной нами серии достаточна для обнаружения всех компонентов генофонда (гаплогрупп), присутствующих в генофонде общей тагарской популяции с частотой более 3.7 % (подробную статистику см. в [Pilipenko et al., 2018]), то есть нам с высокой вероятностью известны все основные компоненты генофонда мтДНК этой популяции.
Сравнительный анализ исследованной нами тагарской с ерии с опубликованными сериями мтДНК из других популяций Евразии как предшествующих эпох (неолит и различные периоды эпохи бронзы), так и с группами ранних кочевников скифского и гунно-сарматского времени показал, что наибольшую близость (т.е. наименьшее генетическое расстояние) тагарское население демонстрирует с носителями различных культур, отно сящихся к так называемому скифскому кругу. Интересно, что среди этих популяций ранних кочевников суммарная тагарская серия занимает промежуточное положение между географически соседствующими с ней пазырыкцами и носителями алды-бельской культуры Тывы, с одной стороны, и классическими скифами Северного Причерноморья, с другой [Pilipenko et al., 2018]. Причем генетическое расстояние со скифами Северного 726
Причерноморья даже меньше, чем с южно-сибирскими ранними кочевниками. Среди популяций предшествующих эпох суммарная тагарская серия демонстрирует наименьшее расстояние с популяциями эпохи развитой бронзы непосредственно из Минусинской котловины, хотя генетическое расстояние с любой из популяций эпохи бронзы больше, чем с ранними кочевниками скифского времени. При этом тагарская серия занимает промежуточное положение между окуневской и ан-дроновской сериями из Минусинской котловины [Там же]. Следует подчеркнуть, что на данный момент далеко не для всех этнокультурных групп эпохи бронзы и раннего железного века, известных археологам и антропологам, получены репрезентативные палеогенетические данные, которые позволили бы проводить статистиче ски до стоверное сравнение с тагарской серией. По мере появления новых данных межпопуляционный сравнительный анализ может быть существенно расширен.
Интересен результат сравнительного анализа серий, соответствующих отдельным этапам развития тагарской культуры, между собой и с другими популяциями ранних кочевников Евразии [Там же]. Серия, соответствующая ранним этапам тагарской культуры (преимущественно, подгорновский этап) оказалась наиболее близка классическим скифам Северного Причерноморья. Серия сарагашенского этапа существенно сближается уже с популяциями пазырыкской и алды-бельской культур Южной Сибири, по-видимому, за счет вовлечения в состав генофонда автохтонных для региона компонентов, преимущественно восточно-евразийского происхождения (именно их доля в сарагашенской серии существенно возрастает). Несколько неожиданным выглядит положение тесинской серии, полностью состоящей из компонентов западно-евразийского происхождения (локализуется среди популяций западной части степного пояса). Однако следует подчеркнуть, что тесинская серия, численностью всего 9 образцов, пока не может быть признана репрезентативной по отношению к популяции носителей тесинского этапа, и наблюдаемая нами картина может объясняться случайными причинами (эффектом малой выборки). Хотя сам факт, что ни один из девяти исследованных тесинских образцов не принадлежит к восточно-евразийскому кластеру гаплогрупп мтДНК, вызывает интерес уже на данном этапе исследования.
Полученные результаты позволяют нам предварительно констатировать, что структура генофонда мтДНК тагарского населения могла сформироваться на основе западно-евразийских компонентов, многие из которых были привнесены на территорию Минусинской котловины еще в эпоху бронзы (в частности, с волной миграции носителей ан-дроновской (фёдоровской) культуры), при участии автохтонных для региона компонентов восточноевразийского происхождения. При этом доля автохтонных южносибирских (и/или центральноазиатских) компонентов в генофонде продолжала увеличиваться при переходе от ранних (подгорнов-ского) к более поздним (сарагашенскому) этапам развития культуры.
Следующим важным этапом палеогенетиче-ского исследования тагарского населения, который продолжается по настоящее время, является анализ структуры мужского генофонда носителей тагарской культуры. К настоящему времени нами исследована серия Y-хромосом тагарского населения численностью более 40 образцов. Более двух третей исследованной серии происходят из комплексов подгорновского этапа, оставшиеся – из сарагашенских комплексов. Разнообразие Y-хромосомы тагарского населения резко контрастирует с мтДНК. Если генофонд мтДНК является смешанным, с большим числом филогенетически и филогеографически контрастных компонентов, то мужской генофонд тагарского населения однообразен. В его составе доминирует один филогенетический кластер Y-хромосомы – гаплогруппа R1a1a. Варианты этого кластера впервые проникают в Минусинскую котловину (и на сопредельные территории юга Сибири) в первой половине II тыс. до н.э. с волной мигрантов-носителей ан-дроновской (фёдоровской) культуры. Варианты гаплогруппы R1a1a составляют основу андронов-ского мужского генофонда как в Минусинской котловине, так и на территории Верхнего Приобья.
Таким образом, структура мужского генофонда тагарского населения (по крайней мере, подгор-новского и сарагашенского этапов) сближает его с пришлым андроновским населением эпохи развитой бронзы. Одновременно, эта черта мужского генофонда отличает тагарскую популяцию от других южносибирских ранних кочевников, в частности, от носителей пазырыкской культуры, для которых, судя по предварительным данным, было характерно высокое разнообразие вариантов в генофонде Y-хромосомы. Следует подчеркнуть, что имеющиеся к настоящему моменту данные не позволяют однозначно констатировать андроновское (фёдоровское) происхождение доминирующего компонента мужского генофонда носителей та-гарской культуры. Дело в том, что в составе гапло-группы R1a1a выделяется несколько субкластеров, далеко не все из которых можно связывать с ан-дроновской (фёдоровской) популяцией. Для прояснения этого аспекта требуется более подробный анализ структуры образцов Y-хромосомы, относящихся к гаплогруппе R1a1a, как из андроновских, так и их тагарских мужских серий. Это углубленное исследование уже осуществляется нашей лабораторией.
Таким образом, на данном этапе тагарское население Минусинской котловины в относительно высокой степени уже охарактеризовано с точки зрения генофонда мтДНК и Y-хромосомы. Однако, решение многих вопросов, связанных с генетической историей этой популяции, зависит от продолжения палеогенетических работ. Основными актуальными и перспективными направлениями исследований генофонда носителей тагарской культуры, на наш взгляд, следует считать следующие.
-
1. Исследование репрезентативных серий образцов ДНК, отно сящихся к отдельным этапам тагарской культуры. Прежде всего, это относится к исследованию населения тесинского этапа, для которого отсутствуют репрезентативные данные по мтДНК и Y-хромосоме. При исследовании тесинских материалов крайне важным является представленность в составе серий образцов, происходящих из комплексов с различными типами погребальной обрядности, и сравнительный анализ между собой этих археологически контрастных серий тесинского этапа. В целом исследования генофонда носителей различных этапов (включая переходные) тагарской культуры следует координировать с процессами уточнения периодизации и хронологии тагарской культуры [Савинов, 2011].
-
2. Существенное увеличение серий Y-хромосомы для всех групп тагарского населения, а также углубленный анализ образцов Y-хромосомы, принадлежащих к кластеру R1a1a.
-
3. Проведение исследований серий образцов ДНК из локальных групп тагарского населения, все еще не охваченных палеогенетическими исследованиями с целью прояснения вопросов локально-территориальной дифференцированности тагарского населения в пределах общего ареала культуры. Особые перспективы для развития данного направления открываются, в частности, в связи с проводимыми в последние годы масштабными спасательными археологиче ски раскопками на территории Хакасии, в результате которых исследованы многочисленные разновременные та-гарские памятники, в частности, на территории Аскизского р-на.
-
4. Разработка вопросов, связанных со степенью родства носителей тагарской культуры из состава коллективных погребений и склепов.
-
5. С методологической точки зрения представляется актуальным привлечение к анализу тагар-ских материалов методов полногеномного анализа аутосомных маркеров. Все более широкая доступность высокопроизводительного секвенирования позволяет рассчитывать на проведение в ближайшие годы полноценных полногеномных исследований репрезентативных серий образцов от носителей тагарской культуры.
-
6. Еще одним перспективным направлением исследований, не связанным напрямую с анализом непосредственно тагарских материалов, является накопление репрезентативных палеогенетических данных о предшествующих популяциях региона (включая носителей карасукской культуры), а также о группах, сменивших тагарское население на территории Минусинской котловины (включая носителей таштыкской культуры). Это позволит в перспективе полноценно использовать один из наиболее информативных подходов к реконструкции генетической истории населения любого региона – анализ репрезентативной диахронной выборки популяций.
Работа выполнена в рамках гранта РНФ № 17-7820193. Работа частично профинансирована из средств государственного задания ИЦиГ СО РАН, № 02592019-0010.
Список литературы Генетический состав носителей тагарской культуры: современное состояние и перспективы исследований
- Вадецкая Э.Б. Археологические памятники в степях Среднего Енисея. - Л.: Наука, 1986. - 180 с.
- Грязнов М.П., Завитухина М.П., Комарова М.Н., Миняев С.С., Пшеницына М.Н., Худяков Ю.С. Комплекс археологических памятников у горы Тепсей на Енисее. - Новосибирск: Наука, 1979. - 167 с.
- Козинцев А.Г. Антропологический состав и происхождение населения тагарской культуры. - Л.: Наука, 1977. -144 с.
- Молодин В.И., Воевода М.И., Чикишева Т. А., Ромащенко А.Г., Полосьмак Н.В., Шульгина Е.О., Нефедова М.В., Куликов И.В., Дамба Л.Д., Губина М.А., Кобзев В.Ф. Население Горного Алтая в эпоху раннего железного века как этнокультурный феномен: происхождение, генезис, исторические судьбы (по данным археологии, антропологии, генетики). - Новосибирск: Изд-во СО РАН, 2003. - 286 с.
- Савинов Д.Г. Проблема хронологии и периодизации тагарской культуры в историческом контексте // "Terra Scythica": материалы международного симпозиума "Terra Scythica". - Новосибарск: Изд-во ИАЭТ СО РАН, 2011. - С. 208-217.
- Учанева Е.Н., Казарницкий А.А., Громов А.В., Лазаретова Н.И. Население Минусинской котловины в раннем железном веке: к вопросу о внутригрупповой и межгрупповой изменчивости // Вестник археологии, антропологии и этнографии. - 2017. - №1. - С. 78-87.
- Keyser C., Bouakaze C., Crubezy E., Nikolaev V.G., Montagnon D., Reis T., Ludes B. Ancient DNA provides new insights into the history of south Siberian Kurgan people // Human Genetics. - 2009. - Vol. 126. - P. 395-410.
- Pilipenko A.S., Trapezov R.O., Cherdantsev S.V., Babenko V.N., Nesterova M.S., Pozdnyakov D.V., Molodin V.I., Polosmak N.V. Maternal genetic features of the Iron Age Tagar population from Southern Siberia (1st millennium BC) // PLoS ONE. - 2018. - Vol. 13. - Iss. 9. -P. 1-24.
- Unterländer M., Palstra F., Lazaridis I., Pilipenko A., Hofmanova Z., Groß M., Sell C., Blöcher J., Kirsanow K.,Rohland N., Rieger B., Kaiser E., Schier W., Pozdniakov D., Khokhlov A., Georges M., Wilde S., Powell A., Heyer E., Currat M., Reich D., Samashev Z., Parzinger H., Molodin V., Burger J. Ancestry, demography, and descendants of Iron Age nomads of the Eurasian Steppe // Nature Communications. - 2017. - Article number: 14615. - P. 1-10.
