Использование бухты Улан-Хада на озере Байкал в голоцене (по фаунистическим материалам)
")
Автор: Номоконова Т.Ю., Горюнова О.И., Лозей Р.дЖ., Савельев Н.А.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Эпоха палеометалла
Статья в выпуске: 2 (46), 2011 года.
Бесплатный доступ
В статье изложены результаты изучения костных фаунистических остатков с многослойного поселения Улан-Хада - одного из опорных геоархеологических объектов Прибайкалья для реконструкций палеоэкологических и культурных изменений в голоцене. Дается полный анализ фаунистической коллекции, сформированной за все годы раскопок. Впервые исследуется ихтиофауна. Уточняется видовой состав млекопитающих, определенных ранее. Проводится сравнение видового разнообразия животных по хронологическим срезам от финального мезолита до позднего железного века. Установлено, что на стоянке в эпоху неолита и в бронзовом веке основным видом промысла была охота на нерпу и парнокопытных (косуля и благородный олень). Значительную роль играло рыболовство, особенно 4,2-3,8 тыс. л.н. (бронзовый век).
Фаунистические остатки, многослойное поселение, мезолит, неолит, бронзовый век, железный век, прибайкалье
Короткий адрес: https://sciup.org/14522855
IDR: 14522855 | УДК: 903
Текст научной статьи Использование бухты Улан-Хада на озере Байкал в голоцене (по фаунистическим материалам)
Поселение в бухте Улан-Хада на оз. Байкал – первый (и многие годы единственный) многослойный памятник эпохи голоцена, обнаруженный на территории Сибири [Петри, 1916]. Результаты междисциплинарных исследований, проводившихся на объекте, позволяют считать памятник опорным для создания периодизации и хронологии голоценовых культур Прибайкалья, для реконструкции природных и климатических изменений в регионе [Горюнова, 1984; Горюнова, Савельев, 1990].
В предлагаемой статье изложены результаты анализа костных остатков с поселения Улан-Хада, проведенного в рамках Байкальского археологического проекта (Университет Альберта, г. Эдмонтон, Канада,
Иркутский государственный университет, Россия). Впервые исследовались фаунистическая коллекция (ок. 3 тыс. костей), собранная за все годы раскопок (1974, 1979, 1982, 1990), и остатки рыб со стоянки. Определение видового состава животных выполнено Т.Ю. Номоконовой и Р.Дж. Лозей в 2009 г. Предварительные данные анализа некоторых костей млекопитающих со стоянки Улан-Хада (ок. 150 костей), определенных ранее А.А. Хамзиной [1991], были существенно дополнены и уточнены.
Методика зооархеологических определений и количественного анализа, используемая в статье, уже была подробно описана в ряде публикаций [Номоконова, Ло-зей, Горюнова, 2006; Nomokonova, Losey, Goriunova, 2009]. Подсчет фаунистических материалов производился по количеству определимых экземпляров (NISP –
Археология, этнография и антропология Евразии 2 (46) 2011
Number of Identified Specimens) и минимальному количеству особей (MNI – Minimum Number of Individuals). Данные единицы широко используются в зооархеологии [Lyman, 2008; Reitz, Wing, 1999].
Описание стоянкии история исследования
Многослойное поселение Улан-Хада расположено в одноименной бухте юго-западного побережья залива Мухор Малого моря оз. Байкал, в 187 км к СВ от г. Иркутска и в 4,5 км к ССЗ от пос. Сахюрта Ольхонско-го р-на Иркутской обл. (рис. 1, 2). Объект был открыт и впервые раскопан экспедицией Российской академии наук, возглавляемой Б.Э. Петри, в 1912–1913, 1916 гг. Ею выявлено 12 культурных слоев, датированных неолитом и «бескерамическим неолитом» (мезолитом) [Петри, 1916, 1926]. В 1959 г. в результате масштабных работ, проводившихся под руководством М.П. Грязнова и М.Н. Комаровой Иркутской экспедицией Ленинградского отделения Института археологии АН СССР, на поселении выделены комплексы бронзового и железного веков [Грязнов, Комарова, 1992]. Дополнительные стратиграфические работы выполнены Л.П. Хлобыстиным в 1963 г. [1964]. Комплексное междисциплинарное изучение много-
Рис. 1. Карта-схема расположения стоянки Улан-Хада.
слойного поселения проводилось Байкальским (Маломорским) отрядом Комплексной археологической экспедиции Иркутского государственного университета в 1974, 1979 (руководители Н.А. Савельев, О.И. Горюнова), 1982, 1990, 1994 гг. (руководитель О.И. Горюнова) [Горюнова, 1984; Горюнова, Савельев, 1990]. Археологические находки залегали в темных гумусированных слоях, разделенных прослойками серо-желтого песка. Генезис отложений – склоновый, осложненный эоловыми процессами. На поселении выделены 12 культурных слоев: слой XI датирован финальным мезолитом, слои X–VIII – разными периодами неолита, слои VII–I – бронзовым веком, пачка нулевых слоев – переходным периодом от бронзового к железному веку – поздним железным веком. Культурные слои бронзового века залегают в погребенной дюне, в которой насчитывается до восьми погребенных почв. На разных участках раскопов количество этих почв меняется, поэтому в ряде случаев они фиксируются как объединенные II–VII, IV–VII, V–VII культурные слои.

Рис. 2. Общий вид бухты Улан-Хада (вид с ЮЗ).
Фаунистические материалы
Исследовались 2 746 костей из всех культурных слоев, кроме слоя X (период раннего неолита) (см. таблицу ). Среди определимых костей – остатки млекопитающих следующих видов: бурый медведь ( Ursus actor ), род волчьих ( Canis ), нерпа ( Phoca sibirica ) , росомаха ( Gulo gulo ), благородный олень ( Cervus elaphus) , косуля ( Capreolus pygargus ) и лошадь ( Equus sp.). Часть материалов определена до уровня семейств оленьих (Cer-vidae) и мышей (Muridae), отрядов грызунов (Rodentia), хищников (Carnivora) и парнокопытных (Artiodactyla). Кости птиц принадлежат роду орланов ( Haliaeetus ). Среди рыб определены окунь ( Perca fluviatilis ), щука ( Esox lucius ), осетр ( Acispenser baeri baicalensis ), cемей-ство карповых (Cyprinidae, в т.ч. сорога ( Rutilus rutilus lacustris )) и представители лососевых (Salmonidae, в т.ч. представители рода сиговых ( Coregonus )).
Культурный слой XI. Фаунистические материалы относятся к финальному мезолиту (7,0–6,6 тыс. л.н.). Костные остатки в основной массе недиагностич- ны (95 %), остальные принадлежат нерпе, крупному парнокопытному и осетру. Минимальное количество особей не превышает единицы. Кости нерпы, видимо, принадлежат взрослой особи: на дистальном эпифизе 4-го метатарсала видна линия сроса. Формирование этого эпифиза у представителей рода Phoca происходит в 8–12 лет [Stora, 2000]. Кости со следами модификации немногочисленны и представлены подработанным фрагментом трубчатой кости, двумя обломками изделий (один из них, возможно, от обоймы вклады-шевого орудия) и жженой кости нерпы.
Культурный слой IX. Фаунистические материалы слоя ассоциируются с периодом развитого неолита (радиоуглеродные даты: 4 030 ± 115 л.н. (СОАН-3335), 4 560 ± 100 л.н. (ЛЕ-1282); 5,5–4,3 тыс. л.н.). Кости животных немногочисленны, среди них определены кости рода волчьих. Все они представлены фрагментами черепа со следами воздействия огня. Данные остатки, как и недиагностичные жженые кости, принадлежат, вероятно, одной особи, но поскольку они сильно фрагментированы, вид (волк или собака) опре-
Кости разных категорий животных на стоянке Улан-Хада, ед.
|
Taксон |
Культурные слои |
Всего |
|||||||||||||||
|
0 |
I |
Iв |
Iн |
II |
III |
IV |
V |
VI |
VII |
IV–VII |
V–VII |
II–VII |
VIII |
IX |
XI |
||
|
Mammalia |
54 |
75 |
50 |
57 |
89 |
17 |
8 |
3 |
– |
– |
11 |
47 |
17 |
180 |
51 |
164 |
823 |
|
Equus sp. |
6 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
6 |
|
Artiodactyla |
2 |
4 |
– |
2 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
1 |
10 |
|
Cervidae |
4 |
– |
2 |
– |
6 |
– |
13 |
– |
– |
– |
– |
1 |
1 |
20 |
– |
– |
47 |
|
Cervus elaphus |
– |
1 |
5 |
– |
4 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
10 |
|
Capreolus pyg. |
1 |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
2 |
|
Carnivora |
– |
1 |
2 |
2 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
5 |
|
Ursus actor |
– |
– |
– |
– |
2 |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
3 |
|
Canidae |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
|
Canis sp. |
– |
– |
17 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
2 |
– |
19 |
|
Phoca sibirica |
5 |
18 |
26 |
36 |
42 |
1 |
29 |
– |
2 |
10 |
3 |
4 |
10 |
37 |
2 |
3 |
228 |
|
c.f. Gulo gulo |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
|
Rodentia |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
3 |
– |
– |
– |
– |
– |
4 |
|
Muridae |
– |
4 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
4 |
|
Aves |
1 |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
3 |
|
Haliaeetus sp. |
– |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
|
Pisces |
1 |
31 |
– |
– |
13 |
208 |
91 |
197 |
4 |
– |
20 |
– |
47 |
315 |
– |
– |
927 |
|
Salmonidae |
– |
– |
– |
– |
– |
258 |
4 |
17 |
2 |
– |
1 |
– |
1 |
50 |
– |
– |
333 |
|
Coregonus sp. |
– |
– |
– |
– |
– |
16 |
1 |
15 |
– |
– |
1 |
– |
– |
10 |
– |
– |
43 |
|
Cyprinidae |
– |
– |
– |
– |
– |
– |
– |
2 |
– |
– |
– |
– |
– |
– |
– |
– |
2 |
|
Rutilis r.lac. |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
|
Perca fluviatilis |
1 |
10 |
– |
– |
3 |
61 |
102 |
9 |
– |
– |
1 |
– |
8 |
54 |
– |
– |
249 |
|
Esox lucius |
1 |
1 |
– |
1 |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
1 |
– |
– |
5 |
|
Acispenser b.c. |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
1 |
– |
– |
– |
1 |
– |
4 |
7 |
|
Неопределимые |
– |
– |
– |
– |
– |
–7 |
– |
– |
– |
– |
– |
– |
– |
5 |
– |
– |
12 |
|
Всего |
78 |
145 |
102 |
99 |
162 |
568 |
249 |
245 |
8 |
12 |
40 |
52 |
84 |
675 |
55 |
172 |
2 746 |
делить невозможно. Остальные материалы – две фаланги нерпы, принадлежащие немолодой особи (сросшиеся дистальные эпифизы фаланг [Ibid.]).
Культурный слой VIII. Остатки животных из этого слоя датируются поздним неолитом (4 150 ± 80 л.н. (ЛЕ-1280), 4 060 ± 80 л.н. (ГИН-4877); 4,2–4,0 тыс. л.н.). Определены: нерпа, семейство оленьих и парнокопытные. Остатки млекопитающих составляют 35 % от общего количества костей. Представлены обработанные кости в виде обломка орудия, острия и изделия, орнаментированного насечками. Трубчатые кости несут следы подшлифовки (1 экз.) и воздействия огня (57 экз.).
Среди костей млекопитающих преобладают остатки нерпы (от MNI=3). Судя по срастанию эпифизов на разных элементах [Ibid.], кости принадлежали, вероятно, как минимум неполовозрелой особи, молодому индивидууму 6–7 лет и уже немолодой нерпе (старше 10–12 лет). Годовые слои в дентине одного клыка также показали возраст от 8 лет [Weber et al., 1998].
Основная масса костных остатков из культурного слоя VIII относится к ихтиофауне (64 %). Определены окунь, щука, осетр и представители семейства лососевых, в т.ч. рода Coregonus (сиг или омуль). Некоторые кости представителей этого рода (вероятно, сига достаточно крупного размера) принадлежали трем особям. Остальная часть коллекции – диафиз локтевой кости птицы крупного размера и пять костных остатков, недиагностичных до любой категории.
Культурные слои VII–II. Фауна пачки слоев от-но сится к периоду раннего бронзового века (4,0– 3,8 тыс. л.н.). Радиоуглеродные даты слоев: VII – 3 660 ± ± 60 л.н. (ЛЕ-883), VI – 3 710 ± 100 л.н. (ЛЕ-1279), V – 4 220 ± 120 л.н. (ЛЕ-1278) и II – 4 000 ± 50 л.н. (ГИН-4876). Видовой состав фауны достаточно разнообразен, включает парнокопытных (благородный олень и косуля), хищников (медведь и нерпа), птиц (род орланов) и рыб. В слое также найдены кости грызуна небольшого размера. В слое II в кострище было зафиксировано 30 неопределимых костей млекопитающих. В слоях II и объединенном VII–II отмечены фрагмент трубчатой кости с элементами обработки и обломок орудия с насечкой. Следы воздействия огня имеются на недиагностичном фрагменте и фаланге нерпы.
Среди остатков млекопитающих преобладают кости нерпы (MNI=5). Следы разделки туш этих животных в виде порезов отмечены на шейном позвонке и 1-м метатарсале. На фаланге и 1-м метатарсале прослежены признаки патологии кости. По проявлениям процесса срастания эпифизов на разных элементах [Stora, 2000] можно предположить, что в отложениях представлены остатки неполовозрелой особи ок. 4 лет, взрослой 8–10 лет, а также взрослых и уже немолодых индивидуумов (10–12 лет и старше).
Наибольшее количество костей в слоях VII–II принадлежит ихтиофауне (76 % от общего количества).
Найдены остатки представителей семейства лососевых (минимум пяти особей, в т.ч. рода сиговых), окуня, карповых (в т.ч. сороги), щуки и осетра. Половина остатков ихтиофауны (560 ед.) – из скопления костей, найденных в слое III (кв. 13–23). Кости принадлежат окуню (61 ед.), минимум двум особям; сигу/омулю (16 ед.), минимум четырем особям; представителям семейства лососевых (сиг/омуль/хариус, 258 ед.), минимум пяти особям; 208 ед. – недиагностичные.
Культурные слои I, Iв и Iн. На ряде участков раскопа слой I разделяется стерильной прослойкой на нижний и верхний. В тех местах, где разделение невозможно было проследить, он рассматривался как объединенный. Нижний слой I датируется развитым бронзовым веком (3,8(3,6)–3,1 тыс. л.н.); для него имеются даты: 3 800 ± 100 л.н. (ЛЕ-1277) и 3 620 ± ± 50 л.н. (ГИН-4875). Материалы верхнего слоя I относятся к позднему бронзовому веку (3,0–2,8 тыс. л.н.). Фаунистические материалы слоев представлены 346 экз. Они представляют в основном млекопитающих (86 %) – парнокопытных (семейства оленей крупных размеров и благородного оленя) и хищников (нерпа, собака и росомаха). В слоях также найдены ко сти мыши и ихтиофауны. Кости рыб немногочисленны, принадлежат окуню и щуке.
В рассматриваемых отложениях, как и в предыдущих слоях, среди остатков млекопитающих преобладают кости нерпы (23 % от фаунистических остатков в слое). Наиболее интересен верхний слой I: в нем представлены 44 кости нерпы минимум двух особей – по срастанию эпифизов молодой, но уже половозрелой и достаточно немолодого индивидуума. Среди найденных элементов скелетов преобладают кости головы и конечностей животных (рис. 3).
Следы разделки туш нерпы в виде порезов зафиксированы на пяти костях – правой лопатке, правой лучевой кости, левой тазовой кости, 1-м метатарсале и 5-м метакарпале (рис. 3). Все эти следы ассоциируются с процессом разделения туш нерп на анатомические части [Boyle, 2005]. Другие следы разделки (снятия шкуры или отделения мяса от кости) не обнаружены. Отделение плечевого пояса от лопатки зафиксировано в виде трех параллельных порезов на последней. Четыре глубокие параллельные бороздки, расположенные перпендикулярно оси на тазовой ко сти, свидетельствуют об отделении головки бедренной кости от таза. Этот же процесс получил отражение на лучевой кости: на внутренней стороне проксимальной и дистальной части имеются следы, связанные с отделением этой части конечности от плечевого пояса и ласт соответственно. Порезы, расположенные перпендикулярно основной оси костей на дорсальных сторонах метатарсала (около проксимальной части) и метакар-пала (около дистального эпифиза), ассоциируются с процессом отделения ласт от туловища.
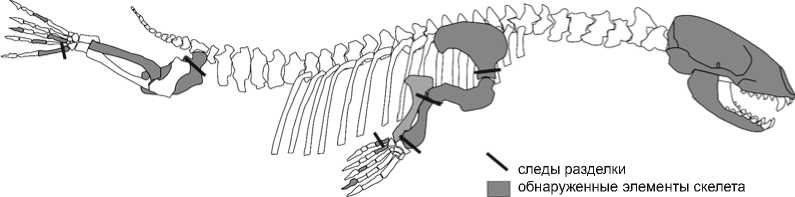
Рис. 3. Элементы скелета нерпы из верхнего слоя I.
следы разделки
■ обнаруженные элементы скелета
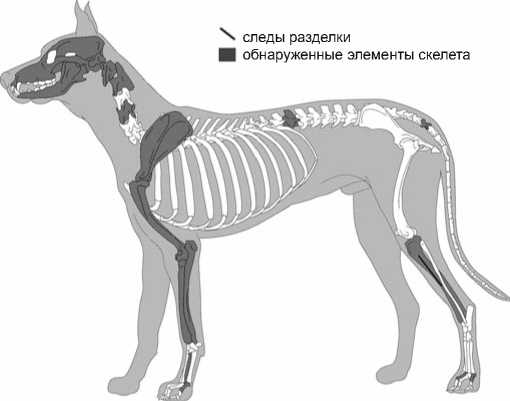
Рис. 4. Элементы скелета собаки из верхнего слоя I.
волчьих, нерпы и грызуна небольшого размера. Минимальное количество особей не превышает единицы; исключением является нерпа: остатки этого вида ассоциируются с двумя особями, по состоянию срастания дистального эпифиза плечевой кости [Stora, 2000] – половозрелой и неполовозрелой.
Остальные материалы немногочисленны и принадлежат птице и ихтиофауне. Рыбы представлены единичными костями щуки и окуня. Сохранность фаунистических остатков в слое в целом хорошая, кроме костей лошади. Находки, демонстрирующие модификацию, единичны: возможно, рог с элементами подработки, две жженые кости и две трубчатые кости с погрызами хищников.
Обсуждение материалов и выводы
Остатки нерпы, найденные в нижнем слое I, представлены костями минимум двух особей. Судя по степени срастания эпифизов на элементах, кости принадлежали молодому и немолодому индивидуумам. Это подтверждается данными изучения годовых слоев в дентине трех клыков из этого слоя: возраст особей 6 и более 10 лет [Weber et al., 1998].
Интерес представляют остатки представителя волчьих (собака или волк), обнаруженные в верхнем слое I. В общей сложности зафиксировано 19 костей. Среди них 17 элементов рода Canis и 2 – хищника среднего размера, скорее всего, тоже рода Canis , вероятно собаки. Возраст особи точно определить пока не удалось; судя по признакам завершения срастания последнего эпифиза, ей более 1,5 лет [Silver, 1969]). На большой берцовой кости имеются следы пореза в виде линии, параллельной длинной оси кости (рис. 4). След достаточно необычный, и связать его с чем-либо определенным на данный момент трудно.
Культурный слой 0. Фаунистические материалы этого слоя относятся к переходному периоду от бронзового к железному – позднему железному веку (2,7–1,0 тыс. л.н.). Основную часть коллекции составляют кости млекопитающих (74 ед.) – лошади, косули, представителей семейства оленьих крупного размера,
Фаунистические о статки с многослойной стоянки Улан-Хада принадлежат в основном млекопитающим (42 %) и ихтиофауне (57 %); несколько костей представляют птиц (см. таблицу ). Кости млекопитающих найдены в каждом проанализированном культурном слое (рис. 5); наибольшее количество – в слоях позднего неолита (VIII) и ранней бронзы (VII–II).
Самыми многочисленными являются остатки нерпы; они зафиксированы в слоях всех периодов – от финального мезолита до позднего железного века, по количе ству их несколько больше в раннебронзовых отложениях. Кости нерпы из слоев бронзового века не сут следы разделки, указывающие, возможно, на обработку туш непосредственно на стоянке. Добытые животные – как правило, достаточно взрослые половозрелые особи, об этом свидетельствуют размеры костей, признаки срастания эпифизов на элементах и данные изучения годовых слоев в дентине клыков.
Наличие остатков немолодых нерп в слоях Улан-Хады отличает поселение от памятников западного побережья Большого моря оз. Байкал в плане выбора древними охотниками категорий животных. Например, на стоянках, расположенных на западном побережье Большого моря Тышкинэ II, Тышкинэ III и Саган-Заба II, где на нерпу вели специализированную охоту в весеннее время во время коллективных леж- бищ, представлены ко сти в основном животных до года и молодых неполовозрелых особей [Weber et al., 1998; Горюнова и др., 2007]. Вероятно, в Улан-Хаде охота на нерпу не была специализированной, охотники брали тех животных (как правило, взрослых, немолодых особей), которые оставались на зимовку в Малом море или случайно заплывали в его заливы.
В слоях развитого неолита и позднего бронзового века найдены остатки волчьих – волка или собаки (см. таблицу , рис. 4). Интересно, что в первом обнаружены обожженные фрагменты черепа, во втором – часть костяка с порезом на большой берцовой кости. Остатки рода волчьих часто представлены в поселенческих комплексах в отложениях начиная с мезолита (Усть-Хайта, Усть-Белая и др.), а также в погребениях неолита и бронзового века Прибайкалья [Медведев, 1971; Конопацкий, 1982; Древние погребения…, 2004; Клементьев, Игумнова, Савельев, 2005]. Фрагменты костяка и черепа соба-ки/волка на стоянках Малого моря встречены впервые.
Необычным для памятников на Малом море является наличие в слоях II и IV раннего бронзового века костей медведя, точнее костей полового члена (баку-лум) минимум двух особей. Такие остатки отмечены впервые на археологических объектах Малого моря, до этого они фиксировались только в китойских ранненеолитических могильниках Прибайкалья [Базалий-ский, Вебер, 2008].
Остатки парнокопытных представлены несколькими костями благородного оленя и косули. Они встречаются почти во всех хронологических срезах Улан-Хады. Кости птиц и грызунов также немногочисленны. Среди остатков птиц удалось проанализировать только одну кость, принадлежащую роду орланов из нижнего слоя I (развитый бронзовый век), остальные кости недиагностичны для видового определения.
Остатков рыб в слоях позднего неолита и раннего бронзового века значительно больше, чем в отложениях других периодов (см. рис. 5). В слоях финального мезолита и развитого/позднего бронзового века кости рыб встречены в небольшом количестве. Среди остатков ихтиофауны преобладают кости лососевых (вероятно, сига) и окуня (см. таблицу ). Также зафиксированы остатки щуки, осетра и представителей семейства карповых, в т.ч. сороги. Вероятное присутствие костей сига в остеологических коллекциях ихтиофауны Улан-Хады позволяет предполагать, что стоянка использовалась, возможно, осенью (октябрь); именно в это время сиг заходит на нерест в Мухорский залив, в начале которого расположена бухта Улан-Хада [Кожов, Мишарин, 1958; Номоконова, Лозей, Горюнова, 2009а]. Однако бухту могли посещать и в другое время.
Сиговые относятся к глубоководным рыбам. Улан-Хада является единственной стоянкой, где найдено большое количество сиговых по сравнению с другими поселениями Малого моря (Берлога и Итырхей) [Losey,
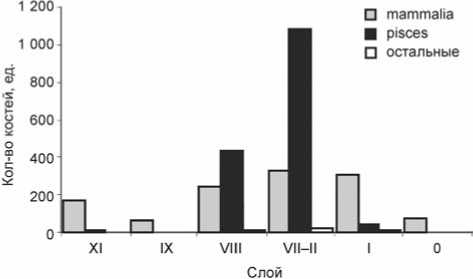
Рис. 5. Распределение категорий животных по слоям.
Nomokonova, Goriunova, 2008; Номоконова, Лозей, Горюнова, 2009б]. Говорить о том, были ли сиговые целенаправленно добываемыми видами рыб или присутствие их остатков в отложениях носит случайный характер, сегодня затруднительно. Остальные виды рыб, отмеченные на стоянке, относятся к круглогодичным обитателям бухты Улан-Хада и других мелководных зон Малого моря и характеризуют прибрежный рыболовный промысел.
Как показывает сравнение видового разнообразия животных в хронологическом аспекте, фауна финального мезолита (слой XI) и развитого неолита (слой IX) немногочисленна и представлена в основном нерпой, семейством оленьих, волком/собакой, осетром и неди-агностичными фрагментами. Слои позднего неолита (VIII) и раннего бронзового века (VII–II) характеризуются значительно большим количеством костных остатков по сравнению с другими культурными отложениями. В видовом составе млекопитающих отмечено появление косули и благородного оленя, а также медведя и орлана. Расширение ихтиофауны произошло за счет появления большого количества представителей сиговых и окуня, а также наличия щуки и карповых (сороги). Материалы развитого/позднего бронзового века из слоя I и 0 переходного от бронзового к железному – позднему железному веку не свидетельствуют о значительном изменении в видовом составе млекопитающих, но отражают резкое сокращение количества ихтиофауны.
В целом бухта Улан-Хада использовалась древними жителями с финального мезолита; ее наиболее активное освоение происходило в неолите и бронзовом веке. Судя по остаткам костей, объектами промысла были в основном нерпа и парнокопытные (косуля и благородный олень). Охотились также на хищников (медведь, собака), но с какой целью – сказать трудно. Рыболовство также имело большое значение, особенно 4,2–3,8 тыс. л.н. (бронзовый век). Основными объектами лова являлись представители сиговых и окунь, а также сорога, щука и осетр.
Фаунистические материалы Улан-Хады представляют собой информативный источник для реконструкции использования бухты древними обитателями региона на протяжении голоцена. Судя по видам добывавшихся животных, стоянка отражает не только локальный спектр фауны Приольхонья, но и культурную специфику в охоте на хищников.
К сожалению, пока трудно определить, происходили ли со временем изменения в выборе того или иного вида животных для промысла. Зооархеологи-ческие материалы стоянки Улан-Хада недостаточно представительны в количественном отношении для такого рода реконструкций. Тем не менее авторами данной работы планируется проведение комплексных исследований голоценовых фаунистических остатков с многослойных стоянок Приольхонья с обсуждением культурных, экологических и климатических факторов, формирующих хозяйственный уклад древних жителей побережья Байкала.
