Holocene fishing in the big sea region of lake Baikal (based on materials from multilayered habitation sites)
")
Author: Nomokonova T.Y., Novikov A.G., Goriunova O.I., Saveliev N.A., Losey R.J., Weber A.W.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Paleoenvironment, the stone age
Article in issue: 4 т.45, 2017.
Free access
Short address: https://sciup.org/145145335
IDR: 145145335 | DOI: 10.17746/1563-0110.2017.45.4.013-023
Text of the article Holocene fishing in the big sea region of lake Baikal (based on materials from multilayered habitation sites)
Fishing is a main subsistence activity among many past and present hunter-gatherer groups. Lake Baikal is one of the most important fishing locations in southeastern Siberia (Fig. 1), and this lake has a wide range of fish species present, including 14 food species and subspecies that are historically considered
to be important dietary sources for Baikal’s human populations (Kozhov, Misharin, 1958: 33–590; Sideleva, 2003: 1–23). Sturgeon ( Acipenser baeri baicalensis ) is one of these species, and inhabits specific parts of the lake and its main tributaries.
The majority of the human-consumed fishes in Lake Baikal can be subdivided as littoral and deep-water species (Kozhov, Misharin, 1958: 101–590: Kozhova, Izmest’eva, 1998: 153–159). For example, the small, shallow, and warm coves of Kurkut and Mukhor gulfs of the Little Sea area (Fig. 1) have year-round inhabitants such as perch ( Perca fluviatilis ), roach ( Rutilis rutilis lacustris ), dace ( Leuciscus leuciscus baicalensis ), and pike ( Esox lucius ). Other species prefer deep and cooler sections of Lake Baikal (such as the Big Sea). Representatives of this group are whitefish ( Coregonus lavaretus baicalensis ), white and black grayling ( Thymallus arcticus baicalensis brevipinnis and Th. arcticus baicalensis ), lenok ( Brachymystax lenok ), and taimen ( Hucho taimen ). The last two species are mainly present in the lake during summer and spend the rest of the year in lake’s tributaries. Whitefish and burbot (Lota lota ) are found in rivers and the lake’s shallow sections mainly during spawning seasons (Kozhov, 1972: 109– 114). White grayling prefers the eastern side of Lake Baikal, but black grayling is widespread throughout the lake and enters its small tributaries for spawning, especially those along the western shore of the Big Sea (Kozhova, Izmest’eva, 1998: 158). Nevertheless, the most well-known species of Lake Baikal is omul ( Coregonus migratorius ), which plays an important role in the contemporary industrial fishery on Lake Baikal.
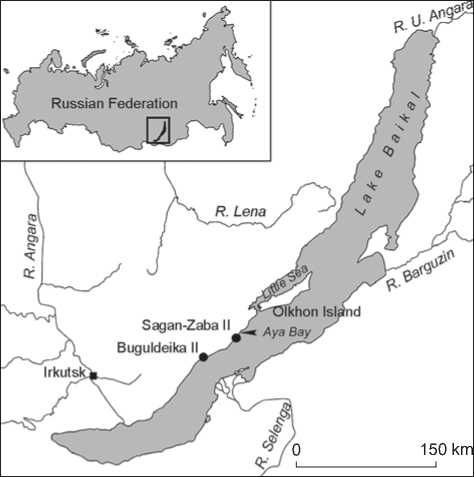
Fig. 1. Map indicating location of the multilayered habitation sites Sagan-Zaba II and Buguldeika II.
Omul is represented by a few populations in the lake, and enters less deep sections of lake and rivers only during the spring-summer spawning migrations (Kozhov, Misharin, 1958: 131–214; Sideleva, 2003: 13–14).
Many researchers had previously addressed questions in regards to the ancient fishing practices carried out on Lake Baikal. Their works were based on typological analysis of tools associated with fishing from habitation sites along the Little Sea shoreline (Svinin, 1976; Novikov, Goriunova, 2005), as well as on the ichthyofaunal remains from these same sites (Tsepkin, 1976; Nomokonova, Losey, Goriunova, 2009a, b, c: 53–75; Nomokonova et al., 2011; Nomokonova, Goriunova, 2012). Multiple publications demonstrated the importance of fishing in lives of Early and Middle Holocene hunter-gatherers inhabiting shores of the Little Sea area. These publications produced lists of identified fish species, possible seasonality indicators, fishing techniques, reconstructed sizes of perch and roach, as well as discussions of changes in uses of fish species and their quantities through different chronological periods (Novikov, Goriunova, 2005; Nomokonova, Losey, Goriunova, 2009a, b, c: 75–91; Losey, Nomokonova, Goriunova, 2008, 2014).
Despite these publications of multiple reconstructions of ancient fishing in the Little Sea area, many problems regarding the use of fish resources in the Big Sea region remain unsolved. This is partially explained by the small amount of multilayered and well-stratified habitation sites containing remains of fishing tools and ichthyofauna along the shores of the Big Sea. This situation changed in 2006–2008 as a result of new excavations conducted by teams of Russian-Canadian expeditions (a joint project between Irkutsk State University and the University of Alberta) at the multilayered habitation sites Sagan-Zaba II and Buguldeika II located on the western shore of Lake Baikal (Fig. 1–3). During these collaborative investigations, a new set of data on ancient fishing in the Big Sea region of the lake was produced. These materials, further discussed in this article, have brought new understanding to the importance of fishing in the subsistence of people living on the shores of Lake Baikal during the Holocene.
Materials and methods
This article is based on the analysis of 1553 ichthyofaunal remains and 39 artifacts associated with fish procurement (hooks, harpoons, net-sinkers, and fish imagery) recovered from excavations of multilayered habitation sites Sagan-Zaba II and Buguldeika II. These sites are located in the central section of the western shoreline of Lake Baikal, about 35 km from each other, and 154 km and 130 km to the northeast from the modern city of Irkutsk. Sagan-Zaba II was discovered for the first time by North-Asian Expedition of IIFF SO AN USSR led by A.P. Okladnikov (Okladnikov, 1974: 17). This expedition also conducted excavations at this location in 1974 and 1975 (Aseev, 2003: 51–61). Buguldeika II was found in 1987 by a team from Irkutsk State University, led by N.A. Saveliev and V.M. Vetrov. This site was further investigated by a joint expedition between the Tourism and Regional Historical Child and Youth Center and the Irkutsk State University, under supervision of V.V. Altukhov and N.A. Saveliev in 1999– 2004. Interdisciplinary studies were carried out at both sites by the Russian-Canadian Expeditions in 2006–2008 (Goriunova et al., 2006, 2008; Bocharova, Korshunov, 2010; Losey, Nomokonova, Saveliev, 2014).
This article analyzes fish remains and fishing items from trenches 4B and 4C at Sagan-Zaba II and trenches 4 and 5 at Buguldeika II. Field methods included stratigraphic excavations and three-dimensional recording of the positions of all cultural remains and sieving of all sediments through screens of 3 mm diameter mesh. This method was very successful, as 99 % of all ichthyofaunal remains were received during the process of sieving. In a few cases, concentrations of fish bones were found. One of them was recorded under the rock of a hearth in layer IV (excavation level 7) at Buguldeika II. In addition, a few Salmonidae vertebrae and bones, partially in anatomical order, were found in the lower layer III, under the remains of a pot, at habitation site Sagan-Zaba II.
Chronological dating of cultural remains at Sagan-Zaba II and Buguldeika II is based on over 90 AMS radiocarbon dates made by Oxford Radiocarbon Accelerator Unit. Detailed analyses of these dates have been discussed in a few publications already (Nomokonova et al., 2013; Losey, Nomokonova, Saveliev, 2014). A summary of the chronological spans of cultural layers for both habitation sites is presented in Table 1. In terms of archaeological periodization, materials are dated from the Mesolithic to the ethnographically contemporary period.
Ichthyofaunal remains were described and identified to the element, portion, side, and to the most specific taxonomic category possible using standard zooarchaeological methods (number of identified specimens and minimum number of individuals), and animal behavioral characteristics were also employed in our interpretations (Nomokonova, Losey, Goriunova, 2006; Lyman, 2008: 21–82; Reitz, Wing, 2008: 142–170).

Fig. 2. View of Sagan-Zaba bay. Photo by A.W. Weber.

Fig. 3. View of archaeological site Buguldeika II. Photo by A.W. Weber.
Methods of working with fishing artifacts were based on typological analysis of items developed for the Cis-Baikal region and on ethnographic analogies (Okladnikov, 1936, 1941, 1950: 246–258; Studzitskaya, 1972, 1976; Novikov, Goriunova, 2005).
Ichthyofaunal remains from Big Sea habitation sites
Fish remains from Sagan-Zaba II and Buguldeika II are represented by 1553 specimens. The majority of them were found at Sagan-Zaba II, but ichthyofauna comprise only 2.1 % of the total faunal remains found at this
Table 1. Chronology of habitation sites
|
Geologic Timescale |
Archaeological Timescale |
Sagan-Zaba II |
Buguldeika II |
|||
|
Cultural layers |
Chronological span, cal BP |
Cultural layers (sublayers) |
Chronological span, cal BP |
|||
|
ф с ф о о о т |
Early |
Mesolithic |
V, IV (9, 8) |
10,410–9030 |
||
|
VII |
9020–8650 |
|||||
|
Middle |
Neolithic |
VI |
8160–7880 |
IV (5–1) |
8610–5590 |
|
|
V lower |
Not dated |
|||||
|
V upper |
6750–6310 |
|||||
|
IV |
5590–4870 |
III |
5660–4650 |
|||
|
Late |
Bronze Age |
III lower |
4440–2000 |
II (3) |
3210–2780 |
|
|
II (2) |
2680–1950 |
|||||
|
Early Iron Age |
III upper |
1970–1540 |
II (1) |
2130–1530 |
||
|
Late Iron Age – ethnographically contemporary period |
II, I |
1230–940 |
I |
2040 – contemporary period |
||
Table 2. Fish species composition and quantities of their remains at Sagan-Zaba II
|
Taxa |
Common name |
Cultural layer |
Total |
||||||
|
VII |
VI |
V |
IV |
IIIl |
IIIu |
II, I |
|||
|
Pisces unidentified |
Fish |
107 |
1 |
14 |
52 |
141 |
107 |
92 |
514 |
|
Salmonidae |
Family of salmonids |
– |
– |
29 |
– |
363 |
428 |
31 |
851 |
|
Coregonus sp. |
Genus of whitefishes |
– |
– |
– |
3 |
8 |
6 |
– |
17 |
|
Thymallus articus |
Grayling |
3 |
– |
– |
– |
1 |
2 |
1 |
7 |
|
Cyprinidae |
Family of cyprinids |
– |
– |
– |
1 |
1 |
– |
– |
2 |
|
Leuciscus baicalensis |
Dace |
– |
– |
– |
– |
1 |
1 |
– |
2 |
|
Rutilus rutilus lacustris |
Roach |
– |
– |
– |
– |
– |
3 |
– |
3 |
|
Acipenser baerii baical. |
Sturgeon |
– |
1 |
10 |
1 |
28 |
5 |
3 |
48 |
|
Esox lucius |
Pike |
– |
– |
3 |
11 |
– |
– |
1 |
15 |
|
Lota lota |
Burbot |
– |
– |
– |
2 |
– |
– |
– |
2 |
|
Perca fluviatilis |
Perch |
– |
– |
31 |
17 |
5 |
2 |
– |
55 |
|
Total |
110 |
2 |
87 |
87 |
548 |
554 |
128 |
1516 |
|
habitation site (total is 74,040 spec.). There were only 37 fish remains found at Buguldeika II, which is only 0.2 % of the total number of animal bones from this location (total is 20,263 spec.).
About 34 % of the ichthyofaunal remains at habitation site Sagan-Zaba II (Table 2) were unsuitable for further identification. Grayling, roach, dace, perch, burbot, pike, sturgeon, as well as members of the genus of coregonids (omul/whitefish), and families of cyprinids and salmonids (grayling/whitefish), were identified at this site. Fish remains were found in each cultural layer of this habitation site, which date to different periods of the Holocene, starting at over 9000 cal BP. They are the most abundant in layer III lower (4440–2000 cal BP)
and III upper (1970–1540 cal BP), and less numerous in layer VI (8160–7880 cal BP) (Fig. 4).
The majority of the identified specimens belong to salmonids. They compose 88 % (875 specimens including the grayling and whitefish/omul categories) of the total number of fish remains found in all cultural layers of Sagan-Zaba II, with exception of layer VI. They are most numerous in lower and upper III layers (Fig. 4). The remaining identified bones and scales are from perch (55 spec.), sturgeon (48 spec.), pike (15 spec.), cyprinids (7 spec., including roach and dace), and burbot (2 spec.). Sturgeon remains were found in almost every layer at this site with exception of layer VII. Perch bones and scales were found in layers spanning from 6750 to 1540 cal BP.
Fig. 4. Relative abundance of salmonid remains and of other species at habitation site Sagan-Zaba II summed by chronological period. a – Salmonidae; b – other; c – total.
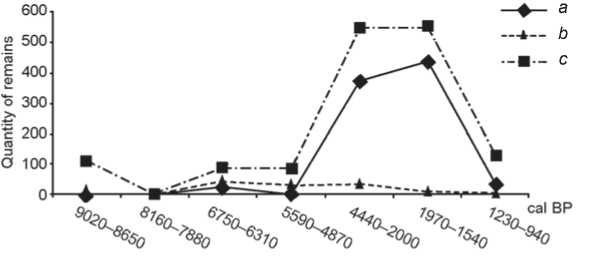
Pike remains were mostly identified in deposits associated with the middle portion of the Holocene (layers V and VI), and were found in very small numbers in layers II and I (1230 – 940 cal BP). Cyprinids and burbot bones are rare in comparison to those of other fish species. Cyprinids were found in layers IV, III lower, and III upper. Remains of burbot were identified only in layer IV (5590–4870 cal BP).
The majority of the ichthyofaunal remains from Buguldeika II (37 spec.) are remains of sturgeon ( Acipenser sp.) with a total of 25 specimens. One pike bone ( Esox lucius ) and two from family of cyprinids (Cyprinidae) were also found. The remaining nine fragments were not suitable for further identification. Many of ichthyofaunal remains, including those from pike and sturgeon, were found in layer IV (22 specimens; sublevels 2, 3, 6–8, all with a chronological span from 10,410 to 6570 cal BP). Layer II contained mainly remains from sturgeon but also two bones from cyprinids. Only one unidentified fish bone was found in layer I. These layers have chronological spans starting from 3210 cal BP through to the ethnographically contemporary period.
Fishing artifacts from habitation sites
Excavations at habitation sites Sagan-Zaba II and Buguldeika II produced 39 artifacts associated with fishing (21 and 18 items, respectively). These include stone net sinkers, fragments of antler and bone harpoon heads, slate shanks of composite fishhooks and their preforms, bone hook or barbs for fishhooks, a small nephrite sinker, and stone fish images (Table 3, Fig. 5).
Fishing artifacts are found at Sagan-Zaba II starting with the layers dating to the Early Holocene. A bilateral and symmetrically barbed antler harpoon head fragment, with splayed base and a hole for attachment for a line, along with a pebble net sinker with notches on opposite sides, were recovered from layer VII.
Sagan-Zaba II Middle Holocene deposits contained 18 artifacts associated with fish procurement. They were found in cultural layers associated with different periods of the Neolithic and chronologically span from 8160 to 4870 cal BP. All layers contained shanks of composite fishhooks, bone harpoon head fragments, and fish images. In terms of numbers, fishhook technology is predominant and includes eight (whole and fragmented) shanks of composite fishhooks and a tip of a single bone hook. Almost all shanks are made from slate, with exception of one bone item from layer IV. These shanks of composite fishhooks have different typological forms associated with different periods of the Neolithic. Layer VI had a curved shank that was 3.2 cm in length and with notches on its upper end and a lateral form of attachment for a hook (the so called Baikal type). A complete shank of similar
Table 3. Fishing tools from habitation sites Sagan-Zaba II and Buguldeika II
|
Artifacts |
Sagan-Zaba II |
Buguldeika II |
||||
|
Early Holocene |
Middle Holocene |
Late Holocene |
Middle Holocene |
Late Holocene |
Total |
|
|
Harpoon heads |
1 |
4 |
1 |
– |
– |
6 |
|
Shanks of composite fishhooks |
– |
8 |
– |
10 |
– |
18 |
|
Hooks for composite fishhooks |
– |
1 |
– |
– |
– |
1 |
|
Net sinkers |
1 |
– |
– |
7 |
1 |
9 |
|
Small net-sinkers |
– |
1 |
– |
– |
– |
1 |
|
Fish images |
– |
4 |
– |
– |
– |
4 |
|
Total |
2 |
18 |
1 |
17 |
1 |
39 |
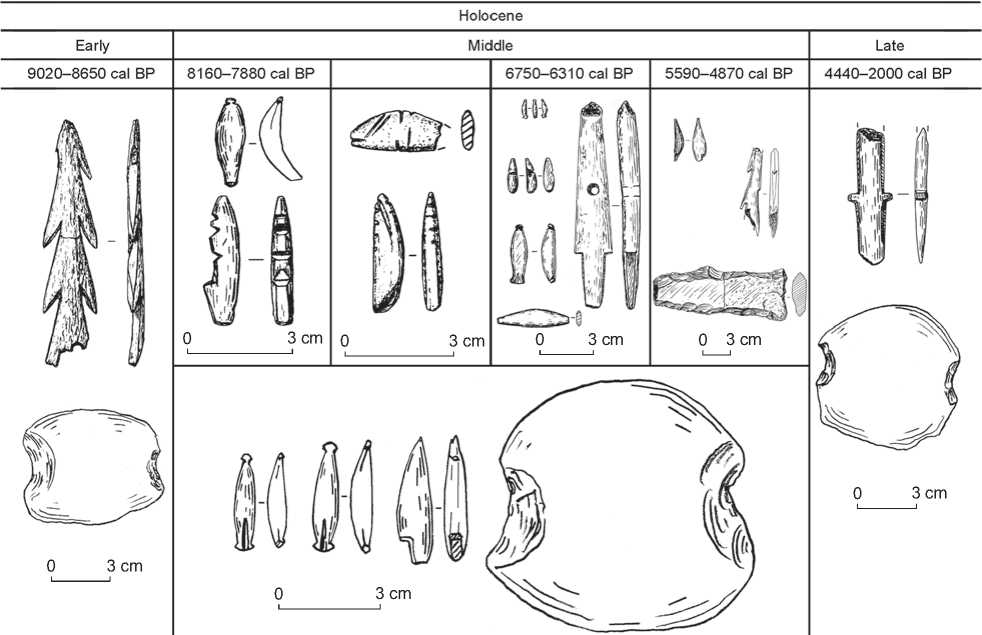
Fig. 5. Distribution of fishing artifacts from habitation sites Sagan-Zaba II and Buguldeika II summed by chronological period.
size (3.3 cm) from layer V lower is straight with convex back, lateral attachment for a hook, and notches on the upper ends for attachment of a line. Its surface is covered with several incisions. Fragments from two shanks were also found in this layer. They have sharpened upper ends, and two incisions are present on one of them. A classic Kitoy shank of a composite fishhook was found in the V upper layer. It is straight with convex back, has halfmoon shaped ends, and frontal attachment for a hook. Its length is 3.5 cm. Another two shanks, one of which is a fragment, are straight with convex backs. The complete shank has a half-moon shaped upper head and a pointed lower end. The complete shank is 1.1 cm long; a fragment is 1.8 cm long. A bone shank of a composite fishhook from layer IV has a convex back and lateral attachment for a hook. Its upper end is sharpened but lower end is widened. The length of this shank is 4.5 cm.
A small nephrite sinker (shank?) was found among the materials of the V upper layer. It has a straight shape with convex back. One end is sharpened; the other end is of oval shape (widened). The middle portion of its back has a half-circle shaped incision. The length of this item is 2 cm.
Bone harpoon heads are all fragmented in the Neolithic layers. One has unilateral barbs (layer VI). Two others are bilaterally barbed (layer IV). Another harpoon fragment was found in the V upper layer and has a tapering base and a hole in its body for a line attachment.
Fish images are also associated here with the process of fishing (4 items). One of them was found in the V lower layer. This item is made from marble and is a stylistic depiction of a fish with a convex back. Its tale is broken off. It has bilateral modelling. An incised line is used to show its mouth. Double incised lines are drawn to show its gills. Incised lines are also visible in the area of its dorsal and pelvic fins. The length of this fish image is around 3 cm. A second fish image, also from marble, has a convex back and a length of 4 cm. It was found in the V upper layer. The image can be described as schematic. Layer IV contained a complete stone fish image of whitefish shape with a length 14.5 cm, as well as a fragment of a tail from another fish image.
Late Holocene deposits at Sagan-Zaba II produced only one artifact associated with fishing. It is a bone harpoon head fragment with straight base with bilateral line guard projections, and was from the layer III lower.
All fishing artifacts at habitation site Buguldeika II (17 out of total 18 items) were found in layers dating to the Middle Holocene. Many of them are shanks


0 5 cm 6
Fig. 6. Shanks of composite fishhooks ( 1–4 ) and stone net sinkers ( 5 , 6 ) from layer IV of habitation site Buguldeika II. Photo by S. Kogai.
of composite fishhooks (6 items) and their pre-forms (4 items). All complete shanks are of the classic Kitoy type. They are straight with convex backs and half-moon projections on both ends (Fig. 6, 1–4 ). The attachment for the hook is frontal. The size range of the length of these items is 2.7–3.2 cm. The shank pre-forms from layer IV (sublevel 7) has a lateral attachment for a hook. One end of it has a few incisions. Seven net sinkers made from flat stones and pebbles were found in layer IV (sublevel 4) (Fig. 6, 5 , 6 ). They are shaped by bilateral notches located on their opposite sides.
One net-sinker was found in the Late Holocene layer II (sublevel 2). It is a flat stone with notches on opposite ends.
Discussion
New data generated from the multilayered habitation sites Sagan-Zaba II and Buguldeika II, located on the western shore of the Big Sea area of Lake Baikal, allowed us for the first time to not only identify species composition and reconstruct fishing techniques, but also to trace changes in use of different fish species through time. Faunal collections contain both littoral and deep-water species such as perch, cyprinids (roach and dace), pike, burbot, sturgeon, and salmonids (grayling and coregonids). The latter group prefers the lake’s deep and cold sections, and these fish are predominant in terms of quantitative estimates among the fish remains at Sagan-Zaba II. Salmonids were not identified at Buguldeika II.
Comparison of ichthyofaunal remains from habitation sites of the Big Sea and Little Sea regions demonstrated substantial differences in their quantitative compositions (Fig. 7). Many of these differences could be explained by location of the habitation sites either near shallow or deepwater sections of the lake’s. For example, ichthyofaunal composition in the Little Sea area includes mostly littoral fish species such as perch, cyprinids, and pike. Coregonid remains are rare, and perhaps, these species were entering
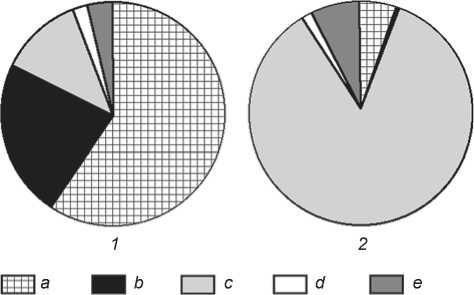
Fig. 7. Quantitative composition of ichthyofaunal remains from the habitation sites of the Little Sea region ( 1 ) and habitation sites of the Big Sea region ( 2 ).
a – perch; b – cyprinids; c – salmonids; d – pike; e – sturgeon.
to coves of this region only during their spawning migrations (Nomokonova, Losey, Goriunova, 2009a, b: 75–85; Nomokonova et al., 2011). Remains of salmonids are predominant at habitation site Sagan-Zaba II, including grayling bones, remains of which were found for the first time in the Olkhon region.
At the same time, there are notable differences between the two habitation sites analyzed in this article. For example, Sagan-Zaba II materials contained variable fish species, including littoral and deep-water ones. Remains of burbot were found here as well, whose bones, just as those from grayling, were found for the first time at habitation sites in the Olkhon region. Ichthyofaunal remains from Buguldeika II were present in substantially lower quantities, and its species composition was also more limited, with just bones and scales from sturgeon, cyprinids, and pike being present.
Analysis of species composition and richness of ichthyofauna by chronological periods shows both similarities and substantial differences between habitation sites located in the Big Sea and Little Sea regions. For example, fishing activities are traced at both locations throughout the whole period of Holocene starting at over 9000 BP. The differences between both regions are in terms of more or less intensive periods of fishing. Fish remains are predominant at the Little Sea habitation sites mostly in deposits of Middle Holocene and earlier, starting approximately at 8000 BP. However, this situation occurs in the Big Sea area mainly in the Late Holocene layers (especially from 4440 to 1540 cal BP).
Ethnohistoric records often indicate that Cis-Baikal pastoralists and those from neighboring regions despised fish and conducted fishing only when they had to (the exception to this was the industrial fishing by Buryat populations inhabiting Baikal shores since the 17th century AD) (Levin, 1897; Mikhailov, 2006: 109–121). However, large numbers of fish remains in the Late Holocene deposits at habitation site Sagan-Zaba II (especially in layers from the Iron Age and ethnohistorically contemporary layers such as III upper, II, and I) clearly demonstrate that fishing traditions of Early and Middle Holocene hunter-gatherers on Lake Baikal had continued among its pastoralists.
Fishing artifacts from the archaeological sites of the western shore of the Big Sea region of Lake Baikal appear in the Early Holocene deposits. Items from layer VII at Sagan-Zaba II allowed for the reconstruction of fishing techniques that existed during the Mesolithic, starting at 9020 cal BP. It is possible that ancient populations speared fish with harpoons and caught them with the use of nets. The last technique is supported by the presence of specially made stone net sinkers found at the habitation site. The use of nets assisted in more effective and productive fishing. Habitation sites of Little Sea also have evidence towards the use of nets in the materials of Late Mesolithic (Novikov, Goriunova, 2005; Nomokonova, Goriunova, 2012).
Neolithic (Middle Holocene) deposits at the analyzed habitation sites of the Big Sea region are distinguished by the numbers of recovered artifacts associated with fishing, with Sagan-Zaba II showing the most variety of such items. Hook and line fishing (angling) items are widespread and demonstrate a presence of individual fishing tools (fishing rods). This is supported by findings of shanks from composite fishhooks and their bone hooks. Shanks with lateral attachment for a hook (the so called Baikal type) are found in deposits throughout the Middle Holocene. Shanks of the Kitoy type with a frontal attachment for a hook (V upper layer of Sagan-Zaba II, and IV layer of Buguldeika II) co-exist with the Baikal type in the chronological span from 8610 to 6310 cal BP. The role of fishing in ancient subsistence along the shores of Lake Baikal is also demonstrated by findings of stone fish images (layers V and IV at Sagan-Zaba II). These stone images, based on ethnographic analogies, were used as fish lures during fishing with harpoons (Okladnikov, 1941, 1950: 246– 258; Studzitskaya, 1976). Variability of technical gear found at habitation sites is evidence of different fishing techniques: the use of the harpoon in shallow sections, and the fishing rod and net in deep-water areas. It is also noticeable that transportation technology (watercraft) is required to fish for deep-water species on Lake Baikal during the summer period.
Comparison of fishing gear found in the Middle Holocene deposits between the habitation sites of the Big Sea region and those from the Little Sea shores showed their similarities (presence of hook and line technologies, composite fishhooks, findings of stone fish images, the large number of harpoon head fragments, and net sinkers). Also repetitive are main typological characteristics of some of these artifacts (shanks of the Baikal type, stylized schematic fish images, whitefish – like fish, etc.) (Novikov, Goriunova, 2005). The presence of Kitoy type shanks of composite fishhooks is new for the western shores of Big Sea, as these types are not found at habitation sites of the Little Sea region.
Fishing also did not lose its importance among the local populations of the Late Holocene. For example, Bronze Age deposits (sublayer 2 of layer II at Buguldeika II, and III lower layer at Sagan-Zaba II) contained a stone net sinker and a harpoon fragment with straight base and line guard projections. It is also interesting that layer III lower at Sagan-Zaba II contained a large number of ichthyofaunal remains, but only a few artifacts associated with fishing. The layer just above it (III upper, Early Iron Age) contained no fishing items at all. It is possible that fishing was done by different types of gear that was not preserved (seine and other nets).
Based on analysis of fishing items and faunal remains from the habitation sites located in the Big Sea region of Lake Baikal, their habitants carried out fishing throughout the Holocene. It is also interesting that despite the importance of fishing in the subsistence of the ancient Baikal populations, this activity is not commonly shown in local rock art images. Fishing is only shown at the rock art panel in Aya bay, located on the western shore of Lake Baikal, about 16 km and 51 km to the northeast from habitation sites Sagan-Zaba II and Buguldeika II, respectively (Fig. 1). The rock art panel has depictions of two realistic style fish images consisting of a smoothed depiction (Fig. 8). Based on its outline (a narrow pointing head, specific body shape), these drawings appear to represent sturgeon or sterlet (Okladnikov, 1974: 36–37). Fish images (along with other animals—snakes and birds) are depicted near the horned anthropomorphic image, perhaps representing a shaman. Rock art images at Aya bay are dated to the Bronze to Early Iron Ages. Perhaps, they are associated with beliefs of ancient populations about Lower (fish and snake) and Upper (birds) Worlds, suggesting a mythological meaning for the image.
Conclusions
The first data on ichthyofaunal remains from the western shore of the Big Sea region of Lake Baikal were generated as a result of excavations at the multilayered habitation sites Sagan-Zaba II and Buguldeika II. This was made possible due to the field excavation methods, which were based on sieving of all archaeological sediments through screens of 3 mm diameter. This method showed that 99 % of all ichthyofaunal remains came from the screens. Application of this technique illustrates a need to use sieving in all further archaeological excavations at other archaeological sites in this region (Nomokonova, Losey, Goriunova, 2006, 2009: 51–53; Nomokonova et al. 2010).
Analysis of ichthyofaunal and fishing artifacts from the multilayered habitation sites Sagan-Zaba II and Buguldeika II produced new interesting data on fish procurement in the Big Sea region of Lake Baikal throughout the Holocene. Fishing as a distinct subsistence strategy starts in this region during the Early Holocene (Mesolithic). Improvements to fishing technologies are noticeable during the Middle Holocene (Neolithic) by the appearance of new types of fishing artifacts and evidence for variable fishing techniques. Different typological characteristics of shanks of

Fig. 8. Rock art images from Aya bay. Photo by A.G. Novikov.
composite fishhooks, harpoons, and fish imagery are identified based on their presence during different chronological periods within the Neolithic. For the first time, reliable information on Baikal fishing, not only during the Bronze Age but also during the Iron Age, was produced as a result of excavations at habitation sites Sagan-Zaba II and Buguldeika II.
New materials from the western shores of the Big Sea region allowed us to compare existing data on fishing from the Little Sea habitation sites and to identify major differences in fish species composition, periods of intensive use of habitation sites, and main subsistence patterns. It has been demonstrated that deep-water fish species, such as salmonids and sturgeon, were procured mostly in the Big Sea region, but littoral and shallowwater species, such as perch, cyprinids, and pike, were fished mainly in the Little Sea region.
Further, differences were identified between these two regions and between chronological periods of more or less intensive fishing. Fish remains in the cultural layers of Middle Holocene and earlier periods are predominant at the Little Sea habitation sites; however, at the Big Sea habitation sites, this situation is more common in the Late Holocene layers. It is also noticeable that ichthyofaunal remains from the Big Sea region are found in small quantities (2.1 % at Sagan-Zaba II and 0.2 % at Buguldeika II) relative to the total number of faunal materials found at these sites. It looks like Baikal seals were the focus of subsistence activities at these locations, which have been previously discussed several times in our other publications (Losey, Nomokonova, Saveliev, 2014; Nomokonova et al., 2015). Fishing was of lesser importance at habitation sites Sagan-Zaba II and Buguldeika II. However, at the Little Sea habitation sites, fishing played a key role in the subsistence
(Nomokonova, Losey, Goriunova, 2009a, b, c: 75– 91; Nomokonova, Goriunova, 2012). Overall, Baikal populations living on shores of Lake Baikal during the Holocene had subsistence patterns composed of complex use of natural resources with fishing playing a significant role.
Acknowledgements
Zooarchaeological analyses were supported by the Social Sciences and Humanities Research Councils’ Major Collaborative Research Initiatives (MCRI SSHRC No. 4122011-1001) of Canada; archaeological examinations were supported by the Russian Science Foundation (Project No. 14-50-00036).
