Использование SNP-маркеров для идентификации локусов PL5 и PL8, контролирующих устойчивость подсолнечника к Plasmopara halstedii (Farl.) Berl. et de Toni
 Berl. et de Toni")
Автор: Рамазанова С.А., Антонова Т.С., Бадьянов Е.В., Сыксин С.В., Меркулов П.Ю.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 3 (179), 2019 года.
Бесплатный доступ
Ложная мучнистая роса, вызываемая оомицетом Plasmopara halstedii (Farl.) Berl. et de Toni, является одной из самых вредоносных болезней подсолнечника. К настоящему времени в мире обнаружено 50 физиологических рас патогена. Эффективным методом контроля возбудителя болезни является введение доминантных генов устойчивости к нему в растение-хозяина. Современные технологии генотипирования с помощью ДНК-маркеров позволяют контролировать наличие этих генов на каждом этапе селекции. Разработаны и апробированы на линиях-дифференциаторах устойчивости подсолнечника, входящих в международный тест-набор для идентификации рас P. halstedii 13 SNP-маркеров генов Pl5 и Pl8, контролирующих устойчивость к 16 расам. Из них были отобраны три SNP-маркера: 8R2, 5S2 и 5 S3, пригодные для идентификации генов Pl5 и Pl8, контролирующих устойчивость подсолнечника к возбудителю ложной мучнистой росы.
Днк-маркеры, мас, r-гены, устойчивость, подсолнечник
Короткий адрес: https://sciup.org/142222537
IDR: 142222537 | УДК: 633.854.78:575.113.1
Usage of SNP-markers for identification of loci PL5 and PL8 controlling sunflower resistance to Plasmopara halstedii (Farl.) Berl. et de Toni
Downy mildew caused by an oomycete Plasmopara halstedii (Farl.) Berl. et de Toni is one of the most dangerous sunflower diseases. At the present moment, there have been found out 50 physiological races of the pathogen. The effective method to control the pathogen is introduction of dominant genes of resistance into the host-plant. The modern technologies of genotyping containing DNA-markers allowed controlling these genes presence at the each stage of breeding process. We developed 13 SNP-markers of genes Pl5 and Pl8 controlling resistance to 16 races and evaluated them on the lines-differentiators of sunflower resistance which are included into an international testing set for identification of P. halstedii races. We selected from them three SNP-markers 8R2, 5S2 and 5S3 which can be used for identification of the genes Pl5 and Pl8 controlling sunflower resistance to downy mildew pathogen.
Текст научной статьи Использование SNP-маркеров для идентификации локусов PL5 и PL8, контролирующих устойчивость подсолнечника к Plasmopara halstedii (Farl.) Berl. et de Toni
Введение. Повсеместно распространенной и крайне опасной болезнью подсолнечника является ложная мучнистая роса, вызываемая оомицетом Plasmopara halstedii (Farl.) Berl. et de Toni. Потери урожая при условиях, благоприятных для развития болезни, могут составлять 50–
70 %. По данным разных авторов, в мире обнаружено в 2006 г. 35 рас, 41 – в 2014 г. и 50 – в 2019 г. [1; 2; 3; 4; 5]. Каждый год публикуются сообщения о возникновении новых рас. При этом ранние, менее вирулентные патотипы, такие как расы 100, 300, 700 и другие, замещаются новыми, более агрессивными [1; 2].
Одним из самых эффективных методов контроля над P. halstedii является введение в растение-хозяина доминантных генов Pl , обеспечивающих устойчивость к болезни. К настоящему времени уже картировано 33 гена Pl , расположенных в пяти группах сцепления LG: 1, 2, 4, 8 и 13 [8].
В настоящее время перспективными для использования в селекции являются кластеры генов Pl 5 и Pl 8 , контролирующие устойчивость к 16 расам P. halstedii . . Они были клонированы с помощью метода ПЦР с вырожденными праймерами, построенными на основе аминокислотной последовательности домена NBS-LRR [6; 7]. Авторы показали, что гены Pl 5 и Pl 8 относятся к non-TIR-NBS-LRR классу R-генов и картированы в тринадцатой группе сцепления на генетической карте SSR. Проведенные в последние годы исследования показали, что между генами Pl 5 и Pl 8 расположены Pl 21 , Pl 31 и Pl 32. На этой же хромосоме картирован ген Pl 22 [8]. Это, вероятно, связано с тем, что в области гена Pl 8 расположены 14 RGCs (Re-sistence Gene Candidate), т.е. «кандидатов» в гены устойчивости, и поэтому многие гены картируют в этом районе.
Целью наших исследований было разработать молекулярные маркеры генов Pl 5 и Pl 8 и изучить возможность их использования для определения наличия этих локусов в линиях-дифференциаторах устойчивости подсолнечника, используемых для идентификации рас P. halstedii .
Материалы и методы. Объектом исследования служили 14 линий-дифференциаторов устойчивости подсолнечника, входящих в международный тест-набор для идентификации рас P. halstedii .
Для выделения ДНК применяли метод, основанный на использовании лизирующего буфера, содержащего гексадеци-лтриметиламмоний бромид (СТАВ), с модификациями [11; 12]. Концентрацию ДНК в полученном препарате определяли визуально по интенсивности свечения пробы объемом 10 мкл в ультрафиолетовом свете в 1%-ном агарозном геле с добавлением 2 мкл бромистого этидия. Электрофорез препаратов ДНК проводили при напряжении 100 V в течение 30 мин в трис-ацетатном буферном растворе (ТАЕ).
Для ПЦР-анализа применили 13 пар праймеров, разработанных для маркирования локуса Pl 8 (табл. 1).
Таблица 1
Характеристика праймеров для маркирования локуса Pl 8
|
Наз ва-ние |
Последовательность |
Размер ампликона (п.н.) |
|
|
8R1 |
F:GATGATTTGGTATTGACGACAGG R:AGAAAGAGATGTGACTAAGCGTAATG G |
642 |
|
|
8R2 |
F: R |
CATTGGATCAACCCAAAAGC : ACGAGTCTGGTAAATCATGGG |
276 |
|
8R3 |
F: R |
CTGCTGCTGCCCTCG : AGAGCAATAAGCAAACAATCGC |
668 |
|
8R4 |
F: R |
AGTTGCCTGAGAATGTTGGC : CCAACTCGACATATCTTCAAACC |
608 |
|
8R5 |
F: R |
AGCGTTAGATGCTTCGTTATCG : CCCATATTGACAAAGAGTTGAGG |
632 |
|
8R6 |
F: R |
AGCGTTAGATGCTTCGTTATCG : CGTCTCTGGTAGATCGTTCACC |
624 |
|
8R7 |
F: R |
ACGACATCGTACCACCATCC : CCGATACCATACCTGAAACCG |
556 |
|
5R2 |
F: R |
CTTCTTCTTCTTTCCCTGTAGTCG : ATGTAACCCAACCCAACTCG |
601 |
|
8S1 |
F: R |
AAATGTTGCAGGGACATATAACC : TCTTCAACTCTAGTCAAGGGTGG |
470 |
|
8S2 |
F: R |
GAATCTCTCATATCCCCTACATCCC : ATACCATACCATACCTGAAACCGG |
379 |
|
5S1 |
F: R |
TCATATGCTCTTGTTTACTAGGATGGC : GGCGAAATTGGTTCCCG |
171 |
|
5S2 |
F: R |
CCATACAAATCGTTGCAGTTCC : GTCTGGTAAATCATGGATCAACTCC |
423 |
|
5S3 |
F: R |
AGTGGGGGGATCTAAATTTTCC : CGTCTCTGGTAGATCGTTCACC |
230 |
Полимеразную цепную реакцию выполняли в реакционной смеси (25 мкл) следующего состава: 67 мМ Трис-HCl (рН 8,8); 16,6 мM сульфата аммония; 1,5– 3,0 мM MgCl 2 ; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по
10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК-полимеразы (НПО «СибЭнзим», Россия). Реакции проводили в термоциклере S1000тм (BioRad, США) при следующих температурных режимах: начальная денатурация при 95 °С в течение 5 мин, далее 37 циклов с последовательной сменой температур: денатурация при 95 ºС, в течение 30 сек., отжиг праймера при 63 ºС – 30 сек., элонгация при 72 ºС – 1 мин 24 сек., и заключительная элонгация при 72 ºС в течение 10 мин.
Электрофорез продуктов амплификации проводили в геле, содержащем 2 % агарозы и SВ-буфер, с использованием камеры для горизонтального электрофореза SE-2 (Хеликон, Россия) при напряжении 200 В, силе тока 100 мА, в течение 30 мин. Для некоторых праймеров электрофорез дополнительно проводили в 8%-ном акриламидном геле в камере для вертикального электрофореза (Хеликон, Россия). Гели окрашивали бромистым этидием. Для визуализации и документирования результатов электрофореза применяли систему цифровой документации видеоизображения BIO-PRINT (Vilber Lourmat, Франция).
Результаты и обсуждение. На начальном этапе работы по поиску ДНК-маркеров для генов устойчивости Pl5 и Pl8 к P. halstedii исследовали молекулярногенетический полиморфизм шести STS (Sequence-Tagged Site) локусов ДНК [6; 7]. В результате ПЦР с этими праймерами не было получено специфичных фрагментов ДНК, характерных для линий XRQ, YVQ, 803-1, PM-17 и DM-2 – носителей кластеров генов Pl5/Pl8 [9]. Нам не удалось разделить линии на устойчивые и восприимчивые так, как это описано в работе Radwan с соавторами [6; 7]. По нашему мнению, это связано с большим размером фланкируемого праймерами участка ДНК и, как следствие, амплификаций фрагментов длиною более 1000 пар нуклеотидов. К тому же сравнение нуклеотидных последовательностей этих ге- нов показало наличие большого количества участков ДНК, совпадающих по нуклеотидному составу. Это также создает проблемы при маркировании данных локусов.
Поэтому на базе известных последовательностей 13 STS-маркеров [6] нами были созданы SNP-праймеры (Single Nucleotide Polymorphism). Работа проводилась на кафедре генетики, биотехнологии и семеноводства МСХА имени К.А. Тимирязева. С целью упрощения проведения исследования и анализа результатов для амплификации были выбраны небольшие участки: от 171 до 668 п.н. Их характеристика и нуклеотидный состав представлены в таблице 1. Температура отжига для этих праймеров подбиралась с минимальными пошаговыми различиями, которые позволили бы их комплексное применение в ПЦР -смеси в дальнейшем.
На начальном этапе разработанные SNP-праймеры апробировали на 14 линиях-дифференциаторах устойчивости подсолнечника из международного тест-на-бора для идентификации рас P. halstedii. Каждая из этих линий содержит один или несколько генов Pl, обеспечивающих их устойчивость к определенным расам патогена. Подробная информация об этих линиях, соответствующих им генах устойчивости и группах сцепления, в которых они картированы на генетической карте SSR-локусов, опубликована ранее [9; 10; 12–21].
Результаты ПЦР с разработанными SNP-маркерами показали, что только с двумя (5R2 и 5S1) не было отжига праймера на матрице, с остальными одиннадцатью были получены продукты амплификации.
По локусу 8R1 при амплификации ДНК линий подсолнечника выявлен фрагмент длиною около 642 п.н. только у линии DM-2 – носителя генов Pl5, Pl11, Pl12 [10]. У остальных линий, содержащих локус Pl5 (803-1, YVQ, РМ-17 и XRQ), рав- но как и у линий с его отсутствием, этой фракции выявлено не было.
С праймером 8R2 получены продукты амплификации разной длины (рис. 1). У линий XRQ и DM-2 получены фракции, отличающиеся от остальных (дорожки 2 и 14). Также у линий YVQ и PM-17 (дорожки 9–10) обнаружены фрагменты одной длины, не выявленные у остальных. Однако для линии 803-1 не получено таких фрагментов (дорожка 13). По нашему мнению, апробацию этого праймера необходимо продолжить на других линиях, являющихся носителями генов Pl 5 и Pl 8.
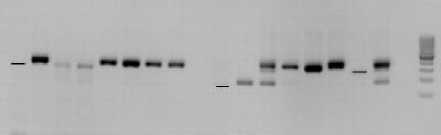
1 2 3 4 5 6 7 8 9 10 И 12 13 14 15 16 К"М
Рисунок 1 – Фореграмма продуктов амплификации ДНК линий-дифференциаторов устойчивости подсолнечника с праймером 8R2. Дорожки: 1 – RHA419;
2 – XRQ; 3, 4 – 83HR4RM; 5, 6 – HIR 34;
7, 8 – PSC8; 9 – YVQ; 10 – PM-17; 11 – HAR-5;
12 – HAR-4; 13 – 803-1; 14 – DM-2;
15 – RHA274; 16 – RHA265, Kˉ – отрицательный контроль, M – маркер молекулярного веса, 2019 г. (ориг.)
С праймером 8R3 отжиг состоялся, однако вместо ожидаемого фрагмента длиною 668 п.н. была выявлена фракция размером около 1100 п.н. Эти фрагменты присутствуют почти у всех линий. Аналогичный результат получен и с праймером 8R4. Получены фрагменты ДНК разных размеров: от 750 до 800 п.н., однако не было выявлено фрагмента специфичного для линий с локусами Pl 5 и Pl 8. С праймером 8R7 также не удалось получить ожидаемый фрагмент размером 556 п.н., а у всех образцов был амплифицирован фрагмент длиною 100 п.н. Точно также и с праймером 8S1 не было получено нужной фракции и не выявлено полиморфизма.
C праймером 8R5 получена фракция размером 632 п.н. только у линий RHA-19 и HA-335, носителей генов Pl arg и Pl 6 соответственно. У остальных линий не выявлено амплифицированных фрагментов.
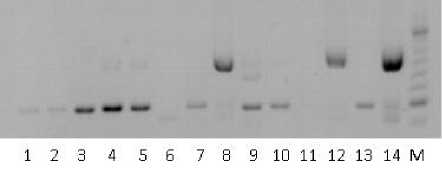
Рисунок 2 – Фореграмма продуктов амплификации ДНК линий-дифференциаторов устойчивости подсолнечника с праймером 8S2. Дорожки: 1 – RHA419, 2 – XRQ, 3 – 83HR4RM, 4 – HIR 34, 5 – PSC8, 6 – YVQ, 7 – Ha335, 8 – HAR-5, 9 – HAR-4, 10 – 803-1, 11 – PM-17, 12 – DM-2, 13 – RHA274, 14 – RHA265, M – маркер молекулярного веса, 2019 г. (ориг.)
На рисунке 2 показано, что при амплификации ДНК с праймером 8S2 у девяти линий получены четкие фракции размером 500 п.н., пока не идентифицированные. У линий YVQ (дорожка 6) (носитель гена Pl 8 ) и RHA265 выявлена фракция длиною 423 п.н. По нашему мнению, маркер 8S2 можно использовать для идентификации данного локуса, однако это требует дополнительных исследований.
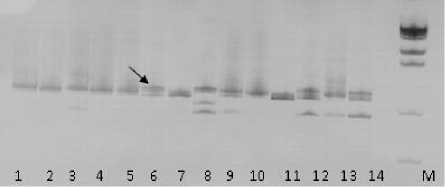
Рисунок 3 – Фореграмма продуктов амплификации ДНК линий дифференциаторов устойчивости подсолнечника с праймером 8S3. Дорожки: 1 – RHA419, 2 – XRQ, 3 – 83HR4RM, 4 – HIR 34, 5 – PSC8, 6 – YVQ, 7 – Ha335, 8 – HAR-5, 9 – HAR-4, 10 – 803-1, 11 – PM-17, 12 – DM-2, 13 – RHA274, 14 – RHA265,
M – маркер молекулярного веса, 2019 г.
(ориг.)
С праймером 8S3 были получены фракции размером 230 п.н., но полиморфизм удалось выявить только при электрофорезе в 8%-ном акриламидном геле (рис. 3). Так же, как и в случае с праймером 8S2, у линии YVQ (носитель гена Pl 8 ) выявлены фракции, не обнаруженные у других линий. По нашему мнению, SNP-праймеры 8S3 и 8S2 пригодны для идентификации локуса Pl 8 в селекционном материале ВНИИМК.
Заключение. Таким образом, наши исследования показали, что три сконструированных нами SNP-праймера – 8R2, 5S2 и 5S3 можно использовать для маркирования локусов Pl 5 и Pl 8 в линиях подсолнечника. Эти маркеры могут стать звеньями системы ДНК-маркеров для идентификации указанных генов и проведения маркер-вспомогательной селекции на устойчивость подсолнечника к возбудителю ложной мучнистой росы.
Список литературы Использование SNP-маркеров для идентификации локусов PL5 и PL8, контролирующих устойчивость подсолнечника к Plasmopara halstedii (Farl.) Berl. et de Toni
- Gulya T.J. Distribution of Flasmopara halstedii races from sunflower around the world // Advances in Downy Mildew Research / Lebeda A., Spencer-Phillips P.T.N. (eds). - Czech Republic, Olomouc, 2007. - Vol. 3. - Р. 121-134.
- Viränyi F., Gulya T.J., Tourvierille de Labrouhe D. Recent changes in the pathogenic variability of Plasmopara halstedii (sunflower downy mildew) populations from different continents // Helia. - 2015. - V. 38. - P. 149-162.
- Sedlářová M., Pospíchalová R., Drábková Trojanová Z., Bartůšek T., Slobodianová, L., Lebeda A. First report of Plasmopara halstedii new races 705 and 715 on sunflower from the Czech Republic - short communication // Plant Protect. Sci. - 2016. - V. 52. - 182187. DOI: 10.17221/7/2016-PPS
- Spring O., Zipper R. New highly aggressive pathotype 354 of Plasmopara halstedii in German sunflower fields // Plant Protect. Sci. - 2018. - V. 54. - P. 83-86. DOI: 10.17221/99/2017-PPS
- Spring О. Spreading and global pathogenic diversity of sunflower downy mildew - Review // Plant Protect. Sci. - 2019. - V. 55. - Р. 149-158. DOI: 10.17221/32/2019-PPS
- Radwan O., Bouzid, M.F., Vear F., Philippon J., Tourvieille de Labrouhe D., Nicolas F., Mouzeyar S. Identification of non-TIR-NBS-LRR markers linked to the locus for resistance to downy mildew in sun flower // Theoretical and Applied Genetics. - 2003. - V. 106. - P. 1438-1446.
- DOI: 10.1007/s00122-003-1196-1
- Radwan O., Bouzidi M.F., Nicolas P., Mouzeyar S. Development of PCR markers for the locus for resistance Flasmopara halstedii in sunflower, Helianthus annuus L. from complete CC-NBS-LRR sequences // Theoretical and Applied Genetics. - 2004. -V. 109. - P. 176-185.
- DOI: 10.1007/s00122-004-1613-0
- Pecrix Y., Penouilh-Suzette C., Munos S., Vear F., Godiard L. Ten broad spectrum resistances to downy mildew physically mapped on the sunflower genome // Front. Plant Sci. - 2018. - V. 9.
- DOI: 10.3389/fpls.2018.01780
- Рамазанова С.А., Антонова Т.С. К вопросу о маркировании локусов P1, контролирующих устойчивость подсолнечника к возбудителю ложной мучнистой росы // Масличные культуры. - 2019. - Вып. 1 (177). - С. 17-23.
- Rahim M., Jan C.C., Gulya T.J. Inheritance of resistance to sunflower downy mildew races 1, 2 and 3 in cultivated sunflower // Plant Breeding. - 2002. - V. 121. - P. 57-60.
- Jocic S., Miladinovic D., Imerovski I., Dimitrijevic A., Cvejic S., Nagl N., Kondic-Spika A. Towards sustainable downy mildew resistance in sunflower // Helia. - 2012. - No 35. - P. 61-72.
- Dußle C.M., Hahn V., Knapp S.J., Bauer E. Plarg from Helianthus argophyllus is unlinked to other known downy mildew resistance genes in sunflower // Theoretical and Applied Genetics. - 2004. - V. 109 (5). - Р. 1083-1086.
- DOI: 10.1007/s00122-004-1722-9
- Imerovski I., Dimitrijevic A., Miladinovic D., Jocic S., Dedic B., Cvejic S., Surlan-Momirovic G. Identification and validation of breeder-friendly DNA markers for Plarg gene in sunflower // Molecular Breeding. -2014. - V. 34 (3). - P. 779-788.
- DOI: 10.1007/s11032-014-0074-7
- Wieckhorst S., Bachlava E., Dußle C.M., Tang S., Gao W., Saski C., Bauer E. Fine mapping of the sunflower resistance locus Plarg introduced from the wild species Helianthus argophyllus // Theoretical and Applied Genetics - 2010. - V. 121 (8). - Р. 1633-1644.
- DOI: 10.1007/s00122-010-1416-4
- Bert P.F., Tourvieille de Labrouhe D., Philippon J., Mouzeyar S., Jouan I., Nicolas P., Vear F. Identification of a second linkage group carrying genes controlling resistance to downy mildew (Plasmopara halstedii) in sunflower (Helianthus annuus L.) // Theoretical and Applied Genetics. - 2001. - V. 103. - P. 992-997.
- Bertero de Romano A., Romano C., Bulos M., Altieri E., Sala C. A new gene for resistance to downy mildew in sunflower // Proc. Intern. Symposium "Sunflower Breeding on Resistance to Diseases", Russia, Krasnodar, 2010, June 23-24. - P. 141-146.
- Mulpuri S., Liu Z., Feng J., Gulya T. J., Jan C.-C. Inheritance and molecular mapping of a downy mildew resistance gene, Pl13 in cultivated sunflower (Helianthus annuus L.) // Theoretical and Applied Genetics. -2009. - V. 119 (5). - P. 795-803.
- DOI: 10.1007/s00122-009-1089-z
- Liu Z., Gulya T.J., Seiler G.J., Vick B.A., Jan C-C. Molecular mapping of the Pl16 downy mildew resistance gene from HA-R4 to facilitate marker-assisted selection in sunflower // Theoretical and Applied Genetics. -2012. - V. 125. - P. 121-131.
- Molinero-Ruiz M. L., Dominguez J., Melero-Vara J.M. Races of Isolates of Plasmopara halstedii from Spain and studies on their virulence // Plant Disease - 2002. - V. 86 (7). - P. 736-740.
- DOI: 10.1094/pdis.2002.86.7.736
- Gedil M.A., Slabaugh M.B., Berry S., Johnson R., Michelmore R., Miller J., Gulya T., Knapp S.J. Candidate disease resistance genes in sunflower cloned using consereved nucleotide-binding site motifs: Genetic mapping and linkage to the downy mildew resistance gene Ph // Genom. - 2001. - V. 44 (2). - Р. 205-212.
- Ahmed S., Tourvieille de Labrouhe D., Delmotte F. Emerging virulence arising from hybridisation facilitated by multiple introductions of the sunflower downy mildew pathogen Plasmopara halstedii // Fungal Genetics and Biology. - 2012. - V.49 (10). - P. 847-855.
- DOI: 10.1016/j.fgb.2012.06.012
