Идентификация Acarapis woodi и Varroa destructor у Apis mellifera с использованием экзогенной ДНК ульевого мусора

Автор: Шамаев Н.Д., Шуралев Э.А., Мукминов М.Н.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Зоотехния и ветеринария
Статья в выпуске: 11, 2025 года.
Бесплатный доступ
Цель исследования – получение генетической информации о циркулирующих гаплотипах A. woodi и V. destructor с использованием экзогенной ДНК на отдельных пасеках Республики Татарстан. Задачи: определение уровня заражения V. destructor и A. woodi на отдельных пасеках в районах Республики Татарстан с циркулирующих гаплотипов и обнаружение следов заражения в колониях медоносных пчел с использованием экзогенной ДНК ульевого мусора. Объекты исследования – A. woodi и V. destructor в колониях A. mellifera на отдельных пасеках в Республике Татарстан в 2024 г. Пчелы подвидов A. m. mellifera и A. m. carnica были собраны в 13 районах Республики Татарстан в период с марта по июнь 2024 г. На одной пасеке в одном районе зафиксирован низкий уровень заражения клещом V. destructor; обнаружен один новый гаплотип, схожий с изолированными в других странах; на двух пасеках в двух районах зафиксирован низкий уровень заражения клещом A. woodi; обнаружен один гаплотип, идентичный изолированным в других странах и один уникальный гаплотип; несмотря на высокую эффективность пчеловодческих практик в Республике Татарстан, экзогенная ДНК ульевого мусора позволила обнаружить следы заражения клещами (ген cox1 мтДНК) в колониях медоносных пчел на шести пасеках в шести районах; была обнаружена зависимость между обнаружением гена cox1 мтДНК A. woodi и V. destructor в экзогенной ДНК ульевого мусора и подвидом A. m. mellifera в исследованных колониях.
Эктопаразиты, Varroa destructor, Acarapis woodi, экзогенная ДНК, медоносная пчела, Apis mellifera, гаплотип
Короткий адрес: https://sciup.org/140313047
IDR: 140313047 | УДК: 575.8:638.123:638.147:638.157 | DOI: 10.36718/1819-4036-2025-11-120-133
Acarapis woodi and Varroa destructor identification in Apis mellifera using exogenous DNA from hive debris
The aim of the study is to obtain genetic information on circulating haplotypes of A. woodi and V. destructor using exogenous DNA from selected apiaries in the Republic of Tatarstan. Objectives: to determine the level of V. destructor and A. woodi infection in selected apiaries in the districts of the Republic of Tatarstan using circulating haplotypes and to detect traces of infection in honey bee colonies using exogenous DNA from hive debris. The objects of the study were A. woodi and V. destructor in A. mellifera colonies in selected apiaries in the Republic of Tatarstan in 2024. Bees of the A. m. mellifera and A. m. carnica subspecies were collected in 13 districts of the Republic of Tatarstan from March to June 2024. Low levels of V. destructor mite infestation were recorded in one apiary in one district; one new haplotype similar to those isolated in other countries was detected; low levels of A. woodi mite infestation were recorded in two apiaries in two districts; one haplotype identical to those isolated in other countries and one unique haplotype were detected; despite high efficiency of beekeeping practices in the Republic of Tatarstan, exogenous DNA of hive debris allowed detection of traces of mite infestation (mtDNA cox1 gene) in honey bee colonies in six apiaries in six districts; a relationship was found between detection of A. woodi and V. destructor mtDNA cox1 gene in exogenous DNA of hive debris and the A. m. mellifera subspecies in the studied colonies.
Текст научной статьи Идентификация Acarapis woodi и Varroa destructor у Apis mellifera с использованием экзогенной ДНК ульевого мусора
Funding: this work was supported by a grant from the Academy of Sciences of the Republic of Tatarstan, awarded to young candidates of science (postdoctoral students) for the purpose of defending their doctoral dissertations, conducting research, and performing work functions in scientific and educational organizations of the Republic of Tatarstan within the framework of the State Program of the Republic of Tatarstan "Scientific and Technological Development of the Republic of Tatarstan".
Введение. Несмотря на то что число колоний медоносных пчел выросло во всем мире, за последние несколько десятилетий продолжают публиковаться сведения об увеличении потерь среди местных колоний [1–6]. Кроме того, спрос на услуги по опылению вырос непропорционально росту колоний медоносных пчел [7, 8]. В этом контексте многочисленные паразиты и заболевания угрожают здоровью медоносных пчел, поэтому обнаружение паразитических клещей, таких как Varroa destructor и Acarapis woodi, в колониях медоносных пчел (Apis mellifera) имеет решающее значение для предупреждения нехватки этих важных опылителей сельскохозяйственных культур [1–10].
Эти эктопаразиты вносят значительный вклад в ежегодное сокращение колоний хозяина. V. destructor , паразитический клещ, губительный для медоносных пчел ( Apis mellifera ), питается гемолимфой и жировым телом пчел и осуществляет передачу патогенных вирусов. В исследованиях сообщается о резистентности к акарицидам (например, флувалинату), что затрудняет контроль за распространением клеща.
Возникнув как паразит A. cerana в Азии, V. destructor перешел на A. mellifera в середине XX в. Дальний Восток является первоначальной точкой проникновения V. destructor в 1970-х гг., вероятно, через трансграничную торговлю с Ки-таем/Северной Кореей [11]. В 1970-х годах он был обнаружен в Приморском крае. В 1980– 1990 гг. он распространился на запад, достигнув европейской части России к 1990-м гг. A. woodi поражает трахеи взрослых пчел, нарушая дыхание, впервые был обнаружен в России в начале XX в. после его глобального распространения в качестве вредителя медоносных пчел и идентификации в Великобритании в 1919 г. [12, 13]. К 1930-м гг. заражения данным паразитом были зарегистрированы в европейской части России, вероятно, благодаря завезенным европейским породам пчел. Советские записи о пчеловодстве указывают на локальные вспышки акарапидоза на Кавказе и в ЦентральноЧерноземных регионах, где умеренный климат и высокая интенсификация пчеловодства способствовали распространению клещей. Однако едва заметные симптомы акарапидоза и отсутствие стандартизированной диагностики привели к занижению данных до 1980-х гг., когда широкое распространение начали получать передовые методы микроскопии. После обнаружения акарапидоза Л.И. Перепеловой в Тульской области в 1926 г. в дальнейшем случаи заболевания были зарегистрированы в 18 регионах современной территории России. Большая часть находок была отмечена в европейской части России, тогда как в азиатской части находок не было. Обширная территория России и климатическое разнообразие создали неравномерную распространенность A. woodi . Популяции диких пчел (например, A. m. mellifera ) могут выступать в качестве резервуаров, хотя данных мало.
Традиционные методы обнаружения V. destructor и A. woodi (визуальная оценка, диссекция пчелы и микроскопия) являются трудоемкими, отнимают много времени и не обладают эффективностью, особенно на ранних стадиях заражения. Молекулярные методы с использованием экзогенной ДНК, определяемой здесь как генетический материал, происходящий извне от пчелы-хозяина, такой как ДНК, полученной от клеща, произвели революцию в диагностике, обеспечив специфическое, быстрое и высокопроизводительное обнаружение. Молекулярные методы с использованием экзогенной ДНК основаны на идентификации уникальных геном- ных областей паразитов с помощью полимеразной цепной реакции (ПЦР), количественной ПЦР (кПЦР) или анализе ДНК окружающей среды, что обеспечивает непревзойденную точность даже при низких уровнях заражения [14, 15]. Для V. destructor молекулярные анализы часто нацелены на маркеры митохондриальной ДНК: гены цитохромоксидазы I (COI) или рибосомальную РНК. Исследования Evans et al. (2003) показали, что праймеры ПЦР, специфичные для последовательностей COI V. destructor, могут обнаруживать одного клеща в объединенных образцах пчел, что превосходит чувствительность традиционных методов [11]. Аналогичным образом анализы кПЦР количественно определяют нагрузку клещей путем амплификации ядерных генов, таких как EF1-α, что позволяет пчеловодам отслеживать динамику заражения и оценивать эффективность лечения. Этот подход особенно ценен для обнаружения форети-ческих клещей, скрытых под склеритами пчел или в ячейках расплода, которые ускользают от визуального осмотра. Недавние достижения, такие как петлевая изотермическая амплификация (LAMP), позволяют проводить диагностику в полевых условиях с использованием портативных устройств, обходя необходимость в термоциклерах. Напротив, обнаружение A. woodi исторически основывалось на микроскопическом исследовании трахей пчел, методе, требующем специализированных знаний и склонном к ложноотрицательным результатам. Молекулярные инструменты обходят эти ограничения, ампли-фицируя видоспецифическую ДНК из тканей пчел или остатков улья. Sammatoro et al. (2012) разработали ПЦР-анализ, нацеленный на ген рРНК A. woodi 18S, способный идентифицировать клещей у пчел с ранними стадиями заражения [16]. Этот метод также отличает A. woodi от родственных видов, таких как A. dorsalis, что снижает риск неправильной диагностики. Кроме того, метабаркодирование eDNA – секвенирование ДНК, извлеченной из субстратов ульев, таких как воск, пыльца или мертвые пчелы, обеспечивает неинвазивную стратегию наблюдения. Фильтруя образцы окружающей среды и амплифицируя ДНК клещей, исследователи могут одновременно проверять несколько колоний, как было продемонстрировано в исследовании 2021 г., которое обнаружило A. woodi в 30 % бессимптомных ульев [17]. Хотя в последние десятилетия A. woodi затмевает V. destructor со своими более разрушительными последствиями для колонии медоносных пчел, A. woodi остается проблемой для российского пчеловодства, особенно в регионах с особыми климатическими и пчеловодческими условиями. Заражения клещами, в том числе вызываемые видами V. destructor и A. woodi, являются яркими примерами угроз, с которыми сталкиваются колонии медоносных пчел. По этой причине текущие передовые исследования распространения эктопаразитов, методов их индикации и идентификации, в том числе молекулярно-генетические исследования, имеют важное значение для самих хозяев - медоносных пчел. Эффективным методом идентификации эктопаразитов, которые не сразу заметны, но ранее контактировали с ульем и самими медоносными пчелами, является поиск и использование следов организмов в ульевом мусоре при помощи эДНК. Раннее обнаружение и отслеживание распространения этих двух эктопаразитов медоносных пчел будет эффективнее с применением данного подхода [18].
Цель исследования - получение генетической информации о циркулирующих гаплотипах A. woodi и V. destructor с использованием экзогенной ДНК на отдельных пасеках Республики Татарстан.
Задачи: определение уровня заражения V. destructor и A. woodi на отдельных пасеках в районах Республики Татарстан с определением циркулирующих гаплотипов и обнаружение следов заражения в колониях медоносных пчел с использованием экзогенной ДНК ульевого мусора.
Объекты и методы
Образцы медоносных пчел. Пчелы подвидов A. m. mellifera и A. m. carnica были собраны в 13 районах Республики Татарстан в период с марта по июнь 2024 г.
Всего было собрано 7 800 медоносных пчел с 26 частных пасек, расположенных в 13 районах Республики Татарстан, Россия. Процесс сбора образцов был приведен ранее [19]. Для сбора образцов в этом исследовании были выбраны следующие районы: Азнакаевский, Альметьевский, Апастовский, Буинский, Верхнеус-лонский, Высокогорский, Елабужский, Зеленодольский, Камско-Устьинский, Лаишевский, Мензелинский, Муслюмовский и Сабинский. Перед выделением ДНК образцы медоносных пчел хранились при температуре -30 °C.
Метод измерения количества пчел в улье. Количество рабочих пчел в улье медо- носных пчел оценивалось с использованием следующей информации: около трети рабочих пчел в улье каждый день собирают нектар и пыльцу. На основе среднего количества полетов в день одной пчелы и времени, потраченного на поиск нектара и пыльцы, можно использовать следующую формулу для расчета количества пчел в улье
N = 3 • (f/0.0138), где N - количество пчел в улье; f - количество пчел, покидающих гнездо за одну минуту (переменная f округлялась до ближайшего целого числа); значение 0.0138 - среднее количество времени, затрачиваемого на поиски нектара и пыльцы, для средней колонии медоносных пчел в средний день.
Метод оценки общей численности популяции клещей в колонии. Образец из примерно 300 живых пчел собирался из 13 районов (2 пасеки в каждом районе) в пластиковый контейнер. Замороженные образцы пчел в дальнейшем использовались для определения нагрузки клещами Varroa destructor и Acarapis woodi [20]. Доля зараженных клещами пчел (из 300) использовалась для расчета уровня заражения всего улья [21]. Измерение количества пчел в улье проводилось в то же время, что и отбор проб для определения уровня заражения клещами, чтобы точно рассчитать общий уровень заражения колонии [22]. Общее количество пчел в колонии было умножено на долю зараженных рабочих в исследуемом образце, чтобы получить размер популяции клеща Varroa destructor и Acarapis woodi в форетической фазе (во время которой клещ находится на/в теле пчелы).
Модифицированный метод оценки общей численности естественно погибших клещей Varroa destructor. Этот метод основан на количественной оценке естественно погибших и упавших вниз улья клещей. Несмотря на то что различные исследования делают противоречивые выводы относительно точности метода оценки естественного погибших клещей для определения общей степени заражения, поскольку количество упавших вниз улья клещей во многом определяется количеством появляющегося зараженного расплода [23], в целом данный метод считается хорошим индикатором заражения колонии [24]. Для оценки общей численности естественного погибших клещей нижний пластиковый поддон вытаскивался из улья и клещи на нем подсчитывались. Мертвые пчелы на по- верхности поддона также проверялись, поскольку в случае падения живого клеща вниз улья мертвые пчелы могут притянусь к себе паразита. Сбор данных проводился в течение 2 недель. Полученный показатель был усреднен для получения среднего недельного количество упавших клещей. Этот период охватывает естественные колебания падения клещей, связанные с циклами динамики популяции в пределах хозяина. На основе среднего недельного количества упавших клещей был рассчитан общий уровень заражения колонии, а именно: число ежедневно падавших клещей было умножено на поправочный коэффициент (250).
Индикация и идентификация клещей Var-roa destructor и Acarapis woodi с использованием экзогенной ДНК. Перед экстракцией ДНК с каждого пластикового поддона со дна улья считали весь ульевой мусор и измельчали ультразвуком в 5 мл вновь добавленной mQ воды. Экстракцию ДНК из образцов ульевого мусора проводили по ранее описанному протоколу [15]. Пробирки встряхивали в течение 1 мин и инкубировали при 40 °C в течение 30 мин. Затем пробирки центрифугировали при комнатной температуре в течение 25 мин при 5000× g с дальнейшим удалением супернатанта. После ресуспендирования осадка в 5 мл mQ воды содержимое 4 пробирок объединяли в одну и еще раз разбавляли mQ водой. Осадок ресуспендировали в 0,5 мл mQ воды после центрифугирования в течение 25 мин при 5000× g при комнатной температуре с удалением супернатанта. Затем в каждый образец добавляли 1 мл CTAB буфера (2 % (w/v) цетилтриметиламмоний бромида; 1,4 M NaCl; 100 мМ Трис-HCl; 20 мМ ЭДТА; pH 8) и 5 мкл раствора РНКазы А (10 мг/мл). Полученную смесь инкубировали в течение 10 мин при 60 °C. Далее добавляли 30 мкл протеиназы К (20 мг/мл), перемешивали и инкубировали 90 мин при 65 °C. Далее образцы охлаждали до комнатной температуры и центрифугировали при 16 000× g в течение 10 мин. После центрифугирования 700 мкл супернатанта переносили в пробирку с 500 мкл хлороформа/изоамилового спирта (24 : 1) и встряхивали. Далее центрифугировали при 16 000×g 15 мин при комнатной температуре. После переноса супернатанта в чистую пробирку объемом 1,5 мл для осаждения ДНК использовали 500 мкл изопропанола, а для промывки – 500 мкл 70 % этанола. Осадки хранили при –20 °C до проведения ПЦР-анализа и до начала использования регидратировали
30 мкл mQ воды. Выделенную ДНК визуально оценивали с помощью электрофореза в 1% агарозном геле в буфере TBE 1X, а качество подтверждали с помощью нанофотометра. Для ПЦР-анализа использовали сконструированные в Primer BLAST Acarapis woodi -специфичные праймеры F_CTCCAGCTGTAACAAGGGTTGA и R_ATT-GGGTGGGATGGCTTAGG для амплификации мтДНК гена cox1 размером 322 п.н. и Varroa destructor -специфичные праймеры F_GGAGTA-GGTACAGGTTGAACGG и R_AGGCTGCTTCTT-CCCTCTTTG для амплификации мтДНК гена cox1 размером 441 п.н.
Построение филогенетического дерева. Филогенетическое дерево было построено с использованием метода ближайших соседей (Neighbor-Joining method) [25]. На рисунке 1 показано оптимальное дерево. Для расчета процент реплик деревьев, в которых связанные таксоны подверглись кластеризации, использовался bootstrap тест (1000 реплик); процент реплик деревьев показан под ветвями. Дерево нарисовано в масштабе, с длинами ветвей в тех же единицах, что и эволюционные расстояния, используемые для выведения филогенетического дерева. Эволюционная дистанция была вычислена с использованием метода Tajima-Nei [25] и находится в единицах числа замен оснований на сайт. Различия в смещении состава между последовательностями рассмотрены с использованием модели замен. В анализ вошли 103 нуклеотидные последовательности. Все участки последовательностей, содержащие пробелы и отсутствующие данные, были устранены (опция полного удаления). Всего в конечном наборе данных было 719 позиций. Эволюционный анализ был проведен в MEGA11 [26].
Статистический анализ. Для интерпретации полученных данных были рассмотрены зависимости между полученными переменными. Переменным были присвоены названия: BS (подвид пчелы A. m. carnica и A. m. mellifera ), AW (визуальное обнаружение клеща A. woodi ), VD (визуальное обнаружение клеща V. destructor ), exAW (обнаружение клеща A. woodi в экзогенной ДНК), и exVD (обнаружение клеща V. destructor в экзогенной ДНК). Корреляционный анализ проводили с использованием теста Пирсона, линейная регрессия проводилась с использованием статистического программного обеспечения для MS Excel.
Результаты и их обсуждение. Клещи A. woo-di были обнаружены в трахеях собранных медо- носных пчел A. m. carnica в Камско-Устьинском районе. Численность популяции трахейного клеща A. woodi в колонии пчелы составила 235 особей на улей из 8 697 пчел (2,7 %, 95 % доверительный интервал: 2,37–3,07 %). Клещи A. woodi были обнаружены в трахеях, а V. destructor – на брюшке сбоку и на груди собранных медоносных пчел A. m. mellifera в Лаишевском районе. Численность популяции трахейного клеща A. woodi в колонии пчелы составила 869 особей на улей из 4 347 пчел (19,99 %, 95 % доверительный интервал: 18,81–21,21 %). Численность популяции V. destructor в колонии пчелы составила 250 особей на улей из 4 347 пчел (5,75 %, 95 % доверительный интервал: 5.08–6.49 %). Клещи A. woodi
были обнаружены в трахеях собранных медоносных пчел A. m. mellifera в Высокогорском районе. Численность популяции трахейного клеща A. woodi в колонии пчелы составила 557 особей на улей из 5 217 пчел (10,67 %, 95 % доверительный интервал: 9,85–11,55 %) (табл. 1). В Альметьевском, Апастовском, Азнакаевском, Буинском, Мензелинском, Муслюмовском, Сабинском, Верхнеуслонском и Зеленодольском, а также в Апастовском, Буинском, Елабужском, Камско-Устьинском, Лаишевском, Муслюмов-ском, Сабинском, Верхнеуслонском, Высокогорском и Зеленодольском районах A. woodi и V. destructor обнаружено не было.
Таблица 1
Оценка общей численности популяции клещей Acarapis woodi и Varroa destructor в колониях медоносной пчелы Apis mellifera на обследованных пасеках Республики Татарстан и обнаружения экзогенной ДНК эктопаразитов
Estimation of the total population size of Acarapis woodi and Varroa destructor mites in colonies of the honey bee Apis mellifera in surveyed apiaries of the Republic of Tatarstan and detection of exogenous DNA of ectoparasites
|
Район |
Номер пасеки в статье |
Общее кол-во взрослых рабочих пчел в улье |
Численность A. woodi в колонии |
Обнаружение A. woodi в эДНК |
Численность V. destructor в колонии |
Обнаружение V. destructor в эДНК |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Альметьевский |
1 |
4565 |
– |
– |
– |
– |
|
2 |
7826 |
– |
+ |
– |
– |
|
|
Апастовский |
3 |
5652 |
– |
– |
– |
– |
|
4 |
4782 |
– |
– |
– |
– |
|
|
Азнакаевский |
5 |
7609 |
– |
– |
– |
– |
|
6 |
12826 |
– |
– |
– |
– |
|
|
Буинский |
7 |
6304 |
– |
– |
– |
– |
|
8 |
8260 |
– |
– |
– |
– |
|
|
Елабужский |
9 |
10217 |
– |
– |
– |
+ |
|
10 |
6304 |
– |
– |
– |
– |
|
|
Камско- Устьинский |
11 |
8697 |
235 |
+ |
– |
– |
|
12 |
6956 |
– |
– |
– |
– |
|
|
Лаишевский |
13 |
4347 |
869 |
+ |
250 |
+ |
|
14 |
8913 |
– |
– |
– |
– |
|
|
Мензелинский |
15 |
8478 |
– |
– |
– |
– |
|
16 |
8260 |
– |
+ |
– |
– |
|
|
Муслюмовский |
17 |
7609 |
– |
– |
– |
– |
|
18 |
6956 |
– |
– |
– |
– |
|
|
Сабинский |
19 |
6304 |
– |
– |
– |
– |
|
20 |
7173 |
– |
– |
– |
– |
|
|
Верхнеуслонский |
21 |
6304 |
– |
– |
– |
– |
|
22 |
7826 |
– |
– |
– |
– |
Окончание табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Высокогорский |
23 |
6957 |
557 |
+ |
– |
+ |
|
24 |
5217 |
– |
– |
– |
– |
|
|
Зеленодольский |
25 |
8478 |
– |
– |
– |
– |
|
26 |
7391 |
– |
– |
– |
– |
Таблица 2
Результаты линейного регрессионного анализа на наличие связи между подвидом пчелы Apis mellifera , заклещеванностью Acarapis woodi и Varroa destructor на обследованных пасеках Республики Татарстан, а также обнаружением экзогенной ДНК эктопаразитов Linear regression analysis for the relationship between Apis mellifera subspecies, Acarapis woodi and Varroa destructor infestation in the surveyed apiaries in the Republic of Tatarstan, and ectoparasite exogenous DNA detection
|
Переменная |
R2 |
Стандартная ошибка |
Уровень значимости p |
|
AW |
–0,712 |
0,136 |
< 0,05 (достоверно) |
|
VD |
0,132 |
0,182 |
> 0,05 |
|
AW |
–1,61659 |
0,22559 |
< 0,05 (достоверно) |
|
VD |
1,44795 |
0,32881 |
< 0,05 (достоверно) |
A. woodi склонен распространяться горизонтально из зараженной колонии в свободную колонию. Это может быть причиной того, что A. woodi так локально распространен на пасеках в данном исследовании. Известно, что взрослые пчелы в большей степени склонны быть ослабленными и/или иметь деформированные крылья во время заражения V. destructor (и в меньшей степени это выражено при заражении A. woodi ); при этом пчелы не могут вылетать из ульев и собирать нектар и пыльцу. В данном исследовании медоносные пчелы A. mellifera собирали нектар и пыльцу и визуально не были ослабленными, также не имели деформированных крыльев, что может быть одним из подтверждений редкого обнаружения клещей V. destructor у A. mellifera на пасеках. Другой причиной малого обнаружения двух видов клеща было применение флавулината (со слов пчеловодов). Однако, даже несмотря на то, что в большинстве случаев на пасеках применялся
флавулинат, а также учитывая тот факт, что в период исследования основная масса клещей V. destructor находится в запечатанном расплоде и определение степени поражения взрослых пчел клещом Varroa не может служить объективным показателем поражения и клещей обнаружить не удалось, была обнаружена экзогенная ДНК A. woodi в Альметьевском и Мензелинском районах и экзогенная ДНК V. destructor в Елабужском и Высокогорском районах. По результатам исследования было выявлено присутствие клещей у двух подвидов медоносной пчелы: A. m. carnica и A. m. mellifera . С использованием переменных BS, AW, VD, exAW и exVD было решено построить корреляционную матрицу Пирсона. По ее результатам не было выявлено корреляции между присутствием клеща A. woodi (r = –0,085) и V. destructor (r = 0,17) на пасеках ни у одного из подвидов медоносной пчелы (рис. 1).
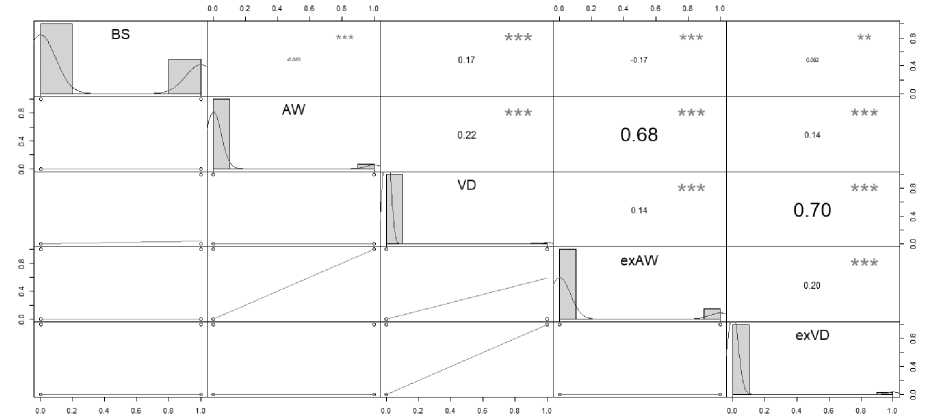
Рис. 1. Корреляционная матрица Пирсона с использованием данных визуального обнаружения клещей A. woodi и V. desructor и обнаружения экpогенной ДНК клещей в ульевом мусоре и данных подвида медоносных пчел с заклещеванных пасек Республики Татарстан
Pearson correlation matrix using visual detection data of A. woodi and V. desructor mites and detection of exogenic DNA of mites in hive debris and honey bee subspecies data from mite-infested apiaries of the Republic of Tatarstan
Выявление экзогенной ДНК высоко коррелировало с визуальным обнаружением клещей A. woodi (r = 0,68) и V. destructor (r = 0,7) на пасеках. Линейная регрессия выявила достоверную связь между AW (визуальным обнаруже-
нием клеща A. woodi ) и подвидом пчелы (табл. 2). Кроме того, обнаружение как клеща A. woodi, так и V. destructor в экзогенной ДНК (exAW и exVD) в обоих случаях было связано с подвидом A. m. mellifera .
Слабая положительная корреляция между за-клещеванностью A. woodi и V. destructor (r = 0,22) (табл. 2) и достоверная связь между наличием экзогенной ДНК A. woodi и V. destructor и подвидом A. m. mellifera могут предполагать взаимное усиление восприимчивости хозяина к заражению одним из видов эктопаразитов. Несмотря на то что в 1964 г. в СССР выявили V. destructor и сопутствующее заболевание варроатоз пчел, считается, что случаи акарапидоза сократились в связи с широким применением различных акари-
IT 11«wM тт1
MW748233
61 MW748228
MW7«229
. MW748231
I---------------- MW748232
\_______________.омваааш
I------------------- LC512730
цидных средств [27]. Вполне возможно, что в условиях применения акарицидов на исследованных пасеках (со слов пчеловодов) эктопаразиты в ульях все равно сохраняются. В связи с этим анализ гаплотипов эктопаразитов особо важен. Экзогенная ДНК A. woodi c обследованных пасек в этом исследовании показала, что клещи имели один гаплотип, идентичный изолированным в других странах (Турция, Япония) – гаплотип «RT-12nov24-tm1» и один уникальный гаплотип – «RT-12dec24-tm15» (рис. 2).
------------- RT 12duc24-«Tl5
---------- MW74823Q
Митохондриальные последовательности cox1 A. woodi в Республике Татарстан, которые полностью идентичны изолятам из Турции, указывают на генетическую близость изолятов. Кроме того, результаты филогенетического анализа показали, что существующие изоляты также тесно связаны с A. woodi, обнаруженным у A. cerana japonica в Японии (LC512730). Судя по тому, что оба подвида медоносной пчелы в данном исследовании были заражены клещом (A. m. mellifera и
A. m. carnica ), принадлежность к какому-либо подвиду в пределах вида A. mellifera не является барьером для распространения A. woodi. Однако в данном исследовании A. m. mellifera была заражена чаще, что статистически подтверждается результатами линейно-регрессионного анализа ( p < 0,05). Существует вероятность наличия выгодных взаимодействий с популяцией конкретного подвида хозяина A. m. mellifera , которые выгоднее для конкретного гаплотипа эктопаразита A. woodi . Подтверждением этому может служить наличие уникального гаплотипа «RT-12dec24-tm15». Остается вопрос об уникальности данного гаплотипа для территории пасеки в пределах Республики Татарстан.
Экзогенная ДНК V. destructor c обследованных пасек в Республике Татарстан в этом исследовании показала, что клещи имели почти идентичный гаплотип («RT-12oct24-t3») с изоля-тами, собранными из Сербии, Турции, Аргентины, Кореи, Вьетнама и Новой Зеландии (рис. 3). Тем не менее наблюдаемый гаплотип отличался на один нуклеотид, что достаточно, чтобы предположить генетическую дифференциацию. Доминирующий корейский гаплотип (штамм K), выявленный у российских клещей с помощью исследований митохондриальной ДНК, в зарубежных исследованиях демонстрирует высокую вирулентность и адаптивность. Однако обнаруженный в данном исследовании гаплотип потенциально может отличаться от ранее зафиксированного корейского штамма К. Присутствие паразитов V. destructor с генетическими различиями указывает на наличие взаимодействий с популяцией хозяина, которые управляют эволюцией V. destructor.
MW725321 HTJ
OK560Q14Asrtnncn NZ 0*560015 Pon rua NZ MW725315 PH
0*6262 57 Kdtwn Wdlingtan NZ 0*6262 58 AsHuricnd NZ 0*626254 Harais Hamilton NZ 0*626255 Endo-icy Hamilton NZ
0*6262 59 AsTtor torr9 NZ
*4 528381 П1 1
RT 12oct24 13
MW599115TWTNm649
MW599108TW KNm 675
MW599111TWTNm 643
M WS99113 TW TN m 638
MW5*9117TWTNm 851
MW599107TW KN m 674
30379389 Ат J1 - ITawan Tadtong
*4 528382 □ 1
MW599142 VN TLMD c 633
MW599144 VN TLMD c 635
MW599123 VN BWH2 c 613
MW599125VN BWH2c615
MW599137VN KSMH3c630
*R528378AI 1
GO379065A£1 1Chr»ZhJia
30379384 AcV1 4Thaland QangChangJay
MW599154 VN TTTH2 c 619
MW599124VN BWH2c614
MWSB9120W BWH1 c610
MW599150 VN ТОРТ c 638
MW599156 VN TTTH2 c 621
MW599133VN KSMH1 c627
MW599136 VN KSM H2 c 668
MW599131 VN KSMH1 c625
MW599152 VN TTTH1 c617
Заключение. Несмотря на то что паразитические клещи V. destructor в целом чаще встречаются на пасеках, чем A. woodi , у обоих видов существуют наиболее «вредоносные» гаплотипы. Исходя из обнаруженных гаплотипов у обоих видов, особенно уникальных, требуется дальнейший анализ на переносимые патогены, так как данное свойство придает гаплотипам их «вредоносность». Связь подвидов медоносной пчелы с паразитическими клещами требует дальнейшей оценки с использованием большей выборки, молекулярной идентификацией подвидов пчелы с оценкой метизации, гигиенического поведения, так как, например, исследования A. m. carnica предполагают, что у подвида повышается процент заклещеванности при гибридизации с A. m. mellifera , но снижается при гибридизации с A. m. iberiensis [27–31]. Кроме того, в дальнейшем необходимо комплексно оценить объективность использования методик расчета клещевой нагрузки в колониях медоносной пчелы на территории исследования.
На отдельных пасеках в районах Республики Татарстан было обнаружено: 1) на трех пасеках в трех районах зафиксирован низкий уровень заражения A. woodi ; обнаружены два гаплотипа: один уникальный и один общий, идентичный изолированным по всему миру; 2) на одной пасеке в одном районе зафиксирован низкий уровень заражения клещом V. destructor ; обнаружен 1 гаплотип, схожий с изолированными по всему миру; 3) несмотря на то что в период исследования основная масса клещей V. destructor находится в запечатанном расплоде и определение степени поражения взрослых пчел клещом Varroa не может служить объективным показателем поражения, экзогенная ДНК ульевого мусора позволила обнаружить следы заражения клещами в колониях медоносных пчел в целом на шести пасеках в шести районах; 4) обнаружение экзогенной ДНК A. woodi и V. destructor было статистически достоверно связано с подвидом A. m. mellifera в исследованных колониях.
