Генетическое разнообразие древних народов Кавказа и сопредельной степи в эпоху энеолита - бронзы (V-II тыс. до н. э.): основные результаты и проблемы культурно-исторической интерпретации
: основные результаты и проблемы культурно-исторической интерпретации")
Автор: Трифонов В.А., Прохорчук Е.Б., Жур К.В.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Статья в выпуске: 262, 2021 года.
Бесплатный доступ
В статье представлен обзор новейших результатов анализа ДНК древних народов Кавказа и сопредельной степи в эпоху энеолита - бронзы (V-II тыс. до н. э.) и рассматриваются проблемы их культурно-исторической интерпретации. Выделено семь особенностей общей палеогенетической картины региона: 1. Формирование генотипа кавказских охотников-собирателей не позднее XII тыс. до н. э. и его раннее распространение за пределы Кавказа; 2. Существование на Кавказе непрерывной генетической преемственности с эпохи верхнего палеолита; 3. Раннее, не позднее VI тыс. до н. э., появление кавказского и связанного с ним переднеазиатского (иранского и анатолийского) генетического компонента в степном генетическом пуле; 4. Сохранение генетических различий между населением Кавказа и открытой степи на протяжении всей эпохи энеолита - бронзы; 5. Существование вдоль северных склонов Кавказа генетической буферной зоны, которую, в зависимости от исторических и климатических условий, занимали степняки или кавказцы; 6. Односторонняя проводимость потока генов через Кавказ только в направлении с юга на север; 7. Преобладание культурного разнообразия над генетическим. Отмечается, что проблемы культурно-исторической трактовки генетических данных связаны в разной степени с несогласованностью между генетической и археологической таксономиями, недооценкой генетиками археологического контекста и наивной интерпретацией археологами различий в ДНК как культурных атрибутов.
Древняя днк, археологические культуры, кавказ, степь, эпоха энеолита - бронзы
Короткий адрес: https://sciup.org/143176023
IDR: 143176023 | DOI: 10.25681/IARAS.0130-2620.262
Genetic diversity of ancient populations in the Caucasus and the adjacent steppes during the Eneolithic-Bronze Age (V-II mill. bc): main results and issues of cultural and historical interpretation
The paper provides an overview of the recent DNA analysis of the ancient populations of the Caucasus and the adjacent steppes during the Eneolithic-Bronze Age (V-II mill. BC) and explores the issues of cultural and historical interpretation of the results. Seven specific features of the general paleogenetic picture of the region have been singled out: 1. Development of the genotype of the Caucasian hunters-gatherers not later than 12 000 years ago and its early spread beyond the Caucasus; 2. Existence of genetic continuity in the Caucasus starting from the Upper Paleolithic; 3. Early appearance of the Caucasus and the related Near East (Iranian and Anatolian) genetic component in the steppe genetic pool, not later than VI mill. BC; 4. Preservation of genetic differences between the population of the Caucasus and the steppe population throughout the entire Eneolithic-Bronze Age; 5. Existence of a genetic buffer zone along the northern slopes of the Caucasus occupied either by the steppe population or the Caucasian population depending on historical and climatic conditions; 6. One-way gene flow across the Caucasus only from the south to the north; 7. Predominance of cultural diversity over genetic diversity. It is emphasized that the issues of the cultural and historical interpretation of genetic data are related, in varying degrees, to inconsistency between genetic and archaeological taxonomies, overlooking of the archaeological context by genetic scientists and naive interpretation of differences in DNA as cultural attributes by archaeologists.
Текст научной статьи Генетическое разнообразие древних народов Кавказа и сопредельной степи в эпоху энеолита - бронзы (V-II тыс. до н. э.): основные результаты и проблемы культурно-исторической интерпретации
За последние три десятилетия прогресс палеогенетических исследований ( Hadelberg et al. , 2015) революционизировал аналитические возможности археологии, одновременно предъявляя более высокие требования к навыкам культурно-исторической интерпретации результатов анализа древней ДНК ( Lebrasseur et al. , 2018). Сегодня палеогенетическому анализу могут быть подвергнуты сохранившиеся фрагменты ДНК древних биологических объектов возрастом до 700 тыс. лет ( Orlando, Ginolhac, Zhang et al. , 2013). Их выбор почти безграничен – от останков людей, животных и растений до бактерий и вирусов ( Jensen, Niemann, Højholt-Iversen et al. , 2019).
Молекулярные технологии секвенирования нового поколения ( Voelkerding et al. , 2009) и предварительной оценки сохранности ДНК в археологических образцах ( Rohland et al. , 2015) вместе с методикой выделения древней ДНК из каменистой части височной кости (костей внутреннего уха), сохраняющей даже в теплом климате до 100 раз больше ДНК, чем в остальных фрагментах скелета человека ( Gamba, Jones, Teasdale et al. , 2014; Pinhasi et al. , 2019; Sirak, Fernandes, Cheronet et al. , 2020) позволили сократить время и затраты на выделение древней ДНК почти до стандартов массового производства. В итоге были созданы технические условия для перехода от анализа единичных образцов исключительной научной ценности (неандертальцы, денисовцы) ( Krings et al. , 1997) к серийным анализам и изучению генетического профиля древних популяций людей, доместицированных животных и растений ( Haak, Forster, Bramanti et al. , 2005; Scheu et al. , 2015; Allaby et al. , 2015).
В сущности, результаты анализов древней ДНК формируют новую область знания о прежде неизвестных генетических аспектах культурных, экономических и социальных процессов в их историческом развитии. Другими словами, палеогенетика открывает для археологии новые возможности анализа истории взаимосвязей биологической природы человека с его культурной сущностью.
Настоящей демонстрацией мощи нового инструмента стала генетическая ревизия двух наиболее значимых гипотез в доисторической археологии Старого Света: предположений о массовых миграциях, сопутствующих неолитизации Европы и распространению индоевропейских языков. Возникшие в разное время независимо друг от друга, эти гипотезы были объединены К. Ренфру в одну чрезвычайно влиятельную теорию ( Renfrew , 1990), которую сам автор раньше других рассчитывал подтвердить данными генетических анализов ( Renfrew , 2000). Едва ли К. Ренфру предполагал, что спустя десятилетие археогенетика, как он назвал это направление исследований, будет располагать данными ( Haak, Lazaridis, Patterson et al ., 2015; Allentoft, Sikora, Sjögren et al. , 2015), истолкованными в пользу главного конкурента его идеи – курганной или степной теории индоевропейской прародины ( Gimbutas , 1970; Mallory , 1989; Anthon y, 2007). Между тем результаты палеогенетических анализов за последние пять лет превосходят объяснительные возможности обеих теорий и едва ли могут безоговорочно рассматриваться как доказательство или опровержение одной из них.
В значительной степени это связано с открытием почти 50 % генетического вклада кавказского происхождения в евразийский степной генетический пул эпохи энеолита – бронзы (Jones, Gonzalez-Fortes, Connell et al., 2015). Это неожиданное обстоятельство меняет не только условия решения индоевропейской проблемы, но и формирует совершенно новый контекст анализа культурных контактов древнего населения Кавказа и сопредельных территорий в доисторическую эпоху.
В идеале, чтобы избежать тенденциозности в интерпретации результатов па-леогенетических анализов, они должны быть сгруппированы независимо от археологической номенклатуры до начала проверки связей между культурными комплексами и генетическими кластерами ( Fu, Posth, Hajdinjak et al. , 2016). На практике эта процедура соблюдается далеко не всегда из-за низкой и неравномерной территориальной и хронологической плотности проанализированных образцов, предвзятости в археологической атрибуции некоторых из них, а также чрезмерном желании решить резонансную проблему при очевидном недостатке данных.
Рассмотрим, какими именно палеогенетическими данными сегодня располагает археология Кавказа и как они меняют представления о его культурном развитии в эпоху энеолита – бронзы.
Генотип кавказских охотников-собирателей
Наиболее значительным открытием в популяционной генетике Кавказа последних лет стало выделение особого генотипа «кавказских охотников-собирателей» ( Jones, Gonzalez-Fortes, Connell et al. , 2015). В мало что говорящем археологам названии были предельно обобщенно зафиксированы его географические и хронологические характеристики. Выбор довольно архаичного названия был продиктован традицией деления древних популяций на охотников-собирателей и фермеров, сложившейся в палеогенетических исследованиях процессов нео-литизации.
Впервые генотип кавказских охотников-собирателей (КОС) был определен у молодого мужчины (30–35 лет) из погребения в мезолитическом слое А2 в гроте Котиасклде и индивида из верхнепалеолитического слоя участка В в пещере Сацурблия в Западной Грузии (рис. 1; табл. 1). Абсолютный археологический возраст человеческих останков был установлен в результате их прямого радиоуглеродного датирования: Котиасклде – 9,529–9,895 калибр. лет до наших дней (95,4 %), в Сацурблия – 13,132–13,380 калибр. лет до наших дней (95,4 %) (Ibid.). Таким образом, результаты анализов древней ДНК демонстрируют генетическую преемственность у населения Западной Грузии на протяжении почти 5000 лет – в привычном археологическом исчислении с 12 до 8 тыс. до н. э., а в климатическом и геологическом – приблизительно с конца позднего дриа-са и начала голоцена. Более того, оказалось, что генетическая преемственность на Кавказе не прерывалась как минимум на протяжении последних 13 000 лет, что подтверждается широким распространением у современных кавказских народов митохондриальных и Y-хромосомных гаплогрупп, выявленных у кавказских охотников-собирателей мезолита и верхнего палеолита (Котиас: H13c, J2a; Сацурблия: K3, J) (Ibid.).
Таблица 1. Археологические памятники XII–II тыс. до н. э. на Кавказе и сопредельных территориях – источники антропологических образцов для генетического анализа
|
№ |
Памятник |
Кол-во обр. |
Культура или эпоха |
Дополнительная информация |
|
1 |
Шарахалсун 6 |
5 |
майкопская, «степной майкоп», ямная, катакомбная |
Wang, Reinhold, Kalmykov et al. , 2019 |
|
2 |
Айгурский 2 |
2 |
«степной майкоп» |
« » |
|
3 |
Ипатово 3 |
1 |
энеолит –«степной майкоп» (?) |
« » |
|
4 |
Золотаревка 2 |
1 |
ямная |
« » |
|
5 |
Расшеватский 1 |
3 |
ямная, северокавказская |
« » |
|
6 |
Расшеватский 4 |
2 |
катакомбная, посткатакомбная |
« » |
|
7 |
Невинномысский 3 |
1 |
посткатакомбная |
« » |
|
8 |
Синюха |
3 |
майкопская |
« » |
|
9 |
Марченкова Гора |
1 |
дольменная |
« » |
|
10 |
Клады |
4 |
майкопская – новосвободненская |
« » |
|
11 |
Унакозовская пещера |
3 |
дарквети-мешоковская |
« » |
|
12 |
Лысогорская 6 |
1 |
северокавказская |
« » |
|
13 |
Горячеводский 2 |
2 |
северокавказская |
« » |
|
14 |
Белый Угол 2 |
1 |
северокавказская |
« » |
|
15 |
Вонючка 1 |
1 |
энеолит |
« » |
|
16 |
Марьинская 3 |
1 |
катакомбная |
« » |
|
17 |
Марьинская 5 |
5 |
майкопская, северокавказская |
« » |
|
18 |
Кабардинка |
2 |
поздняя бронза |
« » |
|
19 |
Прогресс 2 |
3 |
энеолит, северокавказская |
« » |
|
20 |
Баксаненок |
1 |
майкопская ? |
« » |
|
21 |
Кудахурт |
2 |
средняя бронза |
« » |
|
22 |
Ногир 3 |
1 |
майкопская |
« » |
|
23 |
Великентский могильник II |
2 |
куроаракская, бедени (?) |
« » |
|
24 |
Ментеш тепе |
1 |
шулавери-шомутепинская |
Skourtanioti, Erdal, Frangipane et al. , 2020 |
|
25 |
Капс |
2 |
куроаракская |
Wang, Reinhold, Kalmykov et al. , 2019 |
|
26 |
Калаван 1 |
2 |
куроаракская |
Lazaridis, Nadel, Rollefson et al., 2016 |
|
27 |
Катнахпюр |
2 |
поздняя бронза |
« » |
|
28 |
Арени 1 |
5 |
халколит |
« » |
|
29 |
Полутепе |
1 |
шулавери-шомутепинская |
Skourtanioti, Erdal, Frangipane et al. , 2020 |
|
30 |
Хоту |
2 |
мезолит |
Lazaridis, Nadel, Rollefson et al., 2016 |
|
31 |
Зех-Габи |
6 |
халколит |
« » |
|
32 |
Ганджи-Даре |
5 |
неолит |
« » |
|
33 |
Сацурблия |
1 |
верхний палеолит |
Jones, Gonzalez-Fortes, Connell et al. , 2015 |
|
34 |
Котиасклде |
1 |
мезолит |
« » |
|
35 |
Талин |
1 |
куроаракская |
Lazaridis, Nadel, Rollefson et al., 2016 |
|
36 |
Алхантепе |
1 |
лейлатепинская |
Skourtanioti, Erdal, Frangipane et al. , 2020 |
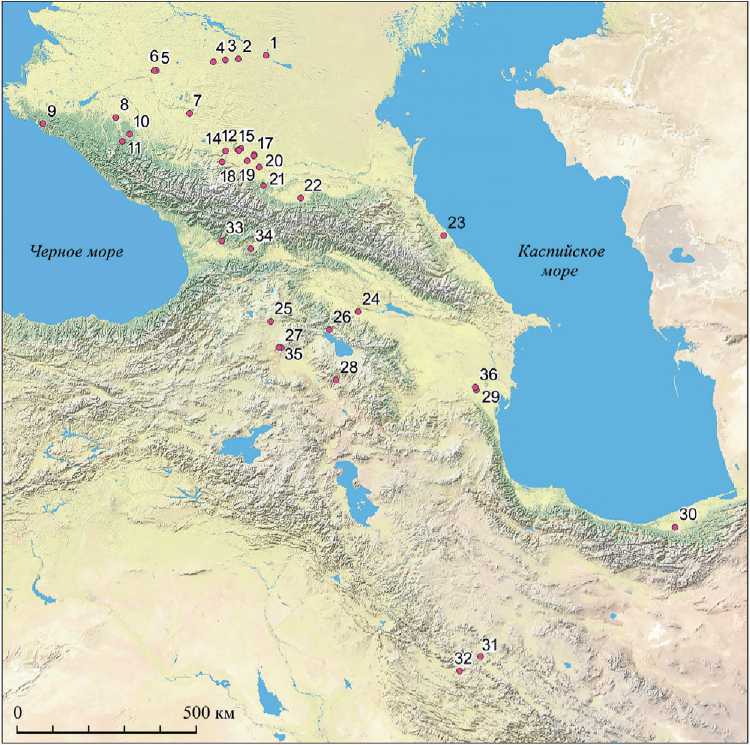
Рис. 1. Карта образцов древней ДНК из археологических памятников XII–II тыс. до н. э. на Кавказе и сопредельных территориях
(номера соответствуют порядковым номерам памятников в табл. 1)
Существенным дополнением к первоначальным характеристикам кавказского генотипа стали результаты анализов древней ДНК населения с территории современной Армении, Азербайджана и Северо-Западного Ирана (рис. 1) (La-zaridis, Nadel, Rollefson et al., 2016; Skourtanioti, Erdal, Frangipane et al., 2020). 25 образцов, в разной комбинации включающие генотип кавказских охотников-собирателей, представляют основные археологические эпохи от мезолита до поздней бронзы и охватывают период между X и II тыс. до н. э. Самые ранние и генетически наиболее близкие образцам из Западной Грузии происходят из мезолитических слоев пещеры Хоту в прикаспийском Северном Иране (9119–8637 калибр. лет до н. э.). Не нарушая генетической преемственности, хронологически за ними следуют образцы из ранненеолитических слоев поселения Ганджи-Дере в иранском Курдистане (прибл. 8200–7600 гг. до н. э.), затем из поздненеолитического слоя холма С (5837–5659 калибр. лет до н. э.) и халколи-тических слоев холмов А, B (4680–3662 калибр. лет до н. э.) поселения Зех-Габи.
На Южном Кавказе (Азербайджан) в VI тыс. до н. э. наибольшее сходство с генотипом кавказских охотников-собирателей и представителей иранского неолита (Ганджи-Дере) демонстрирует генетический профиль двух индивидов из погребений на поселениях шулавери-шомутепинской культуры Ментеш тепе и Полутепе (5717–5670; 5508–5376 калибр. лет до н. э.). При этом отмечается уменьшение градиента кавказского и иранского неолитического генетического наследия в эпоху позднего неолита – раннего халколита через всю Анатолию с востока на запад ( Skourtanioti, Erdal, Frangipane et al. , 2020).
Поздний халколит в Азербайджане представлен всего одним образцом из погребения на поселении лейлатепинской культуры Алхантепе (3776–3661 калибр. лет до н. э.), и в его генетическом профиле доминирует такое же кавказское и иранское неолитическое наследие, как и у большинства представителей восточноанатолийского позднего халколита и ранней бронзы (Арслантепе, Титриш Уйюк).
Халколитическая серия из 5 образцов с генотипом КОС представлена погребениями из пещеры Арени 1 в Армении. Черепа трех человек из вторичных погребений в сосудах (горизонт 3, ранний поздний халколит) датируются периодами 4250–4050, 4330–4060, 4230–4000 калибр. лет до н. э. Еще два образца из горизонта 2 (средний поздний халколит) датированы по археологическому контексту 4300–3700 гг. до н. э.
В середине и втор. пол. IV тыс. до н. э. кавказский генотип в сочетании с генетическим наследием представителей анатолийского и иранского халколита представлен у населения куроаракской культуры Армении.
Группа из пяти образцов с близким генетическим профилем происходит из погребения могильника Талин (погр. 115) (3347–3090 калибр. лет до н. э.), двух погребений грунтового могильника Калаван 1, одно из которых было датировано 2619–2410 калибр. лет до н. э. ( Lazaridis, Nadel, Rollefson et al. , 2016), и двух образцов из двойного погребения могильника Капс (3501–3128, 3631– 3369; 3338–3030, 3341–3030, 3366–3106 калибр. лет до н. э.) ( Wang, Reinhold, Kalmykov et al. , 2019).
Самый поздний образец генотипа похожего профиля зафиксирован у погребенного из кургана 1 могильника Катнахпюр в Армении (1501–1402 калибр. лет до н. э.).
На Северном Кавказе самый ранний образец комбинированного генотипа кавказских охотников-собирателей и представителей анатолийского халколита происходит из энеолитического погребения трех близких родственников в Унако-зовской пещере (Адыгея) (4680–4486/4599–4456/4536–4371 калибр. лет до н. э.) (Ibid.).
В IV тыс. до н. э. генотип кавказских охотников-собирателей в комбинации с генетическим наследием населения эпохи анатолийского и иранского халколита представлен 12 образцами из погребений майкопской культуры, причем без существенных различий относительно периода или локального варианта этой культуры.
В самом начале III тыс. до н. э. аналогичный по комбинации генотип определен у двух человек из коллективного погребения в катакомбе 1 Великентского могильника (Харман-тепе) в прикаспийском Дагестане. Погребение относится к великентской культуре куроаракского круга ( Kohl, Magomedov , 2014).
Во II тыс. до н. э. образцы с похожим генетическим профилем происходят из дольмена (Марченкова Гора, д. 13) и двух погребений могильника Кудахурт в горной части, соответственно, Северо-Западного и Центрального Кавказа.
Генетически образцы, представляющие энеолит, майкопскую, куроаракскую и дольменную культуры на Северном и Южном Кавказе, неразличимы друг от друга ( Wang, Reinhold, Kalmykov et al. , 2019) и не выходят за рамки генотипа, характерного для армянского и иранского халколита ( Lazaridis, Nadel, Rollefson et al. , 2016).
Для оценки субрегиональных различий на Северном Кавказе пока нет генетических образцов, синхронных образцам с Южного Кавказа VI тыс. до н. э. – шулавери-шомутепинской культуры. В V и IV тыс. до н. э. эти различия связаны главным образом с изменением соотношения главных генетических компонентов – кавказского, иранского и анатолийского.
Учитывая значительные территориальные и хронологические лакуны в существующей палеогенетической базе данных, новые анализы неолитических и халколитических популяций с Южного Кавказа, из Восточной Анатолии и Северной Месопотамии могут это заключение уточнить ( Skourtanioti, Erdal, Fran-gipane et al. , 2020).
Кавказский генотип охотников-собирателей за пределами Кавказа
Полагают, что генотип кавказских охотников-собирателей сформировался с отделением этой популяции от так называемых западных охотников-собирателей приблизительно 45 тыс. л. н. и от предков неолитических фермеров – примерно 25 тыс. л. н., где-то в период максимума последнего оледенения (LGM) ( Jones, Gonzalez-Fortes, Connell et al. , 2015).
Вероятно, в период первого значительного потепления (Аллередский интерстадиал) приблизительно 14 тыс. л. н. в популяциях Юго-Западной Европы появляются генетические признаки ближневосточного происхождения, в том числе близкие к кавказскому генотипу ( Fu, Posth, Hajdinjak et al. , 2016).
Раннее влияние кавказского генотипа на западноевропейский генетический пул фиксируется с конца эпохи раннего неолита, причем оно оказалось сильнее на севере, а не на юге Европы ( Jones, Gonzalez-Fortes, Connell et al. , 2015).
В Западной Азии ближайшими генетическими родственниками кавказским охотникам-собирателям приходятся ранние фермеры из иранского Загроса (приблизительно 10 тыс. л. н.), но при этом их родство носит опосредованный характер ( Broushaki, Thomas, Link et al. , 2016).
Для археологии северных от Кавказа территорий особый интерес представляет кавказский вклад в генетический профиль степных популяций эпохи энеолита – бронзы.
Установлено, что генотип кавказских охотников-собирателей в сочетании с генотипом восточных охотников-собирателей – потомков древних евразийцев представлен у энеолитического населения на Нижней Волге и в подгорной зоне предкавказской степи уже в V, а возможно, даже с конца VI тыс. до н. э. (Хва-лынский II мог., погр. 1, 12, 17: хронологический диапазон 5200–4000 гг. до н. э. (95,4 %); Прогресс 2, 1/37: 4336–4173 калибр. лет до н. э., 4/9: 4336–4173 калибр. лет до н. э.; Вонючка 1, 1/8: 4229–4065 калибр. лет до н. э.) ( Mathieson, Lazaridis, Rohland et al. , 2015; Wang, Reinhold, Kalmykov et al. , 2019).
В середине – втор. пол. IV тыс. до н. э. кавказский генотип почти в аналогичной «степной» комбинации представлен у обитателей предкавказской степи. Эта небольшая группа из 7 погребений под довольно провокативным названием «степной майкоп», заимствованным из полевого жаргона. Археологические основания для включения в группу трех из семи погребений довольно спорны по следующим причинам.
Погребение 18 в кургане 2 (Шарахалсун 6) относится к катакомбной культуре и, по данным стратиграфии, не может относиться ни к энеолитическим, ни к майкопским ( Яковлев , 2004), а его датировка, вероятнее всего, результат ошибочной маркировки образца ( Izbitser , 2018).
Курганы в районе станицы Марьинской расположены в пределах подгорной зоны Ставрополья – метрополии майкопской культуры, поэтому нет оснований относить одни погребения из этих курганов к майкопской культуре, а другие к «степному майкопу» (Марьинская 5, 1/32).
Безынвентарное погребение 187 из кургана 2 (Ипатово 3) с неустановленной позой погребенного первоначально было атрибутировано как энеолитическое ( Кореневский и др. , 2007) и к «степному майкопу» отнесено по результатам радиоуглеродного датирования ( Wang, Reinhold, Kalmykov et al. , 2019).
Вероятно, эти особенности археологической группировки отразились и на результатах общей генетической характеристики группы, тем более что погребенный из Большого Ипатовского кургана (п. 187) – один из двух представителей с генетическими отклонениями от стандарта «степного майкопа». Генотип этого и еще одного индивида из погребения 11 в кургане 2 (Шарахалсун 6) включает генетический компонент, характерный для представителей анатолийского и иранского неолита. Наиболее вероятно, что источником этой генетической примеси к «степному майкопу» были современные им популяции к югу от Кавказа. С учетом скорости генетических мутаций, формирование такой разновидности генотипа могло произойти примерно за 560 лет или в течение 20 поколений (Ibid.).
Сочетание кавказского генотипа охотников-собирателей с комбинированным генетическим наследием анатолийских и иранских фермеров, впервые зафиксированное у «степных майкопцев», представлено и в генетическом профиле более позднего степного населения Предкавказья – ямной, северокавказской и катакомбной культур. Однако по своему происхождению анатолийско-иранское наследие у представителей этих культур, возможно, не южного, а западного происхождения, так как оно сочетается с заметным присутствием генотипа западных охотников-собирателей (Ibid.). Последний компонент отсутствует у двух индивидов из могильника Кабардинка (к. 2, п. 1б: 2196–1977 калибр. лет до н. э.; к. 2, п. 2: 2188–2026 калибр. лет до н. э.), включенных в степную группу поздней северокавказской культуры, хотя могильник расположен в горной части Северного Кавказа.
Еще один индивид, чей генотип не укладывается в степные стандарты, происходит из погребения в могильнике Невинномысский 3 (к. 6, п. 5: 2116– 1925 калибр. лет до н. э.). Его генетический профиль ближе всего к профилю представителей «степного майкопа». В археологической группировке это погребение отнесено к посткатакомбной лолинской культуре (Ibid.).
В итоге археология Кавказа и предкавказской степи располагает сейчас данными о генотипах 65 индивидов, ассоциированных с 14 культурными группами эпохи энеолита – бронзы (табл. 1). Среди них нет ни одной «чистой» популяции. Генетический профиль всех групп представляет комбинацию из нескольких – от 2 до 4 – главных компонентов, различных по возрасту и происхождению. Причем количественное соотношение этих элементов не является постоянной величиной и зависит от количества учтенных гипотетически родственных популяций при генетическом анализе примесей.
Согласно результатам анализов, почти половина генетического наследия всех кавказских групп – горных и степных – связана с кавказским генотипом охотников-собирателей, или смешанным кавказским и иранским неолитическим генотипом. Вероятно, этот условно «кавказско-переднеазиатский» генетический компонент проник в Восточную Европу достаточно рано, так как уже в VI тыс. до н. э. зафиксирован на северо-западе России (Южный Олений остров в Ладожском озере) и в Нижнем Поволжье (Лебяжинка 4) ( Haak, Lazaridis, Patterson et al. , 2015). Его присутствие в степной зоне со временем увеличивается, достигнув максимума у населения, ассоциированного с ямной культурой.
Вторым главным компонентом в степном генетическом профиле эпохи энеолита – ранней бронзы остается генотип восточноевропейских охотников-собирателей. В таком сочетании оба генотипа представлены у населения северокавказской степи в эпоху раннего энеолита.
В это время уже существовала генетическая граница между населением открытой степи и Кавказом. У кавказского населения, в отличие от степного, второй главный генетический компонент переднеазиатского происхождения, родственного генотипу представителей анатолийского или, в комбинации, анатолийского и иранского халколита. Присутствие минимальной генетической примеси, характерной для восточных и западных охотников-собирателей, вероятно, является частью генетического вклада анатолийских фермеров, а не степного населения ( Lazaridis, Nadel, Rollefson et al. , 2016; Wang, Reinhold,Kalmykov et al. , 2019).
На Кавказе этот генетический профиль с вариациями соотношения главных компонентов (кавказского, анатолийского и иранского) разделяют представители как минимум шести культур по обеим сторонам Главного хребта: даркве-ти-мешоковская, шулавери-шому-тепинская, лейлатепинская, майкопская, доль-менная и куроаракская. Основной хронологической тенденцией здесь является региональная генетическая гомогенизация населения с неолита до эпохи бронзы ( Skourtanioti, Erdal, Frangipane et al. , 2020).
У населения предкавказской степи траектория генетических изменений в IV и III тыс. до н. э. выглядит иначе и не укладывается в простую двухкомпонентную модель, сочетающую генетическое наследие раннего степного энеолита и анатолийского или иранского халколита. Исключение составляют только два представителя «степного майкопа», чей генетический профиль является комбинацией энеолитического степного и анатолийского/иранского неолитического наследия.
Для более поздних степных кавказских групп, включая ямную, северокавказскую и катакомбную, характерно постепенное увеличение анатолийского генетического компонента в сочетании с генетическим наследием западных охотников-собирателей. Эта комбинация указывает на западное, а не южное происхождение анатолийского вклада в степной генетический пул. Источник этого генетического влияния окончательно не определен, но на его роль подходят как энеолитические группы из северо-причерноморской степи, так и культура шаровидных амфор, которые включают все четыре компонента – кавказский, западный и восточный типы охотников-собирателей плюс анатолийский неолитический ( Wang, Reinhold, Kalmykov et al. , 2019).
Есть еще два палеогенетических наблюдения, важных для анализа археологического контекста.
Одно из них касается расчетов даты смешения генетических компонентов при формировании комбинаций, характерных для различных территориально-хронологических популяций. Для кавказской («горной») популяции эпохи ранней бронзы эта дата отстоит от конца IV тыс. до н. э. на 121 ± 35 поколение, что означает, что она сформировалась (1 поколение – 28 лет) примерно в середине VII тыс. до н. э., – одновременно с родственной позднехалколитической анатолийской популяцией (Арслантепе и др.). На это же время приходится формирование поздненеолитической шулавери-шомутепинской популяции, но от этой даты группу отделяет всего 34 ± 15 поколения ( Skourtanioti, Erdal, Frangipane et al. , 2020).
Второе важное наблюдение связано с определением направления основных генетических потоков.
Нет никаких признаков инфильтрации генотипов степного пула эпохи энеолита – бронзы ни на южный Кавказ, ни на сопредельные территории Cеверо-Западного Ирана и Восточной Анатолии (Ibid.).
Обсуждение
Культурно-историческая интерпретация результатов палеогенетических анализов
Тематика культурно-исторической интерпретации результатов палеогенети-ческих анализов пока ограничена, главным образом, реконструкцией миграций древнего населения, и этому есть несколько объяснений.
Первые полногеномные исследования образцов древней ДНК эпохи энеолита – бронзы с просторов евразийских степей и Кавказа проходили в условиях острого соперничества между несколькими западноевропейскими и американскими палеогенетическими лабораториями, осложненного труднодоступностью подходящих антропологических коллекций на территории бывшего СССР. Финансирование проектов осуществляли западные научные фонды, соответственно, главными идеологами проектов были западноевропейские и американские генетики и археологии. Участие в проектах археологов и антропологов из России ограничивалось в основном консультациями, сбором образцов и описанием их археологического контекста.
Парадоксальным образом высокую научную мотивацию генетикам обеспечивали исключительно амбициозные цели археологов, намеревавшихся решить старую проблему географической локализации прародины индоевропейских языков новыми средствами.
Две статьи с первыми результатами анализов ДНК населения евразийской степи энеолита – бронзы соперничающие группы генетиков и археологов опубликовали демонстративно синхронно в 2015 г. ( Haak, Lazaridis, Patterson et al. , 2015; Allentoft, Sikora, Sjögren et al. , 2015). Чуть позднее в том же году вышли еще две статьи ( Jones, Gonzalez-Fortes, Connell et al. , 2015; Mathieson, Lazaridis, Rohland et al. , 2015), из которых стало ясно, что всем четырем командам генетиков остро не хватало образцов, которыми располагали конкуренты. Несмотря на это, во всех четырех публикациях с разной долей уверенности евразийские потоки генов интерпретировались как индикаторы массовых миграций и распространения индоевропейских языков. Это вызвало огромный, в целом позитивный научный и общественный резонанс, на фоне которого сомнения некоторых археологов расценивались как проявление корпоративного консерватизма ( Klejn, Haak, Lazaridis et al. , 2018).
Надо признать, что археологическим лидерам проектов – К. Кристиансену и Д. Энтони – удалось очаровать своих молодых коллег-генетиков теорией степной индоевропейской прародины до такой степени, что они отступили от процедуры независимой от археологической группировки результатов генетических анализов. Эта уступка археологам в определенной степени компенсируется недвусмысленными комментариями палеогенетиков к различным фрагментам теории степной прародины. По их мнению, не находят генетического подтверждения ни миграция из степи через Балканы в Анатолию ( Mathienson, Alpaslan-Roodenberg, Posth et al. , 2018), ни степной маршрут распространения иранских языков ( Broushaki, Thomas, Link et al. , 2016). Другими словами, теория степной прародины во всех своих миграционных составляющих генетически пока не подтверждается.
Открытие в степном генетическом профиле эпохи энеолита – бронзы существенного кавказского компонента стало причиной оригинальной попытки согласовать теории степной и анатолийской индоевропейской прародины. В этом предположении Кавказу отводится роль канала, по которому народы с кавказским и переднеазиатским генетическим наследием, говорящие на протоиндоевропейских языках, перетекали с юга на север и дальше на восток и запад. Почему сам Кавказ при этом оставался преимущественно неиндоевропейским, гипотеза не поясняет ( Wang, Reinhold, Kalmykov et al. , 2019).
Если избегать наивной и буквальной интерпретации результатов генетических анализов, о чем предупреждают палеогенетики (Skourtanioti, Erdal, Frangipane et al., 2020), то часть этих данных может оказаться важным аргументом в культурно-историческом объяснении археологических явлений. Например, открытие генетического единства населения восточноевропейской степи и западноевропейской культуры шнуровой керамики в середине III тыс. до н. э. (Haak, Lazaridis, Patterson et al., 2015; Allentoft, Sikora, Sjögren et al., 2015) резоннее рассматривать в контексте не лингвистической индоевропейской головоломки, а почти забытой сейчас дискуссии о родстве «шнуровых» культур Западной и Восточной Европы (Hausler, 1963; 1976; Мерперт, 1976; 1977)
Археологам еще предстоит научиться пользоваться результатами палеоге-нетических анализов, не вырывая из контекста отдельные характеристики популяционного генетического профиля – гаплотипы и гаплогруппы Y-хромосомы и мтДНК. Установлено, что гаплотипы мтДНК как самостоятельный признак мало пригодна для обнаружения признаков массовых миграций ( Emery et al. , 2015), а пространственная локализация гаплогрупп мтДНК (например, на Кавказе) не имеет такой же выраженной территориальности, как гаплогруппы Y-хромосомы ( Wang, Reinhold, Kalmykov et al. , 2019).
По этой причине реконструкции сверхдальних миграций из Западной Европы на Кавказ или в прямо противоположном направлении на основе сходства отдельных гаплогрупп мтДНК и нДНК в равной степени сомнительны ( Nedo-luzhko et al. , 2014; Wang, Reinhold, Kalmykov et al. , 2019). Поучительно, что в обоих случаях результаты генетических анализов используются как весомые аргументы в пользу двух взаимоисключающих объяснений сходства изображений на стенах мегалитических гробниц на Кавказе (Новосвободная) и в Европе (Гехлитцш) ( Резепкин , 2012; Hansen , 2018). Здесь уместно отметить, что даже доминирование в Западной Европе гаплогруппы R1b может быть связано не только с мигрантами бронзового века из восточноевропейской степи ( Haak, Lazaridis, Patterson et al. , 2015). Присутствие этой гаплогруппы на самом западе Европы за несколько тысяч лет до «степняков» указывает на ее более раннюю европейскую историю ( Fu, Posth, Hajdinjak et al. , 2016).
Если оценивать роль палеогенетических анализов в решении конкретных проблем археологии Кавказа эпохи энеолита – бронзы, оставив в стороне загадку индоевропейской прародины, то в первую очередь следует отметить почти полное отсутствие майкопского вклада в генетический профиль населения сопредельной степи. В контексте полученных результатов «степной майкоп» выглядит позднеэнеолитической группой степных аборигенов, чье близкое соседство с высокоразвитой культурой обернулось для них односторонними выгодами и возможностью обогатить свою материальную культуру некоторыми майкопскими символами, например, небольшими орнаментированными сосудами невиданного в степи качества. Эта ситуация может стать поводом к пересмотру модели взаимодействия технически отсталого энеолитического севера с развитым раннебронзовым югом и означать, что майкопская культура никогда не стремилась колонизировать предкавказскую степь, в которой не было ни привлекательных для нее ресурсов, ни зажиточных партнеров, ни опасных врагов. Для степи она оставалась относительно пассивным донором технических инноваций, и ее самый северный анклав на Нижнем Дону (константиновская культура) был ассимилирован и угас еще до заката метрополии в предгорьях Северного Кавказа.
Нет майкопского генетического наследия и у населения ямной культуры в предкавказской степи. Одновременно отсутствуют и смешанные комплексы с аутентичными майкопскими и ямными вещами. Вместе с данными стратиграфии все это складывается в картину смены культур, при которой едва ли была возможна широкомасштабная передача технологий майкопской культуры ям-ной. Их большая часть исчезла вместе с майкопской культурой, а у ямной, возможно, были и альтернативные источники для заимствований на юге.
Хронология и география распространения северокавказской культуры оставляет несколько вариантов для культурно-исторической интерпретации ее генетического профиля. В степном Предкавказье и подгорной зоне эта культура приходит на смену ямной и майкопской, поэтому с равной вероятностью генетическое наследие этих культур могло войти в генотип северокавказцев.
Отдельного комментария заслуживает наблюдение о совпадении генетической и географической границ между Кавказом и степью в эпоху энеолита – бронзы ( Wang, Reinhold, Kalmykov et al. , 2019). При детальном рассмотрении картина на самом деле более сложная – открытую степь от подножия северных склонов Кавказа отделяет довольно широкая подгорная зона и часть Ставропольской возвышенности, которые в разные исторические и климатические эпохи были заселены народами кавказского или степного происхождения. Другими словами, граница была довольно подвижной, и Кавказ представлял не столько мост между Европой и Азией, сколько асимметричный полупроницаемый барьер на миграционном пути между континентами ( Yunusbayev, Metspalu, Järve et al. , 2012).
Самым неожиданным результатом анализов древней ДНК народов Кавказа эпохи палеометалла оказался вывод о высокой степени однородности всего кавказского генетического пула независимо от культурной и хронологической ассоциации образцов ( Wang, Reinhold, Kalmykov et al. , 2019). Оказалось, что культурное разнообразие Кавказа превосходит генетическое и там, где археологи проводят четкую границу (например, между майкопской и куроаракской культурами), палеогенетики различий не видят. Причем этот феномен проявляет себя как в синхронном, так и асинхронном срезах. При смене майкопской культуры дольменной, или ямной – катакомбной, культурные изменения по археологическим меркам выглядят радикальными, а генетические, как оказалось, имеют вид вариаций или вообще отсутствуют. Эти различия генетики и археологи уловили одновременно, предлагая несколько вариантов согласования данных в рамках или новой номенклатуры археогенетических кластеров ( Eisenmann, Bánffy, van Dommelen et al. , 2018), или ревизии основ традиционной археологической таксономии и критериев различных уровней культурной дифференциации (культуры, технокомплексы, индустрии, традиции и т. д.) ( Riede et al. , 2019).
Косвенным образом эти расхождения и попытки их устранения отражают разную природу связанных между собой культурных и генетических процессов (модель генетической и культурной коэволюции или теория двойного наследования) ( Cavalli-Sforza, Feldman , 1981; Boyd, Richerson , 1985). Вопрос в том, как именно генетический и археологический пространственно-временные континуумы пересекаются и взаимодействуют друг с другом.
Археология и палеогенетика: цели и средства
Надо признать, что, пусть и не с первой попытки, палеогенетикам удалось продемонстрировать мощь нового инструмента изучения человеческого прошлого, который навсегда войдет в арсенал археологов наряду с радиуглеродным методом датирования, изотопными анализами, ГИС и ЛИДАР ( Sedig , 2019). Оценивая достоинства метода, в первую очередь отмечают почти недоступную археологам возможность проследить перемещение людей прямым, а не косвенным («по горшкам») образом. Поэтому в фокусе исследований чаще всего оказывается генетическое тестирование миграционных концепций разного масштаба и тематики.
Второе достоинство – в способности определять степень кровного родства, третье – выделять и датировать определенные генетические мутации, связанные с изменениями в образе жизни людей. В комбинации с другими методами это открывает археологам почти безграничные возможности в изучении разнообразия социальных и экономических форм жизнеобеспечения.
Однако нельзя сказать, что сотрудничество между археологами и генетиками складывается гладко. С одной стороны, развитие техники секвенирования и получение массовых данных высокого разрешения вводят генетиков в искушение пренебречь археологическим контекстом при культурно-исторической интерпретации генетических данных. С другой, у археологов нет ясных представлений о реальных возможностях палеогенетики ( Lebrasseur et al. , 2018).
Палеогенетики вовсе не собираются переходить к обслуживанию археологов, как бы этого последним ни хотелось. Палеогенетические программы исследований только отчасти совпадают с археологическими, главным образом на этапе территориально-хронологической селекции древних антропологических образцов. Для палеогенетиков археология остается одним из инструментов в изучении популяционной истории человечества, примерно так же, как для археологов палеогенетика – один из методов изучения культурных сообществ. При таком различии целей, естественно, трудно ожидать равноправной научной кооперации. Для нее нужна настоящая общность интересов, а не потребительский расчет.
Значительное превосходство культурного разнообразия над генетическим, как это оказалось на Кавказе в эпоху бронзы, вызывает прямой вопрос: что происходит с людьми – носителями культуры, которая исчезает из археологического пространства без признаков мора, истребления или эмиграции? Остается предположить, что, утратив свою культурную идентичность, они растворяются среди населения других культур, добавляя ему свою долю генетического разнообразия. В таком случае именно процесс смены культур и связанной с ним генетической рекомбинации представляет область взаимного научного интереса археологов и генетиков. Это та часть фундаментальной проблемы связей между культурным и генетическим развитием человеческих сообществ, решение которой генетики и археологи могут найти только сообща.
Культура и ДНК – археология и палеогенетика: новые ориентиры исследований
Если о культуре как явлении судить по ее основной функции – формированию групповой идентичности, то в системе основных общественных структур – экономической, социальной и культурной – последняя окажется ведущей, так как она создает базовые условия для развития двух остальных ( Pagel , 2012). Вероятно, что по своей природе культура является продуктом генетической эволюции человека и, как метафорично высказался эволюционный биолог Э. Уилсон, «гены держат культуру на поводке», добавив, правда, что «на очень длинном» ( Wilson , 1978).
Культура – это эволюционное преимущество человека перед другими видами в поддержании генетического разнообразия. Постоянное воспроизводство многообразия культур является необходимым условием и частью внебиологиче-ского механизма поддержания благоприятных условий для сохранения генетического разнообразия вида рода Homo.
Границы между культурами служат барьерами на пути несущих угрозу генетических потоков, связанных с миграцией, агрессией или эпидемией.
Нестабильность внешних условий существования представляет серьезный вызов традиционным культурам с низкой хозяйственно-экономической адаптивностью. За пределами возможностей хозяйственных перемен неизбежно наступает период экономического упадка, демографического стресса и, как следствие, кризиса культурной идентичности, так как верность прежним ценностям больше не гарантирует процветания.
Как только лимиты изменчивости оказываются исчерпанными – жизненный цикл культуры завершается, она угасает, уступая место более приспособленной к новым природным и культурно-историческим условиям.
Эмпирический опыт археологии свидетельствует, что все культуры «смертны», но доживают при этом до разного возраста. От чего именно может зависеть продолжительность их жизненного цикла, демонстрируют, в частности, результаты прикладных генетических исследований, например, двух современных популяций шапсугов, проживающих по разные стороны главного хребта на Западном Кавказе ( Почешхова , 2008). Обе группы испытали критическое сокращение численности и пострадали в конце XIX в. от эффекта «бутылочного горлышка» – опасного сокращения генофонда и уменьшения его разнообразия. В одной из групп тенденция к сокращению популяции со временем не изменилась и к ней добавились сопутствующие генетические проблемы в виде роста числа наследственных заболеваний, а в другой – наметился явный тренд к восстановлению, главным образом за счет поддержки ключевых элементов традиционной культуры. Конечно, полного восстановления генетического разнообразия этой популяции шапсугов уже никогда не произойдет, но она, скорее всего, переживет родственную на противоположном склоне Кавказского хребта.
Перспективы изучения взаимосвязей между состоянием генофонда популяций и этапами «расцветов и закатов» ассоциированных с ними культур могут оказаться в равной степени продуктивными для палеогенетиков и археологов.
Актуальность подобных исследований будет только возрастать из-за легкости их перевода из исторической плоскости в современную, возможностей прикладных исследований и прогностической ценности результатов.
Заключение
Археология Кавказа и предкавказской степи располагает сейчас данными о генотипах 65 индивидов из 33 памятников, ассоциированных с 14 культурными группами эпохи неолита – бронзы V–II тыс. до н. э. Эти результаты трудно переоценить, так как они закладывают основы нового археогенетического направления в кавказской доисторической археологии. Независимо от того, в какой степени эти данные будут дополнены и даже пересмотрены в будущем, они навсегда останутся отправной точкой в грядущих исследованиях.
Можно выделить семь основных черт общей палеогенетической картины региона.
Главной, прежде неизвестной особенностью является раннее, не позднее XII тыс. до н. э., формирование генотипа кавказских охотников-собирателей и его распространение за пределы Кавказа.
Второй характерной чертой является существование на Кавказе непрерывной генетической преемственности с эпохи верхнего палеолита.
Третий важный элемент – раннее, не позднее VI тыс. до н. э., появление кавказского и связанного с ним переднеазиатского (иранского и анатолийского) генетического компонента в степном генетическом пуле.
Четвертая особенность кавказской археогенетической ситуации состоит в сохранение генетических различий между населением Кавказа и сопредельной открытой степи на протяжении всей эпохи энеолита – бронзы.
Пятым региональным признаком является существование вдоль северных склонов Кавказа генетической буферной зоны, которую, в зависимости от исторических и климатических условий, занимали степняки или кавказцы.
Шестая характерная черта – односторонняя проводимость потока генов через Кавказ только в направлении с юга на север.
Наконец, седьмая региональная особенность – это преобладание культурного разнообразия над генетическим.
Возможно, полученные результаты не соответствуют ожиданиям тех археологов, кто рассчитывал на немедленное генетическое решение проблем происхождения культур или как минимум локализацию их исходных областей распространения, включая прародину призрачных индоевропейцев. Генетическая и культурная сущности людей пересекаются между собой, но они не идентичны друг другу, и если было бы наоборот, то необходимость в значительной части археологических исследований просто бы отпала. У генетики нет средств для решения археологических проблем, но она может помочь найти их. Для этого прежде всего предстоит согласовать генетическое и археологическое таксономическое группирование.
В археологии палеогенетика – это больше, чем новая технология. Она привносит элементы новой идеологии, ориентированной на изучение природы не столько сходств, сколько бесконечно воспроизводимых различий, составляющих основу культурного и генетического разнообразия. Это принципиальное смещение акцентов может помочь традиционной археологии обрести новую мотивацию и в большей степени отвечать общественному запросу на актуальные знания.
Список литературы Генетическое разнообразие древних народов Кавказа и сопредельной степи в эпоху энеолита - бронзы (V-II тыс. до н. э.): основные результаты и проблемы культурно-исторической интерпретации
- Кореневский С. Н., Белинский А. Б., Калмыков А. А., 2007. Большой Ипатовский курган на Ставрополье как археологический источник по археологии бронзового века на степной границе Восточной Европы и Кавказа. М.: Наука. 232 с.
- Мерперт Н. Я., 1976. Древнеямная культурно-историческая область и вопросы формирования культур шнуровой керамики // Восточная Европа в эпоху камня и бронзы / Отв. ред. Л. В. Кольцов. М.: Наука. С. 103–127.
- Мерперт Н. Я., 1977. Из истории древнеямных племен // Проблемы археологии Евразии и Северной Америки / Отв. ред. Н. Л. Членова. М.: Наука. С. 68–80.
- Почешхова Э. А., 2008. Геногеографическое изучение народов Западного Кавказа: автореф. дис. … д-ра мед. наук. М. 48 с.
- Резепкин А. Д., 2012. Новосвободненская культура (на основе материалов могильника «Клады»). СПб.: Нестор-История. 343 с.
- Яковлев А. В., 2004. Отчет о раскопках курганного могильника Шарахалсун 6 в 2001 г. // Архив ИА РАН. Р-1. № 6695.
- Allaby R., Gutaker R., Clarke A. C., Pearson N., Ware R., Palmer S. A., Kitchen J. L., Smith O., 2015. Using archaeogenomic and computational approaches to unravel the history of local adaptation in crops // Philosophical Transactions of the Royal Society. B: Biological Sciences. 370 (1660). 20130377.
- Allentoft M. E., Sikora M., Sjögren K.-G. et al., 2015. Population genomics of Bronze Age Eurasia // Nature. Vol. 522 (7555). P. 167–172.
- Anthony D., 2007. The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton: Princeton University Press. 568 p.
- Boyd R., Richerson P., 1985. Culture and the evolutionary process. Chicago: University of Chicago Press. 340 p.
- Broushaki F., Thomas M. G., Link V. et al., 2016. Early Neolithic genomes from the eastern Fertile Crescent // Science. Vol. 353 (6298). P. 499–503.
- Cavalli-Sforza L., Feldman M., 1981. Cultural transmission and evolution: a quantitative approach. Princeton: Princeton University Press. 368 p.
- Eisenmann S., Bánffy E., van Dommelen P. et al., 2018. Reconciling material cultures in archaeology with genetic data: The nomenclature of clusters emerging from archaeogenomic analysis // Scientific Reports. Vol. 8. 13003.
- Emery L. S., Akey J. M., Bamshad M. J., Magnaye K. M., Bigham A. W., 2015. Estimates of Continental Ancestry Vary Widely among Individuals with the Same mtDNA Haplogroup // American journal of human genetics. Vol. 96. Iss. 2. P. 183–193.
- Fu Q., Posth C., Hajdinjak M. et al., 2016. The genetic history of Ice Age Europe // Nature. Vol. 534 (7606). P. 200–205.
- Gamba C., Jones E. R., Teasdale M. D. et al., 2014. Genome flux and stasis in a five millennium transect of European prehistory // Nature Communications. Vol. 5. P. 5257.
- Gimbutas M., 1970. Proto-Indo-European Culture: The Kurgan Culture during the Fifth, Fourth, and Third Millennia B.C. // Indo-European and Indo-Europeans / Eds.: G. Cardona, H. Hoenigswald, A. Senn. Philadelphia: University of Pennsylvania Press. P. 155–197.
- Haak W., Forster P., Bramanti B. et al., 2005. Ancient DNA from the first European farmers in 7500-yearold Neolithic sites // Science. Vol. 310 (5750). P. 1016–1018.
- Haak W., Lazaridis I., Patterson N. et al., 2015. Massive migration from the steppe was a source forIndo-European languages in Europe // Nature. Vol. 522 (7555). P. 207–211.
- Hadelberg E., Hofreiter M., Keyser C., 2015. Introduction. Ancient DNA: the first three decades // Philosophical Transactions of the Royal Society. B: Biological Sciences. Vol. 370. No. 1660. 20130371.
- Hansen S., 2018. Elements for an Iconography of Bronze Age Graves in Europe // Anatolian Metal VIII. Rahden: Marie Leidorf. S. 281–293. (Der Anschnitt. Beiheft; 39.) (Veröffentlichungen aus dem Deutschen Bergbau-Museum Bochum; nr. 226.)
- Hausler A., 1963. Ist eine Ableitung der Schnurkeramikvon der Ockergrabkultur möglich? // Forschungen und Fortschritte. Bd. 37 (12). S. 363–368.
- Hausler A., 1976. Die Graber der älteren Ockergrabkultur zwischen Dnepr und Karpaten. Berlin: Akademie-Verlag. 223 S.
- Izbitser E., 2018. Bulls and wagons: an «innovative» approach to representing archaeological data // Tyragetia. Serie nouă. Vol. XII. No. 1. P. 115–120.
- Jensen T. Z. T., Niemann J., Højholt Iversen K. et al., 2019. A 5700 year-old human genome and oral microbiome from chewed birch pitch // Nature Communications. Vol. 10. 5520.
- Jones E. R., Gonzalez-Fortes G., Connell S. et al., 2015. Upper Palaeolithic genomes reveal deep roots of modern Eurasians // Nature Communications. Vol. 6. 8912.
- Klejn L. Haak W., Lazaridis I. et al., 2018. Discussion: Are the Origins of Indo-European Languages Explained by the Migration of the Yamnaya Culture to the West? // European Journal of Archaeology. Vol. 21. Iss. 1. P. 3–17.
- Kohl P., Magomedov R., 2014. Early Bronze developments on the West Caspian coastal plain // Paléorient. Vol. 40. No. 2. P. 93–114.
- Krings M., Stone A., Schmitz R. W., Krainitzki H., Stoneking M., Pääbo S., 1997. Neandertal DNA sequences and the origin of modern humans // Cell. Vol. 90. Iss. 1. P. 19–30.
- Lazaridis I., Nadel D., Rollefson G. et al., 2016. Genomic insights into the origin of farming in the ancient Near East // Nature. Vol. 536 (7617). P. 419–424.
- Lebrasseur O., Ryan H., Abbona C., 2018. Bridging Archaeology and Genetics // Environmental Archaeology. Interdisciplinary Contributions to Archaeology / Eds.: E. Pişkin, A. Marciniak, M. Bartkowiak. Cham: Springer. P. 111–132.
- Mallory J. P., 1989. In Search of the Indo-Europeans: Language, Archaeology, and Myth. London: Thames & Hudson. 288 p.
- Mathieson I., Lazaridis I., Rohland N. et al., 2015. Genome-wide patterns of selection in 230 ancient Eurasians // Nature. Vol. 528 (7583). P. 499–503.
- Mathieson I., Alpaslan-Roodenberg S., Posth C. et al., 2018. The genomic history of southeastern Europe // Nature. Vol. 555 (7695). P. 197–203.
- Nedoluzhko A., Boulygina E. S., Sokolov A. S., Tsygankova S. V., Gruzdeva N. M., Rezepkin A. D., Prokhortchouk E. B., 2014. Analysis of the Mitochondrial Genome of a Novosvobodnaya Culture Representative using Next-Generation Sequencing and Its Relation to the FunnelBeaker Culture // Acta Naturae. Vol. 6. № 2 (21). P. 31–35.
- Orlando L., Ginolhac A., Zhang G. et al., 2013. Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse // Nature. Vol. 499 (7456). P. 74–78.
- Pagel M., 2012. Wired for Culture: Origins of the Human Social Mind. The Natural History of Human Cooperation. New York: W. W. Norton. 432 p.
- Pinhasi R., Fernandes D., Sirak K., Cheronet O., 2019. Isolating the human cochlea to 620 generate bone powder for ancient DNA analysis // Nature Protocols. 14. P. 1194–1205.
- Renfrew C., 1990. Archaeology and Language: The Puzzle of Indo-European Origins. Cambridge: Cambridge University Press. 346 p.
- Renfrew C., 2000. Archaeogenetics: Towards a population prehistory of Europe // Archaeogenetics: DNA and the population prehistory of Europe / Eds.: C. Renfrew, K. Boyle. Cambridge, UK: McDonald Institute for Archaeological Research. P. 3–12.
- Riede F., Hoggard C., Shennan S., 2019. Reconciling material cultures in archaeology with genetic data requires robust cultural evolutionary taxonomies // Palgrave Communications. Vol. 5. 55.
- Rohland N., Harney E., Mallick S., Nordenfelt S., Reich D., 2015. Partial uracil–DNA–glycosylase treatment for screening of ancient DNA // Philosophical Transactions of the Royal Society. B: Biological Sciences. 370 (1660). 20130624.
- Scheu А. et al., 2015. The genetic prehistory of domesticated cattle from their origin to the spread across Europe // BMC Genetics. Vol. 16. 54.
- Sedig J., 2019. Ancient DNAs Impact on Archaeology: What Has Been Learned and How To Build Strong Relationships // The SAA Archaeological Record. Vol. 19. P. 26–32.
- Sirak K., Fernandes D., Cheronet O. et al., 2020. Human auditory ossicles as an alternative optimal source of ancient DNA // Genome Research. 30 (3). P. 427–436.
- Skourtanioti E., Erdal Y. S., Frangipane M. et al., 2020. Genomic History of Neolithic to Bronze Age Anatolia, Northern Levant, and Southern Caucasus // Cell. Vol. 181. Iss. 5. P. 1158–1175.
- Voelkerding K. V., Dames S. A., Durtschi J. D., 2009. Next-generation Sequencing: From Basic Research to Diagnostics // Clinical Chemistry. Vol. 55. No. 4. P. 651–658.
- Wang C., Reinhold S., Kalmykov A. et al., 2019. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions // Nature Communications. Vol. 10 (1). P. 590.
- Wilson E. O., 1978. On Human Nature. Cambridge, MA: Harvard University Press. 260p.
- Yunusbayev B., Metspalu М., Järve М. et al., 2012. The Caucasus as an asymmetric semipermeable barrier to ancient human migrations // Molecular biology and evolution. Vol. 29. No. 1. P. 359–365.
