Генетическое разнообразие в популяциях кур русская белая, пушкинская и корниш на основе анализа гомозиготных районов

Автор: Рейнбах Наталья Романовна, Вахрамеев Анатолий Борисович, Рябова Анна Евгеньевна, Макарова Александра Владимировна, Федорова Зоя Леонидовна
Журнал: Молочнохозяйственный вестник @vestnik-molochnoe
Рубрика: Сельскохозяйственные и ветеринарные науки
Статья в выпуске: 3 (47), 2022 года.
Бесплатный доступ
Проведено сравнительное изучение распределения гомозиготных районов в генофондных породах кур для выявления связи между результатами селекции и признаками продуктивности. Получены новые данные по генетической архитектуре распределения гомозиготных районов в популяциях пород русская белая, корниш и пушкинская. Полученные новые знания о генах, связанных с продуктивностью птиц способствуют более широкому мониторингу популяций и внедрению в селекционно-генетический процесс.
ДНК, SNP, Gallus gallus, ROH
Короткий адрес: https://sciup.org/149141106
IDR: 149141106 | УДК: 575.222.5-636.033 | DOI: 10.52231/2225-4269_2021_3_131
Genetic diversity in populations of Russian White, Pushkin and Cornish chickens on the basis of homozygous areas analysis
A comparative study of the distribution of homozygous regions in the gene pool breeds of chickens was carried out to identify the relationship between the selection results and productivity traits. New data on the genetic architecture of the distribution of homozygous regions in the populations of the Russian White, Cornish and Pushkin breeds have been obtained. The obtained new knowledge about the genes associated with the productivity of birds contributes to a wider monitoring of populations and implementation in the selection and genetic process.
Текст научной статьи Генетическое разнообразие в популяциях кур русская белая, пушкинская и корниш на основе анализа гомозиготных районов
Технология SNP-скринирования является высокоэффективным инструментом генетического анализа, способным выявлять структурные особенностиучастков генома для использования вразведенииживотных. Сопутствующим феноменом селекции и воспроизводстве популяций с малой численностью является инбридинг. Для оценки инбридинга необходимо идентифицировать районы генома с гомозиготными SNP [1, 2]. Сравнительная оценка особенностей распределения гомозиготных районов в геноме кур является важным источником информации об изменчивости генофонда в исследуемой популяции и положительном ассортативном скрещивании: инбридинге, селекционном давлении [3, 4, 5]. Это особенно желательно при разведении малочисленных групп для оценки степени инбридинга [6, 7].
Идентификация протяженных гомозиготных сегментов генома (ROH) свидетельствует о наличии инбридинга на относительно близких родственников [8]. Эти ROH-сегменты представляют особый интерес и встречаются в одних и тех же районах хромосом у многих видов животных. Длина гомозиготных районов (ROH) в геноме конкретного животного зависит от направления и интенсивности отбора, дрейфа генов и эффективной численности исходной популяции [6]. Длинные ROH типичны для инбредных особей. Напротив, короткие ROH могут использоваться как индикаторы более «древнего» или «стихийного» инбридинга [7, 9, 10, 11].
В связи с этим изучение гомозиготных районов в породах редких и исчезающих пород кур представляет интерес.
Русская белая порода представляет очень важный элемент анализа ROH. Порода создана путем поглотительного скрещивания белыми леггорнами отечественных кур. Особый интерес представляет популяция Русской белой породы ВНИИГРЖ (Русская белоснежная). История ее формирования интересна и применением очень высокой степени инбридинга при её выведении, высочайших уровней отбора на различные показатели на разных этапах селекции. Так, структура породы основывалась на двух линиях: 9787, имеющей генеалогические ветви трёх петухов: 9187, 3906, 3939. Вторая линия 1365 имеет в своей основе две ветви: 9507 и 9525. [12]
Высокие уровни отбора по гипотермии 39,1% в первый год селекции и затем три года подряд (63,3%, 56,1%, 56,4%), сочетание массового отбора с индивидуальным подбором пар, затем использование семейной селекции при отборе на устойчивость к лейкозу и сочетания семейной селекции использованием уникального петуха 9787, потомство которого оказалось очень устойчиво к болезни Марека, привело к наличию высокой степени инбридинга в популяции русской белой породы кур.
Анализ ROH в этой породе может выявить наличие следов селекции, проводимой в период становления породы и отделить от гомозиготности, появившейся в последние периоды работы с этой породой.
Пушкинская порода кур, утверждённая 11.05.2007 с датой приоритета 17.07.2006, также имеет интересное происхождение. Сочетание белого леггорна – породы яичного направления с генофондом цветных финальных мясных гибридов «Бройлер-6», имеющим тесное родство с корнишами, позволяет предполагать наличие выраженной гетерозиготности. Поэтому анализ гомозиготных районов в пушкинской породе может позволить исследовать общие корни мясных и яичных кур.
Огромное значение в мясном птицеводстве придается породе корниш. Почти все промышленные кроссы используют генетику породы корниш, и изучение ROH мясной родительской формы имеет большое научное и практическое значение.
Таким образом, сравнительное изучение распределения гомозиготных районов по хромосомам у пород кур русская белая, пушкинская и корниш в результате селекции является актуальным аспектом в исследовании потенциала аборигенных пород кур.
Материалы и методы исследования
Материалом для исследования являлось ДНК от 3 пород кур (Gallus gallus): Русской белой, Пушкинской и Корниш. Птица содержится в биоресурсной коллекции ВНИИГРЖ «Генетическая коллекция редких и исчезающих пород кур» (г. Санкт-Петербург ‒ Пушкин) и входит в «Сетевую биоресурсную коллекцию сельскохозяйственных животных, птиц, рыб и насекомых» [13]. Это уникальная база для исследований, которая в настоящее время включает в себя 40 пород и популяций кур различного направления продуктивности.
Популяция русских белых кур (Русская белоснежная) создана в период 1954‒1970 гг. во ВНИИГРЖе, устойчивая к неоплазматическим заболеваниям и к выращиванию цыплят от 1 до 10 дней при температуре не более 24 °С, более низкому температурному фону при выращивании молодняка и содержанию взрослых кур в птичниках без обогрева. Данные куры представляют интерес для биотехнологической промышленности для изготовления вакцин, сывороток и других медицинских препаратов.
Пушкинская порода кур выведена в Экспериментальном хозяйстве ВНИИ генетики и разведения сельскохозяйственных животных за 1976‒2005 годы на основе поглотительного скрещивания экспериментальной популяции черно-пёстрых австралорпов с курами породы Белый леггорн. В ходе становления породы проводилось вводное скрещивание с московскими белыми курами, а также цветными финальными гибридами кросса «Бройлер-6», созданного на основе пород Плимутрок и Корниш [14].
Куры Корниш ‒ широко известная мясная порода, имеющая высокий генетический потенциал мясной продуктивности. Представители этой породы выведены английскими селекционерами и отличаются высокой скоростью роста, достижением очень высокой живой массы, хороших мясных качеств (высокий убойный выход, соотношение массы мышц к общей массе и т.д.). Скрещивание птиц породы Корниш с некоторыми другими породами и мясными гибридами позволяет получать очень продуктивные промышленные бройлерные кроссы. Описание породы обращает внимание на массивность тела и некоторые другие особенности кур.
Скрининг образцов ДНК проводился микрочипом Illumina Chicken 60K SNP iSelect BeadChip («Illumina», США). Качество генотипированных SNP-локусов контролировалось при помощи программы PLINK 1.9. Для анализа отбирались образцы ДНК с качеством генотипирования по SNP-локусам более 95%, что оценивалось с использованием программы Genome Studio («Illumina», США). Поиск гомозиготных районов по отдельным хромосомам проводился с помощью программы PLINK 1.9 и c помощью библиотеки detectRuns в Rstudio согласно следующим параметрам: размер окна 15 SNP, порог перекрытия окон 0.1, минимальное число SNP в районе 15, максимальное число гетерозиготных SNP в окне ‒ 1.
Результаты
На первом этапе проводилось изучение полногеномной архитектуры популяций изучаемых пород по распределению гомозиготных районов. Было выявлено внутрипородное сходство особей и дифференцированность популяций друг от друга. В таблицах 1, 2, 3 представлены результаты проведенного анализа распределения гомозиготных районов.
Было изучено распределение гомозиготных районов по хромосомам в породах кур Русская белая, Пушкинская и Корниш. Выявлены гомозиготные районы с частотой встречаемости 50% и выше на хромосомах 1‒9, 11, 12 и 15. Наивысшие показатели FROH отмечены у кур породы Корниш на хромосамах 5 и 11 (0,24±0,01 и 0,24±0,03 соответственно). На хромосоме 1 наибольшее количество гомозиготных районов выявлено у кур этой же породы (6,73±0,46), при этом количество ROH у двух других пород на данной хромосоме в 1,7 и 1,8 раз этот показатель ниже (у Пушкинской ‒ 3,6±0,54; у Русской белой ‒ 3,89±0,36). На хромосомах 2–9, 11 и 12 наибольшее количество гомозиготных районов у кур породы Корниш, а на хромосоме 15 – у кур породы русская белая. Наименьшее количество ROH отмечено на хромосомах 1, 2, 5, 6, 8, 9, 11, 12 у кур породы Пушкинская, на хромосоме 15 ‒ у кур породы корниш и на хромосомах 3, 4, 7 наименьшее количество гомозиготных районов у кур породы русская белая.
История накопления большого количества ROH-районов у кур породы Корниш обусловлена интенсивным промышленным использованием породы, широким использованием инбридинга, часто в очень высокой степени, вплоть до тесного и высоком селекционном давлении при разведении.
У Русской белой выявленные ROH-районы предположительно связаны с историей выведения породы. Ранее в исследовании Е.С. Федоровой (2022) были выявлены гены-кандидаты у кур породы Русская белая, предположительно связанные с их адаптацией к низким температурам. Двенадцать приоритетных генов-кандидатов на специфичных для породы островах ROH, которые могут быть потенциальнымигенами-кандидатами,связаннымисвысокойадаптивной способностью кур RW в холодных условиях. Гены-кандидаты, связанные с метаболизмом липидов (SOCS3, NDUFA4, TXNRD2, IGFBP1, IGFBP3), поддержанием температуры тела на холоде (ADIPOQ, GCGR, TRPM2), несократительным термогенезом (RYR2, CAMK2G, STK25) и развитием мышц (METTL21C) представляют наибольший интерес [15].
У кур породы Пушкинская накопление гомозиготных районов, возможно, связано с интенсивным отбором на яичную продуктивность и мясные качества. Ранее в исследовании Н.В. Дементьевой и Ю.С. Щербакова на второй, шестой и восьмой хромосомах были найдены гены, отвечающие за продуктивные качества.
На второй хромосоме были обнаружены два локуса, играющие различную функциональную роль. Первый регион включал гены, связанные с обменом кальция, яичной продуктивностью, окраской оперения, репродукцией и иммунитетом. Во второй регион входили гены, отвечающие за формирование яичной продуктивности, роста, метаболизма липидов печени. На шестой хромосоме были аннотированы гены, связанные с развитием мышц, ростом клеток, устойчивостью к стрессу и иммунитетом. На восьмой хромосоме в область селекционного давления (гомозиготный регион) входили гены устойчивости к кислородному голоданию и гены риска опухолей печени у кур [16].
На рисунке показана встречаемость гомозиготных районов на 1 хромосоме исследуемых пород кур. Было отмечено, что на данной хромосоме у более чем 50% особей у пород Корниш и Пушкинская имеются общие локусы накопленния гомозиготных районов в районе участка 17,5‒21,5 Mb (мегабаз). Возможно, в процессе формирования породы пушкинская сохранились и были отобраны особи с гомозиготными участками с таковыми у породы Корниш. Возможно, эти локусы могут служить маркером для отбора более тяжелой птицы в пушкинской породе.
Имеются также общие участки, накопленные у более 50% особей у пород Пушкинская и Русская белая в районе 42,7‒44,7 Mb и 158‒161 Mb. Вероятнее всего, это обусловлено сохранением участков гомозиготности при выведении пушкинской породы с участием белых леггорнов, которые, в свою очередь, были использованы при выведении Русской белой породы. Накопление гомозиготных районов можно также объяснить селекционным давлением при отборе в этой еще молодой породе, который проводился в направлении сохранения мясной продуктивности при увеличении производства яичной продукции. Пушкинская порода кур широко востребована фермерскими хозяйствами как птица, приспособленная к неблагоприятным условиям содержания при сохранении продуктивных качеств.
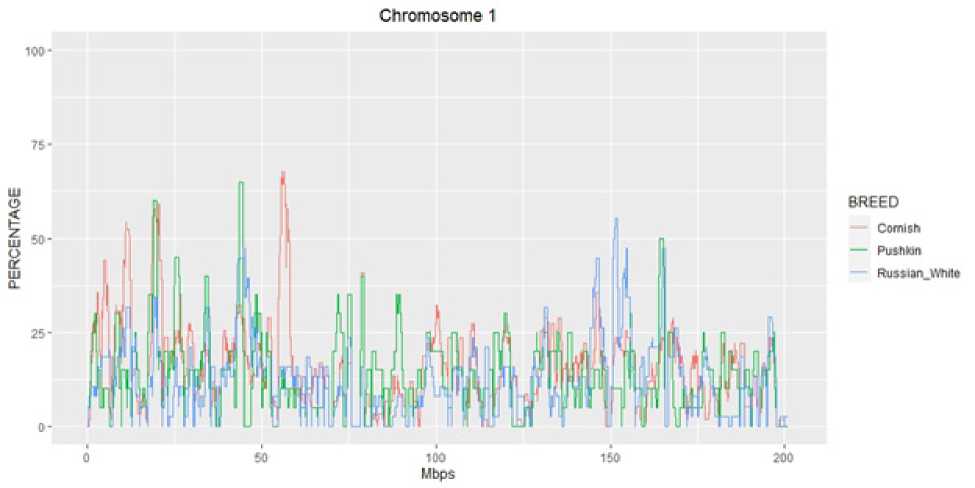
Рисунок 1 - Встречаемость гомозиготных районов на 1 хромосоме в породах кур
Таблица 1 ‒ Протяженность и количество гомозиготных районов в породе Русская белая, n = 30
|
Хромосома |
Количество гомозиготных районов |
Суммарная протяженность гомозиготных районов |
Средняя протяженность района |
F ROH |
|
1 |
3,89±0,36 |
20000±2716 |
4684±406 |
0,1±0,01 |
|
2 |
3,77±0,3 |
19786±1912 |
5326±410 |
0,13±0,01 |
|
3 |
1,7±0,22 |
6897±1096 |
3357±339 |
0,06±0,01 |
|
4 |
2,3±0,25 |
11564±1529 |
4516±524 |
0,12±0,01 |
|
5 |
1,06±0,19 |
5116±1047 |
3122±610 |
0,08±0,01 |
|
6 |
0,93±0,17 |
3261±752 |
1962±360 |
0,08±0,02 |
|
7 |
1,16±0,21 |
4079±964 |
2198±332 |
0,11±0,02 |
|
8 |
1,1±0,25 |
3359±819 |
1723±331 |
0,11±0,03 |
|
9 |
0,87±0,16 |
2898±600 |
2234±482 |
0,12±0,02 |
|
11 |
1,33±1,19 |
4670±482 |
3244±575 |
0,23±0,03 |
|
12 |
0,72±0,08 |
2169±314 |
1511±209 |
0,1±0,02 |
|
15 |
0,73±0,17 |
1573±393 |
994±216 |
0,12±0,03 |
Таблица 2 ‒ Протяженность и количество гомозиготных районов в породе Корниш, n = 22
|
Хромосома |
Количество гомозиготных районов |
Суммарная протяженность гомозиготных районов |
Средняя протяженность района |
F ROH |
|
1 |
6,73±0,46 |
28476±1880 |
4274±133 |
0,15±0,01 |
|
2 |
5,81±0,47 |
29276±2811 |
5069±264 |
0,2±0,02 |
|
3 |
3,09±0,26 |
12549±1135 |
4131±226 |
0,11±0,01 |
|
4 |
4,09±0,4 |
18824±1856 |
4438±303 |
0,2±0,02 |
|
5 |
3,72±0,16 |
14577±775 |
3978±211 |
0,24±0,01 |
|
6 |
1,4±0,16 |
4815±691 |
3075±508 |
0,13±0,02 |
|
7 |
2,5±0,2 |
7107±624 |
2956±188 |
0,19±0,01 |
|
8 |
1,5±0,26 |
4402±778 |
2282±292 |
0,15±0,03 |
|
9 |
0,9±0,2 |
2734±588 |
1993±384 |
0,11±0,02 |
|
11 |
1,78±0,25 |
4827±697 |
2471±348 |
0,24±0,03 |
|
12 |
1,32±0,15 |
3519±468 |
2340±302 |
0,17±0,02 |
|
15 |
0,55±0,14 |
891±241 |
743±185 |
0,07±0,02 |
Таблица 3 ‒ Протяженность и количество гомозиготных районов в породе
Пушкинская, n = 20
|
Хромосома |
Количество гомозиготных районов |
Суммарная протяженность гомозиготных районов |
Средняя протяженность района |
F ROH |
|
1 |
3,6±0,54 |
18571±3667 |
4537±524 |
0,09±0,02 |
|
2 |
3±0,33 |
14834±1595 |
5218±488 |
0,1±0,01 |
|
3 |
2,9±0,32 |
13746±2592 |
4364±653 |
0,12±0,02 |
|
4 |
2,85±0,33 |
12944±1845 |
4519±455 |
0,14±0,02 |
|
5 |
0,7±0,2 |
5346±1939 |
3941±1600 |
0,08±0,03 |
|
6 |
0,85±0,22 |
3263±883 |
211±626 |
0,08±0,02 |
|
7 |
1,2±0,23 |
3767±831 |
2036±407 |
0,1±0,02 |
|
8 |
0,8±0,14 |
2839±760 |
2297±470 |
0,09±0,02 |
|
9 |
0,65±0,17 |
2506±896 |
1782±502 |
0,1±0,04 |
|
11 |
0,95±0,17 |
2702±553 |
2030±394 |
0,13±0,02 |
|
12 |
0,45±0,14 |
1011±308 |
928±285 |
0,05±0,01 |
|
15 |
0,65±0,15 |
1399±363 |
1232±326 |
0,11±0,03 |
Выводы
В результате проведенного исследования гомозиготных районов кур пород Русская белая, Пушкинская, Корниш можно сделать вывод о том, что накопленное большое количество ROH-районов у породы Корниш обуславливается ее спецификой выведения, поскольку данная порода предназначается для промышленного производства. Породы
Пушкинская и Русская белая имеют более разнородную генетическую структуру, что подтверждается меньшим накоплением гомозиготных районов. Дальнейший план исследований предполагает более углубленный анализ отмеченных гомозиготных районов для поиска кандидатных генов и изучения их экспрессии.
Список литературы Генетическое разнообразие в популяциях кур русская белая, пушкинская и корниш на основе анализа гомозиготных районов
- Randhava I.A.S., Khatkar M.S., Thomson P.C., Raadsma H.W. A meta-assembly of selection signatures in cattle//PLOS One. – Vol 11 (4). – 2016. - e0153013.
- Andersson L, Georges M. 2004Domestic-animal genomics: deciphering the genetics of complex traits//Nat Rev Genet. - Vol 5 (3). –. – C.202–212.
- Bosse M., Megens H.J., Madsen O., Paudel Y., Frantz L.A.F., Schook L.B Crooijmans R.P & Groenen M.A. Regions of homozygosity in the porcine genome: consequence of demography and the recombination landscap//PLoS Genetics. - Vol 8(11). – 2012. - e1003100.
- Cardoso T.F., Amills M., Bertolini F., Rothschild M., Marras G., Boink G., Jordana J., Capote J., Carolan S., Hallsson J.H., Kantanen J., Pons A & Lenstra J.A. Patterns of homozygosity in insular and continental goat breeds//Genetics Selection Evolution. - Vol 50 (1). – 2018. – 56c.
- Howard J T., Tiezzi F., Huang Y., Gray K A & Maltecca C. Characterization and management of long runs of homozygosity in parental nucleus lines and their associated crossbred progeny//Genetics Selection Evolution, Vol 48 (1). – 2016. – 91c.
- Kirin M., McQuillan R., Franklin C.S., Campbell H., McKeigue P.M & Wilson J.F. Genomic runs of homozygosity record population history and consanguinity//PLoSOne. – Vol 5 (11). – 2011. - e13996.
- Mastrangelo S., Ciani E., Sardina M.T., Sottile G., Pilla F & Portolano B. Runs of homozygosity reveal genome-wide autozygosity in Italian sheep breeds//Animal Genetics. - Vol 49 (1). – 2018. – C.71–81.
- Peripolli E, Munari DP, Silva MVGB, Lima ALF, Irgang R, Baldi F. Runs of 489 homozygosity: current knowledge and applications in livestock//Animal Genet. – Vol 48. – 2016. – C.255–271.
- Mastrangelo S., Tolone M., Di Gerlando R., Fontanesi L., Sardina M.T & Portolano B. Genomic inbreeding estimation in small populations: evaluation of runs of homozygosity in three local dairy cattle breeds//Animal. - Vol 10 (5). - 2016. – C.746–754.
- Peripolli E., Munari D.P., Silva M V.G.B., Lima A.L.F., Irgang R & Baldi F. Runs of homozygosity: current knowledge and applications in livestock// Animal Genetics. – Vol 48 (3). – 2016. – C.255-271.
- Purfield D.C., Berry D.P., McParland S & Bradley D.G. Runs of homozygosity and population history in cattle//BMC Genetics. - Vol 13. – 2012. – 70c.
- Соколова А. Н. Генетико-селекционные методы создания популяции кур с повышенной устойчивостью к неоплазмам. Дисс. На соиск. уч ст. доктора наук в виде научного доклада//С-Пб-Пушкин – 1999 – 56С.
- Центр коллективного пользования: «Генетическая коллекция редких и исчезающих пород кур» [Электронный ресурс]. – 2016. – URL:https://vniigen.ru/ckp-geneticheskaya-kollekciya-redkix-i-ischezayushhix-porod-kur/ (дата обращения 06.09.2022)
- Патент. Пушкинская. Куры. https://reestr.gossortrf.ru/sorts/9358991/ RU №3633 11.05.2007 с датой приоритета 17.07.2006.
- Fedorova E.S., Dementieva N.V., Shcherbakov Y.S., Stanishevskaya O.I. Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation. // Biology (Basel). – Vol 11 (4). – 2022. – 547с.
- Дементьева Н. В., Щербаков Ю. С., Митрофанова О. В., Вахрамеев А. Б., Хлесткин В. К. Анализ накопления районов гомозиготности у кур пушкинской породы с использованием данных полногеномного генотипирования. // Экологическая генетика. – Vol 20(1). – 2022. – С.31-39
