Идентификация сортов сои (Glycine max L.) с использованием микросателлитных локусов ДНК
 с использованием микросателлитных локусов ДНК")
Автор: Рамазанова С.А.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (166), 2016 года.
Бесплатный доступ
Был изучен внутрисортовой полиморфизм сортов сои. Выявлено, что два сорта Лада и Вилана являются генетически неоднородными. В работе было проанализировано 23 сорта сои селекции ВНИИМК по девяти известным микросателлитным локусам (SSR). Результаты амплификации ДНК показали, что из девяти изученных SSR-локусов восемь оказались полиаллельными, а один - Soygy2 - мономорфным. Среднее число аллелей на локус составило 3,2, у локуса Satt9 было выявлено максимальное количество аллелей -пять. С использованием системы из девяти SSR-маркеров были идентифицированы сорта сои, составлены их молекулярно-генетические паспорта и установлена уникальность каждого сорта.
Соя, пцр, ssr-локусы, днк, идентификация
Короткий адрес: https://sciup.org/142151307
IDR: 142151307 | УДК: 633.853.52:34.23.37
Identification of soybean (Glycine max L.) cultivars using microsatellite DNA loci
Intravarietal polymorphism of soybean cultivars was studied. It was resulted two cultivars Lada and Vilana are heterogeneous. Totally 23 soybean cultivars of VNIIMK’s breeding were analyzed on nine known microsatellite loci (SSR). DNA amplification resulted that eight from nine studied loci appeared to be polyallelic, and one locus - Soygy 2 is monomorphic. An average number of alleles per locus was 3.2; locus Satt 9 was revealed to have maximal allele numbers -five. Using system of nine SSR-markers, soybean cultivars were identified, their molecular-genetic passports were done and uniqueness of each cultivar was proved.
Текст научной статьи Идентификация сортов сои (Glycine max L.) с использованием микросателлитных локусов ДНК
Введение. Важным элементом при разработке современных селекционных программ улучшения культурной сои является установление особенностей ее генетического полиморфизма. Соя, по сравнению со многими видами сельскохозяйственных растений, имеет относительно низкий уровень генетической вариабельности. Разработка подходов к дифференциации и идентификации сортов этой культуры актуальна для генетико-селекционных исследований и защиты авторских прав.
Наиболее удобными для идентификации генотипов в настоящее время являются молекулярно-генетические маркеры, т.е. маркеры на основе макромолекул: запасные белки, изоферменты и полиморфные фрагменты ДНК. Они в меньшей мере подвержены фенотипической изменчивости, чем морфологические маркеры, и в большинстве случаев имеют кодоминантное наследование. На их основе проводится биохимическая паспортизация многих сельскохозяйственных культур.
По данным многих авторов, высокий уровень полиморфизма у сои удалось выявить только по микросателлитным локусам [1; 2; 3; 4]. Микросателлиты (SSRs) представляют собой простые, наиболее доступные, удобные и относительно недорогие маркеры, пригодные для иденти- фикации генотипов [6; 7]. Они позволяют дифференцировать особи, отличающиеся друг от друга небольшим количеством генов, что не всегда заметно фенотипически.
Целью этой работы было изучение молекулярно-генетического полиморфизма генома культурной сои ( Glycine max L.) с использованием ПЦР-анализа микроса-теллитных локусов ДНК и создание на этой основе системы маркеров, пригодной для дифференциации, идентификации и паспортизации сортов сои селекции ВНИИМК.
Материалы и методы. В работе использованы 23 сорта сои ( G. max ) селекции ВНИИМК. Для исследования были выбраны девять пар SSR-праймеров: Satt1, Satt2, Satt5, Satt9, Soypr1, Sat1, Sat36, Soygy2, Soyhsp176.
ДНК выделяли из фрагментов зеленых листьев 5–10 растений. Выделение проводили по модифицированному методу [8].
Полимеразную цепную реакцию осуществляли в объеме реакционной смеси 25 мкл, содержащем: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 1,5–3 мM MgCl 2 ; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 еди-ницу рекомбинантной термостабильной ДНК полимеразы (Москва, ГОСНИИГЕНЕТИКА). Амплификацию проводили в приборе Терцик (ДНК-технология, Россия). Подробно температурные режимы для каждого праймера опубликованы ранее [9]. Концентрацию ДНК определяли по интенсивности окрашивания ее бромистым этидием в 1 %-ном агарозном геле.
Электрофорез продуктов амплификации проводили в агарозном геле (2 % агароза, 1х ТАЕ-буфер). Документирование результатов электрофореза обеспечивалось при помощи системы цифровой документации видеоизображения BIO-PRINT (Vilber Lourmat, Франция).
Результаты и обсуждение. Первым этапом работы было выявить внутрисор-товой полиморфизм сортов сои селекции ВНИИМК по изученным микросателлит-ным локусам. Поскольку соя является самоопыляющейся культурой, предполагается, что культивируемые сорта генетически однородны. Однако бывают исключения, поэтому для паспортизации сорта по микросателлитным маркерам нами была выделена ДНК с десяти растений двух сортов: Фора и Лада (из каждого проростка ДНК выделялась отдельно), и проведен ее анализ по девяти микроса-теллитным локусам.
Сорт Фора – выровненный сорт, и анализ состояния микросателлитных локусов подтвердил это. На рисунке 1 представлены фореграммы продуктов амплификации ДНК сорта сои Фора по четырём локусам. Каждая дорожка – это амплифицированный фрагмент ДНК отдельного растения. Как видно, все фракции имеют одинаковую длину пар нуклеотидов, т.е. различий в размерах фракций нет. Следовательно, полиморфизм по данным локусам внутри сорта отсутствует. Анализ был проведен по всем девяти локусам. На остальных фо-реграммах различий также нет. Наши данные подтверждают, что у сорта сои Фора отсутствует внутрисортовой полиморфизм.
Совсем другая картина наблюдается у сорта Лада. По локусу Satt5 у трех растений фракции ДНК большего размера – дорожки 5, 6 и 8 (рис. 2А). По локусу Satt2 из 10 фракций ДНК каждого отдельного растения выделяется фрагмент на шестой дорожке (рис. 2Б) он большего размера (по количеству пар нуклеотидов). По локусу Soyhsp176 выделяется фракция на второй дорожке (рис. 2В): она меньшей длины. По локусу Satt9 фрагмент на второй дорожке отличается от остальных большим размером (рис. 2Г). По осталь-
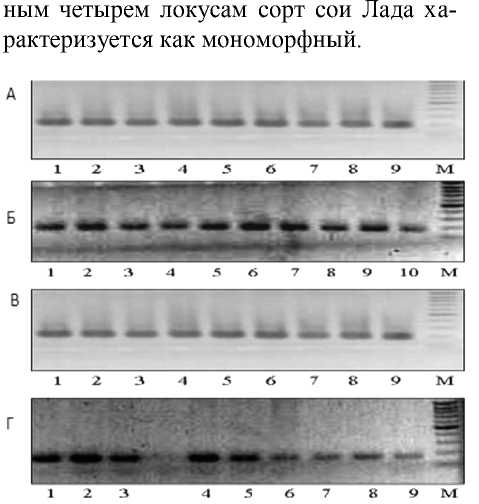
Рисунок 1 – Фореграмма продуктов амплификации ДНК сорта сои Фора по локусам А – Satt9, Б – Soypr1, В – Satt5,
Г – Satt 2. Дорожки 1–9 – ДНК отдельных растений, М – маркер молекулярного веса 100 bp DNA Ladder
Soyhsp176 – три и у трех локусов – Satt 2, Soypr1, Soygy2 и Sat36 – по два аллеля.
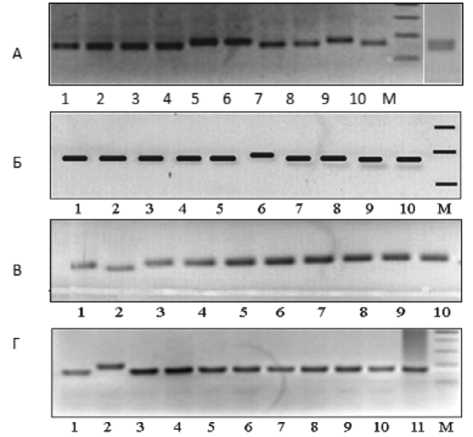
Рисунок 2 – Фореграммы продуктов амплификации ДНК сорта сои Лада по локусу А – Satt5, Б – Satt2, B – Sohsp176, Г – Satt9 . Дорожки 1–10 – ДНК отдельных растений, 11 – смесь ДНК, М – маркер молекулярного веса 100 bp DNA Ladder
Таким образом, из девяти микросател-литных локусов по четырем этот сорт сои оказался неоднороден. Это свидетельствует о внутрисортовом полиморфизме по изученным микросателлитным локусам.
Из изученных нами сортов внутрисор-товой полиморфизм микросателлитных локусов выявлен и у сорта Вилана. Остальные изученные сорта однородны. Таким образом, при создании молекулярногенетического паспорта сорта сои требуется учитывать внутрисортовой полиморфизм.
Результаты амплификации ДНК двадцати трех сортов сои показали, что из девяти изученных SSR-локусов восемь полиаллельны и только локус Soygy2 оказался мономорфен. Всего в этой группе генотипов было выявлено 29 аллелей. Число аллелей на локус варьировало от 1 до 5, что в среднем составило 3,2. У локуса Satt9 было выявлено максимальное количество аллелей – пять, у трех локусов – Satt1, Satt5 и Sat1 – по четыре, у локуса
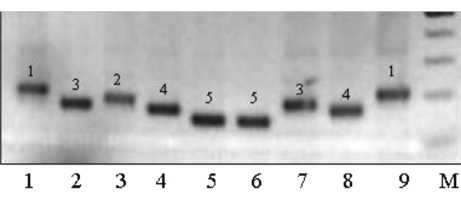
Рисунок 3 – Фореграмма продуктов амплификации ДНК сортов сои по локусу Satt9. Дорожки 1–9 – фрагменты ДНК сортов, М – маркер молекулярного веса 100 bp DNA Ladder
Аллели SSR-локусов представлены на фореграммах фрагментами ДНК разного молекулярного веса (длины пар нуклеотидов). На рисунке 3 показаны фореграм-мы результатов амплификации ДНК некоторых сортов по локусу Satt9, выявившему наибольшее количество аллелей 5. Нумерацию их по каждому локусу проводили следующим образом: фраг- мент ДНК с максимальным значением молекулярного веса обозначали цифрой 1 (дорожки 1, 9) и далее по мере уменьшения молекулярного веса цифрами 2 (дорожка 3), 3 (дорожки 2, 7), 4 (дорожки 4, 8) и 5 (дорожки 5, 6).
Для всех изученных сортов сои селекции ВНИИМК получены уникальные наборы аллелей. Различия наблюдались по одному и более локусам. К примеру, сорта Лань, Валента, Рента, РВБ и Лакта, отличаются друг от друга по одному локусу Satt9. У остальных сортов есть отличия по двум и более локусам. Линии Д-6 и Д-4 являются сестринскими и поэтому очень похожи, но все же их можно различить по локусу Sat1 (таблица).
Таблица
Формулы генотипов сои селекции ВНИИМК
|
Сорт |
Формула ∗ |
|
Лань (ВНИИМК) |
А 2 B 2 C 2 D 1 E 1 F 1 G 2 H 3 I 2 |
|
Лира (-//-) |
A 1 B 1 C 1 D 3 E 2 F 1 G 2 H 2 I 2 |
|
Фора (-//-) |
A 2 B 1 C 2 D 2 E 1 F 4 G 1 H 3 I 2 |
|
Валента (-//-) |
A 3 B 2 C 2 D 4 E 1 F 2 G 2 H 3 I 2 |
|
Дельта (-//-) |
A 3 B 2 C 2 D 5 E 1 F 1 G 2 H 3 I 2 |
|
Парма (-//-) |
A 3 B 1 C 1 D 4 E 2 F 2 G 2 H 2 I 2 |
|
Рента (-//-) |
A 3 B 2 C 2 D 2 E 1 F 2 G 2 H 3 I 2 |
|
РВБ (-//-) |
A 3 B 2 C 2 D 3 E 1 F 2 G 2 H 3 I 2 |
|
РВФ (-//-) |
A 2 B 2 C 2 D 2 E 1 F 2 G 2 H 3 I 2 |
|
Веста (-//-) |
A 2 B 1 C 3 D 3 E 1 F 1 G 2 H 3 I 2 |
|
Вилана (-//-) |
A 3 A 4 B 2 C 2 D 1 E 1 F 2 G 2 H 3 I 2 |
|
Лакта (-//-) |
A 2 B 2 C 2 D 2 E 1 F 1 G 2 H 3 I 2 |
|
Ника (-//-) |
A 2 B 2 C 2 D 1 E 2 F 1 G 2 H 3 I 2 |
|
Дива (-//-) |
A 2 B 2 C 1 D 5 E 1 F 1 G 2 H 3 I 2 |
|
Диана (-//-) |
A 2 B 1 C 2 D 2 E 3 F 2 G 2 H 3 I 2 |
|
Памела (-//-) |
A 3 B 1 C 2 D 3 E 1 F 4 G 2 H 3 I 2 |
|
Д-6 (-//-) |
A 1 B 2 C 1 D 3 E 2 F 1 G 2 H 2 I 2 |
|
Д-4 (-//-) |
A 1 B 2 C 1 D 3 E 2 F 2 G 2 H 2 I 2 |
|
Альба (-//-) |
A 2 B 2 C 2 D 3 E 2 F 4 G 2 H 3 I 2 |
|
Трембита (-//-) |
A 3 B 2 C 4 D 3 E 2 F 3 G 2 H 2 I 2 |
|
Лиана (-//-) |
A 3 B 2 C 4 D 2 E 2 F 2 G 2 H 2 I 2 |
|
Лада (-//-) |
A 3 B 2 C 3 C 2 D 4 E 2 F 2 G 2 H 2 I 2 |
|
Б-2 (-//-) |
A 2 B 2 C 3 D 1 E 1 F 1 G 2 H 3 I 2 |
∗ Примечание: код локуса A – Satt1; B – Satt2;
C – Satt5; D – Satt9; E – Soypr1; F – Sat1; G – Sat36;
H – Soyhsp176, I – Soygy2
Для каждого сорта на основании полученного набора аллелей микросателлит-ных локусов были составлены молекулярно-генетические паспорта или, так на-66
зываемые, генетические формулы генотипов. Большими буквами латинского алфавита был обозначен код локуса, а нижний индекс – аллельное состояние данного локуса. В качестве примера в таблице представлены молекулярногенетические формулы изученных сортов сои.
Выводы. В результате проведенной работы было установлено, что дискриминационный потенциал изученной маркерной системы достаточно высокий для того, чтобы использовать ее для идентификации и паспортизации сортов культурной сои. Изучение внутрисортового полиморфизма микросателлитных локусов ДНК выявило генетическую неоднородность сортов сои Лада и Вилана. С использованием системы из девяти SSR-маркеров были идентифицированы 23 сорта селекции ВНИИМК, для каждого из них получены уникальные наборы аллелей. На основании данных об аллельном разнообразии изученных микросателлит-ных локусов разработаны молекулярногенетические формулы, которые предложено использовать как соответствующие паспорта для идентификации и сертификации сортов сои.
Список литературы Идентификация сортов сои (Glycine max L.) с использованием микросателлитных локусов ДНК
- Rongwen J., Akkaya M.S., Bhagwat A.A. . The use of microsatellite DNA markers for soybean genotype identification//Theor. Appl. Genet. -1995. -V. 90. -P. 43-48.
- Maughan P., Saghai-Maroof M., Buss G. Microsatellite and amplified sequence length polymorphism in cultivated and wild soybean//Genom. -1995. -V. 38. -P. 715-723.
- Abe J., Xu D., Suzuki Y., Kanazawa A., Shimamoto Y. Soybean germplasm pools in Asia revealed by nuclear SSRs//Theor. Appl. Genet. -2003. -V. 106. -No 3. -P. 445-453.
- Diwan N., Cregan P.B. Automated sizing of fluorescent-labeled simple sequence repeat (SSR) markers to assay genetic variation in soybean//Theor. Appl. Genet. -1997. -V. 95. -No 5-6. -P. 723-733.
- Yamanaka N., Okabe A., Adachi T., Yang C. Characteristics and genetic diversity of soybean genetic resources in Northeast China//JIRCAS Research Highlights. -2003. -. -Режим доступа: http://www.jircas.affrc.go.jp/DB/guide-eng.html.
- Использование ПЦР-анализа в генетико-селекционных исследованиях: науч.-метод. рук-во/Под. ред. Сиволапа Ю.М. -Киев: Аграрна наука, 1998. -156 с.
- Сулимова Г.Е. ДНК-маркеры в генетических исследованиях: типы маркеров, их свойства и области применения//Успехи современной биологии. -2004. -Т. 124. -№ 3. -С. 260-271.
- Saghai-Maroof M.A., Soliman K.M., Jorgensen R.A., Allard Ribosomal R.W. DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics//PNAS USA. -1984. -81. -P. 8014-8018.
- Рамазанова С.А., Гучетль С.З., Челюстникова Т.А., Антонова Т.С. Полиморфизм микросателлитных локусов ДНК сортов сои селекции ВНИИМК//Сб. докл. межд. науч.-практ. конф. «Современные проблемы научного обеспечения производства подсолнечника», посвященной 120-летию со дня рождения академика В С. Пустовойта», ВНИИМК, Краснодар, Россия, 19-22 июля 2006. -С. 234-239.
