Исследование полиморфизма биохимических признаков в роде соя - Glycine Willd

Автор: Зеленцов С.В., Петибская В.С., Мошненко Е.В.
Статья в выпуске: 2 (133), 2005 года.
Бесплатный доступ
Исследование полиморфизма биохимических признаков в роде соя - Glycine Willd. С.В. Зеленцов, В.С. Петибская, Е.В. Мошненко. «МАСЛИЧНЫЕ КУЛЬТУРЫ. Научно-технический бюллетень», ВНИИМК, 2005, вып. 2 (133) Исследован полиморфизм биохимических признаков - содержания в семенах белка и масла, активности ингибиторов трипсина и жирнокислотного состава масла у сортов культурной сои и 6 дикорастущих видов Китайского и Австралийского генетических центров. Наличие в гомологических рядах видов сои подрода Soja высокобелковых и низкоингибиторных форм открывает возможность выведения сортов сои с высоким содержанием белка и пониженной трипсинингибирующей активностью.
Короткий адрес: https://sciup.org/142150727
IDR: 142150727 | УДК: 633.853.52:581.19:575.113
Research of polymorphism of biochemical traits in the genus soya - Glycine Willd
Research of polymorphism of biochemical traits in the genus soya - Glycine Willd. Zelentsov S.V., Petibskaja V.S., and E.V. Moshnenko. "OIL CROPS" Scientific bulletin, VNIIMK, 2005 is. 2 (133). Polymorphism of biochemical traits - protein and oil cjntent in soybean seeds, activity of trypsin inhibitors and fatty acid composition of oil at soybean varieties and 6 wild-growing species of the Chinese and Australian genetic centers is investigated. Presence in homologous lines of soybean species of subgenus Soja the high protein's and low trypsin inhibitor's forms allows breeding soybean varieties with the high protein content and the lowered trypsin inhibitor activity.
Текст научной статьи Исследование полиморфизма биохимических признаков в роде соя - Glycine Willd
ВНИИ масличных культур
ИССЛЕДОВАНИЕ ПОЛИМОРФИЗМА БИОХИМИЧЕСКИХ ПРИЗНАКОВ В РОДЕ СОЯ – GLYCINE WILLD.
Селекционно-генетическое улучшение сои неразрывно связано с расширением признакового полиморфизма вновь создаваемых сортов сои, которое может достигаться путём целенаправленного выявления новых источников и доноров биохимических признаков и их сочетаний в генофонде культурной сои. Первым этапом на этом пути является определение перспективных, биологически непротиворечивых направлений их улучшения и диапазонов изменчивости.
По последней внутриродовой классификации Палмера, Хаймовица и Нельсона (Palmer et al., 1996) род Glycine представлен 18 многолетними травянистыми видами Австралийского и однолетними травянистыми видами Китайского центров происхождения, разделённых на 2 подрода: Glycine Willd . и Soja (Moench) F.J. Herm . (Hymowitz, 1970, 1987; Palmer et al., 1996)
Австралийские виды сои, входящие в подрод Glycine Willd., отличаются многолетним циклом развития, широким геномным полиморфизмом, и представляют собой наиболее архаичные формы сои. Соответственно, комплекс признаков этих видов может условно служить начальной точкой отчёта при изучении филогенеза сои (Hymowitz, 1970; Корсаков, 1982).
Подрод Soja состоит из двух видов: G. soja и G. maх (культивируемый вид). Сюда же относится спорный вид – соя изящная или тонкая Glycine gracilis Skvortzovii (Скворцов, 1927; Жуковский, 1964), в последнее время определяемый как подвид культурной сои – G. max var. gracilis Teplyak. (Теплякова, 1997).
Виды сои Китайского центра происхождения объединёны общим геномом GG (Palmer et al., 1996) и признаны эволюционно более продвинутыми из-за однолетнего цикла развития. Филогенетически наиболее архаичным видом здесь является дикорастущий вид уссурийской сои G. soja Sieb. et Zucc . (син: G. ussuriensis Reg. et Maack.). Этот вид практически всеми систематиками признан прямым предком возделываемой культурной сои G. max (Скворцов, 1927, Сунь Син-дун, 1958; Енкен, 1959; Hymowitz, 1970; Корсаков, 1982).
Реально осуществить прогноз полиморфизма биохимических признаков у культурной сои можно на основе закона гомологических рядов Н. И. Вавилова. Во всех случаях, для установления теоретически допустимого диапазона изменчивости полиморфных признаков на основе гомологической изменчивости, а также биологически допустимых направлений расширения их полиморфизма требуется сравнительное изучение экспрессии этих признаков между родственными видами (Вавилов, 1935). В связи с этим представляется актуальным изучение филогенеза биохимических признаков при установлении гомологических рядов между культурной соей и прямыми дикорастущими предковыми формами, и на этой основе определение перспективных направлений в селекции культурной сои на изменение биохимического состава семян (Корсаков, 1971; Зеленцов, 1998).
Материалы и методы. Материалом для исследований полиморфизма рода Соя ( Glycine Willd.) служили 3 сорта вида G. max ; 6 сортообразцов вида сои изящной G. max subsp. gracilis (G. gracilis) , собранных сотрудниками ВНИИР им. Н. И. Вавилова в зоне Китайского генетического центра и обозначенных нами как G. gracilis1 , а также полученных от межвидовых скрещиваний G. max и G. soja , морфологически аналогичных естественным формам G. gracilis и обозначенных нами как G. gracilis2 ; 4 сортообразца G. soja , полученных из ВНИИР (г. Санкт-Петербург) и ВНИИ сои (г. Благовещенск, Амурской обл.), а также 4 дикорастущих многолетних вида Австралийского генетического центра – G. ca-nescens, G. clandestina, G. tabacina и G. tomentella .
Все виды сои выращивались на полях селекционного севооборота ВНИИМК с соблюдением принятой в Краснодарском крае технологии возделывания сои.
Биохимические анализы качества зерна сои проводили в отделе биохимии ВНИИ масличных культур. Оценку сортов культурной сои и коллекционных сортообразцов других видов сои на содержание белка, масла и трипсинингибирующей активности белка (ТИА) проводили путём инфракрасной спектроскопии. Жирно-кислотный состав масла определяли методом газожидкостной хроматографии.
Результаты и обсуждение. Эффективная селекция культурной сои на изменение биохимических показателей началась только в конце XX века и велась, в основном, в направлении увеличения содержания белка, снижения ТИА, а также изменения содержания отдельных жирных кислот в масле. В некоторых случаях были получены обнадёживающие результаты. Поэтому уровень экспрессии биохимических признаков у большинства форм даже в пределах G. max в целом можно считать сложившимся естественно (табл. 1).
Таблица 1 – Основные биохимические признаки у видов сои подродов
Soja (Moench) F.J. Herm. и Glycine Willd.
ВНИИМК, 2001-2003 гг.
|
Сорто-образец |
Вид |
Геном |
Содержание белка, % |
Содержание масла, % |
ТИА, мг/г |
|
Подрод Soja (Moench.) F.J. Herm. |
|||||
|
Лань |
G. max |
GG |
42,0±3,5* |
22,2±2,4 |
22,6±4,8 |
|
Лира |
«» |
«» |
40,9±3,6 |
22,3±2,0 |
23,0±5,3 |
|
Куниц |
«» |
«» |
41,5±1,1 |
20,8±0,3 |
22,6±1,4 |
|
К-5367 |
G. gracilis1 |
GG |
44,8±1,0 |
16,2±0,3 |
15,1±1,0 |
|
К-5683 |
«» |
«» |
43,7±1,1 |
15,5±0,7 |
15,1±1,1 |
|
К-521 |
«» |
«» |
43,2±1,6 |
15,9±0,1 |
16,0±1,0 |
|
К-5141 |
G. gracilis2 |
GG |
43,4±1,4 |
17,9±0,6 |
17,7±1,2 |
|
К-5142 |
«» |
«» |
42,8±1,4 |
18,1±0,6 |
18,6±1,9 |
|
К-4947 |
«» |
«» |
41,3±1,9 |
19,6±0,9 |
21,2±2,6 |
|
К-1007 |
G. soja |
GG |
46,4±0,3 |
14,7±0,4 |
11,9±0,4 |
|
№ 93 |
«» |
«» |
46,8±0,1 |
12,0±0,6 |
8,5±0,9 |
|
№ 133 |
«» |
«» |
46,4±0,8 |
11,1±0,3 |
8,3±0,4 |
|
ZYD 403 |
«» |
«» |
47,8±0,8 |
10,9±0,4 |
6,9±1,0 |
|
Подрод Glycine Willd. |
|||||
|
б/н |
G. clandestina |
A1A1 |
38,4±1,6 |
10,6±0,1 |
9,5±4,3 |
|
б/н |
G. canescens |
AA |
38,6±0,5 |
11,2±1,6 |
12,0±1,3 |
|
б/н |
G. tomentella |
DD2 |
38,1±0,7 |
12,7±0,6 |
16,7±0,1 |
|
б/н |
G. tabacina |
B 2 B 2 |
37,7±1,1 |
9,5±0,6 |
13,9±0,5 |
Примечание: * здесь и далее среднее значение признака ± стандартное отклонение
Как следует из данных, представленных в табл. 1, минимальным средним содержанием белка в семенах отличались многолетние австралийские виды сои с диапазоном варьирования от 37,7 до 38,6 %. В подроде Soja наименьшее количество белка накапливали культурные сортообразцы G. max . А наибольший уровень белка был у дикорастущей уссурийской сои (до 47,8 %). Филогенетически более продвинутые формы G. gracilis снизили содержание сырого протеина в среднем на 3 %.
Дикорастущие виды подрода Glycine отличались минимальным количеством масла в семенах. Например у вида G. tabacina в среднем за 3 года его количество составило всего 9,5 %, у вида G. tomentella – 12,7 %.
При сравнительном изучении форм сои Китайского генетического центра максимальное количество масла было выявлено у культурных сортов G. max , в среднем за 2001-2003 гг. составляющее 21,8 %, с диапазоном варьирования от 20,8 до 22,3 %. Соя изящная накапливала несколько меньше масла – в среднем 17,2 %, а минимальное среднее количество масла – 12,2 % было обнаружено у вида G. soja.
Возможно, пониженный уровень белка и масла у дикорастущих видов может определяться укороченным периодом роста и развития бобов, который ограничивает дальнейшее накопление в семенах этих компонентов, и увеличивает тем самым долю клетчатки. Вероятны также видовые генетические различия в экспрессивности этих показателей.
Увеличение ТИА в филогенезе рода Soja шло более высокими темпами, по сравнению с изменениями в накоплении масла. Например, минимальное значение этого показателя у уссурийской сои составило в условиях 2001 г. всего 6,0 мг/г, тогда как у некоторых культурных образцов активность ингибиторов трипсина достигала 29,0 мг/г (например, в сорте Лира в 2003 г.).
Теоретический и практический интерес представляет сравнительное изучение соотношения жирных кислот в масле различных видов сои. Существующие мировые тенденции в потребности отдельных компонентов соевого масла имеют разнонаправленный характер. В частности, пищевая промышленность предъявляет требования к увеличению мононенасыщенной олеиновой кислоты (С18:1), которая позволяет существенно увеличить термо- и оксистабильность масла. В то же время объединённый комитет ФАО/ВОЗ подчёркивает важность потребления организмом человека полиненасыщенных кислот – линолевой и линоленовой.
Сравнительный жирно-кислотный состав масел различных видов сои приведён в табл. 2.
Как следует из приведённых в табл. 2 данных, жирно-кислотный состав масла в пределах исследуемых подродов сои заметно различается. Австралийские дикорастущие многолетние виды в среднем имели практически удвоенное, по сравнению с сортами вида G. max содержание линоленовой кислоты в масле. Максимальное количество C18:3 обнаружено у видов G. clandestina и G. canescens – 30,5 и 29,5 % соответственно.
Существенных различий между сортами G. max и G. gracilis на широте г. Краснодара не было обнаружено. У сои изящной ( G. gracilis1 ) выявлено несколько сниженное (в среднем на 5 %) содержание олеиновой (С18:1) и на 3 % увеличено содержание линоленовой (С18:3) кислот.
У дикорастущей уссурийской сои G. soja обнаружено практически удвоенное (до 18,1 %) содержание линоленовой кислоты по сравнению со средними значениями этой кислоты у культурных форм G. max .
Минимальное в подроде Soja содержание линолевой (С18:2) кислоты, составляющее в среднем около 50 %, выявлено у культурной сои. Максимальное абсолютное количество, до 62,1 %, обнаружено у сортообразцов G. soja , а минимальное, до 45,8 % – у G. max.
Различия между видами сои по содержанию пальмитиновой и стеариновой кислот были незначительными. Несколько сниженное их количество отмечено у сортооб-разцов вида G. soja.
Таблица 2 – Содержание жирных кислот в масле у сортов и форм сои подродов Soja (Moench) F. J. Herm. и Glycine Willd ., %
ВНИИМК, 2001-2002 гг.
|
Сорто-образец |
Вид |
Геном |
Жирная кислота |
||||
|
пальмитиновая, С16:0 |
стеариновая, С18:0 |
олеиновая, С18:1 |
линолевая, С18:2 |
линоленовая , С18:3 |
|||
|
Подрод Soja (Moench.) F.J. Herm . |
|||||||
|
Лань |
G. max |
GG |
10,3±0,1 |
2,6±0,2 |
22,0±1,1 |
56,5±2,1 |
8,6±1,1 |
|
Лира |
То же |
То же |
10,4±0,2 |
2,4±0,3 |
37,0±1,3 |
45,8±0,8 |
5,7±0,1 |
|
Куниц |
То же |
То же |
8,9±0,1 |
3,1±0,1 |
21,0±1,6 |
58,0±1,8 |
9,0±0,6 |
|
К-5367 |
G. gracilis1 |
GG |
9,6±0,1 |
2,0±0,5 |
18,6±0,3 |
57,1±0,9 |
12,7±0,8 |
|
К-5683 |
То же |
То же |
10,3±0,1 |
2,6±0,1 |
19,0±0,9 |
56,3±0,4 |
11,9±0,4 |
|
К-521 |
То же |
То же |
10,0±0,2 |
2,2±0,1 |
19,4±0,3 |
56,7±0,6 |
11,8±0,9 |
|
К-5141 |
G. gracilis2 |
GG |
10,4±1,1 |
2,6±0,3 |
20,0±8,3 |
57,3±4,9 |
9,8±4,8 |
|
К-5142 |
То же |
То же |
11,5±0,4 |
2,8±0,2 |
21,2±1,1 |
55,1±0,3 |
9,4±0,7 |
|
К-4947 |
То же |
То же |
10,9±0,4 |
2,7±0,5 |
20,6±2,3 |
57,2±2,3 |
8,6±0,9 |
|
К-1007 |
G. soja |
GG |
11,6±0,5 |
3,3±0,2 |
19,0±1,2 |
55,4±2,4 |
10,8±0,3 |
|
№ 93 |
То же |
То же |
9,3±0,2 |
1,8±0,4 |
10,6±2,5 |
61,1±1,7 |
17,0±0,7 |
|
№ 133 |
То же |
То же |
8,6±0,4 |
2,1±0,2 |
9,1±0,9 |
62,1±3,0 |
18,1±1,1 |
|
ZYD 403 |
То же |
То же |
9,8±0,3 |
1,9±0,2 |
10,3±1,8 |
60,3±0,1 |
17,7±2,3 |
|
Подрод Glycine Willd. * |
|||||||
|
б/н |
G. clandestina |
A1A1 |
11,4 |
3,0 |
8,4 |
46,7 |
30,5 |
|
б/н |
G. canescens |
AA |
12,3 |
2,5 |
9,5 |
46,2 |
29,5 |
|
б/н |
G. tomentella |
DD2 |
13,8 |
4,8 |
16,7 |
49,3 |
15,5 |
|
б/н |
G. tabacina |
B 2 B 2 |
15,7 |
3,8 |
14,3 |
52,9 |
13,4 |
Примечание: * данные за 2001 г.
Представляется актуальным изучение динамики филогенеза при установлении гомологических рядов по биохимическим признакам между культурной соей и прямыми дикорастущими предковыми формами подрода Soja и многолетними видами подрода Glycine Willd . , и на этой основе определения перспективных направлений в селекции сои на изменение биохимического состава семян
Большой теоретический и практический интерес представляет изучение динамики признака содержания белка в семенах в филогенезе подрода Soja . Наиболее древние в подроде формы сои G. soja отличались высоким, до 50-52 % содержанием белка. Последующее окультуривание сои с образованием последовательно видов G. gracilis и G. max сопровождалось снижением содержания белка в семенах (рис. 1).
Можно ожидать, что установленная динамика эволюции этого признака сохранится, и дальнейшая селекция сои без учета содержания белка будет сопровождаться снижением этого показателя в семенах.
Уже в настоящее время, большинство современных сортов селекции США имеют содержание белка 37-39 %. В связи с этим одной из наиболее актуальных задач мировой селекции в последние годы стало создание сортов сои с повышенным содержанием белка в семенах.
Средние значения содержания белка в семенах многолетних видов сои Австралийского генетического центра составляют 37-38 % с незначительными колебаниями по годам. Одной из основных причин пониженного содержания белка, наряду с генетическими особенностями синтеза белков в семенах австралийских видов, может быть стремительное прохождение всех этапов роста и развития бобов. Так, например, в условиях г. Краснодара у сортов сои местной селекции вида G. max продолжительность от цветения до созревания отдельных бобов составляет в среднем от 60 до 90 дней в зависимости от группы созревания сорта. Тогда как у всех исследуемых нами 4-х австралийских видов этот период составляет всего 28-35 дней. Дополнительным подтверждением этой гипотезы может служить высокая положительная корреляция r=0,92 между содержанием белка в семенах многолетних видов сои и продолжительностью периода от начала цветения до созревания бобов.
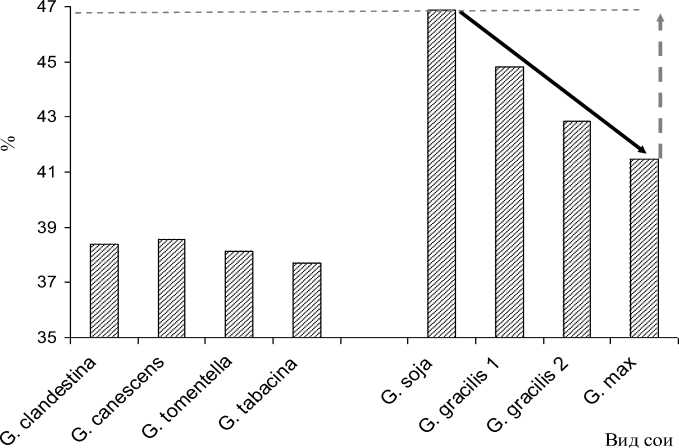
Рисунок 1 – Содержание белка в семенах различных видов сои и динамика эволюции этого признака в подроде Soja тренд эволюции признака направление возможного генетического улучшения признака верхний предел признака в исследуемом генофонде рода
Кроме этого, существенные различия по признаку содержания белка у представителей разных подродов в пределах рода Glycine , с одной стороны, свидетельствует о значительных генетических различиях между подродами Soja и Glycine , а с другой стороны, дополнительно подкрепляют уверенность в правильном отнесении отдельных видов сои в подроды по морфологическим признакам.
В целом максимальные значения этого показателя у G. soja , достигавшие в отдельные годы почти 50 % содержания белка, на основании закона гомологических рядов позволяют допустить возможность обнаружения или создания форм сои с аналогичным уровнем белка и в пределах G. max (см. рис. 1).
Поскольку соя является культурой двойного использования, немаловажное значение имеет и содержание масла в семенах. Изучение динамики накопления масла в семенах различных видов сои позволило выявить увеличенную долю масла в семенах филогенетически молодых форм (рис. 2).
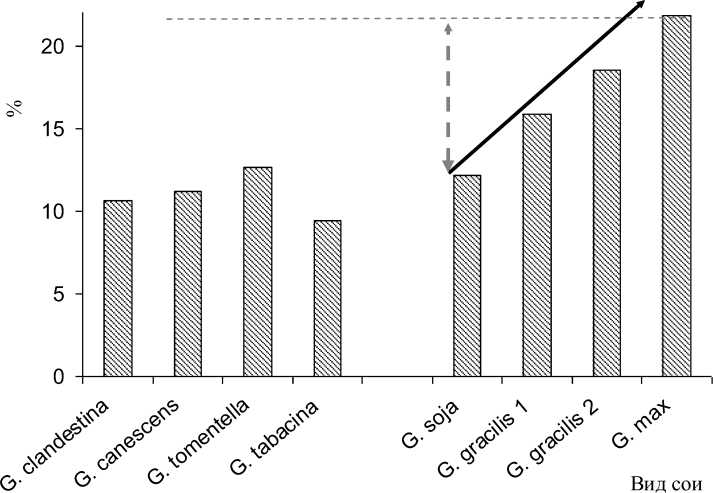
Рисунок 2 – Содержание масла в семенах некоторых видов сои и динамика эволюции этого признака в подроде Soja тренд эволюции признака направление возможного генетического улучшения признака верхний предел признака в исследуемом генофонде рода
В настоящее время содержание масла в мировой коллекции культурной сои G. max варьирует в пределах 15-27 %, крайне редко выходя за пределы установленного диапазона. Селекция в направлении увеличения этого показателя до настоящего времени остаётся практически безуспешной из-за отсутствия форм-доноров высокого содержания масла. Единственно возможным способом получения высокомасличного соевого сырья остаётся возделывание сои на бедных или истощённых почвах, не позволяющих сформировать достаточное количество белка в семенах.
Сравнительный анализ филогенеза масличности разных видов сои позволяет установить одну из причин неудач в селекции высокомасличных сортов.
Как следует из рис. 2, вид G. max имеет наибольшие значения масличности в подроде Soja . Дикорастущие предковые формы G. gracilis и G. soja имеют последовательно более низкие значения масличности семян.
Многолетние австралийские виды сои также отличаются невысоким содержанием масла в семенах. В этой группе минимальной масличностью (9,45 %) отличался вид сои табачно-бурой G. tabacina, а максимальным значением этого показателя (12,67 %) – вид сои войлочной – G. tomentella.
В связи с отсутствием доноров более высоких значений масличности у других видов сои, в рамках закона гомологических рядов отсутствует благоприятный прогноз на дальнейшее увеличение содержания масла в семенах.
Представляет интерес совместная филогенетическая эволюция изменения содержания белка и масла в семенах сои (рис. 3).
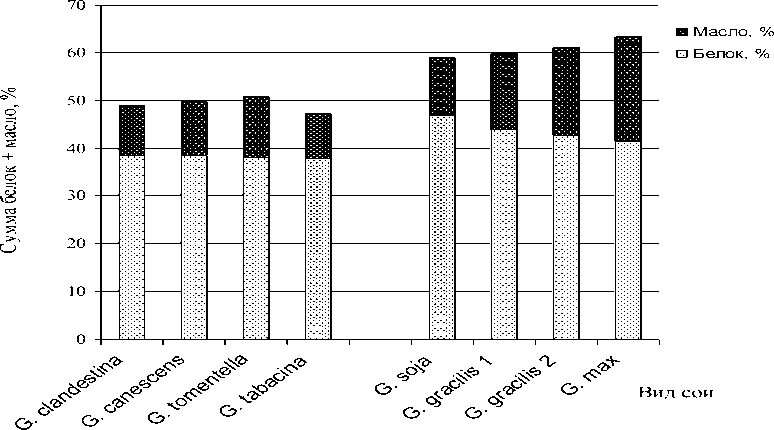
Рисунок 3 – Динамика совместной эволюции признаков содержания белка и масла в подроде Soja
На рис. 3 видно, что в подроде Soja увеличение масличности семян идет на фоне уменьшающегося содержания белка. Тем не менее, сумма этих показателей увеличивалась в процессе эволюции и у культурной сои достигла 60-62 %. При этом у видов подрода Glycine этот составной показатель не превышал 50-51 % уровень от массы семени.
Несмотря на увеличение показателя суммы белка и масла в филогенезе подрода Soja , дальнейший прогноз динамики этого показателя затруднителен. Причиной этому является практически полное отсутствие работ, посвященных изучению этого вопроса. Однако, по аналогии с эволюцией масличности семян, можно сделать предварительное заключение о проблематичности дальнейшего увеличения суммы белка и масла в роде Glycine .
Биохимический анализ показал, что дикорастущие виды сои отличались более низкими значениями трипсинингибирующей активности (ТИА), по сравнению с культивируемым видом G. max . (рис. 4). Более того, последовательное окультуривание сои сопровождалось заметным увеличением активности ингибиторов трипсина, достигая у отдельных форм культурной сои уровня 38 мг/г. Выявленная тенденция эволюции этого признака в подроде Soja в перспективе может привести к снижению питательной ценности продуктов и кормов из сои за счёт высокого уровня ТИА.
В настоящее время разработано множество технологических и биологических способов и приемов снижения активности ингибиторов трипсина. Однако попытки снизить активность этих антипитательных веществ до допустимого уровня путем, например термической обработки, приводят к значительному разрушению также и ценных компонентов зерна, в результате чего соя теряет свои преимущества как источник полноценных сбалансированных питательных веществ.
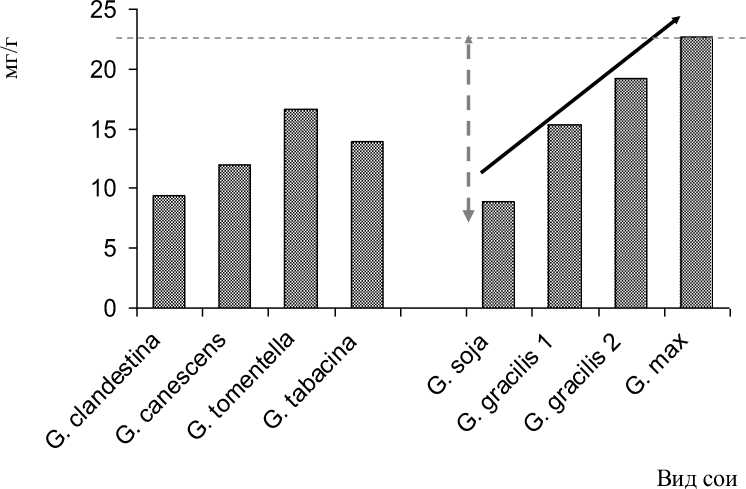
Рисунок 4 – Активность ингибиторов трипсина в роде Glycine и динамика филогенетической эволюции ТИА в подроде Soja тренд эволюции признака
^ _ направление возможного генетического улучшения признака верхний предел признака в исследуемом генофонде рода
Наиболее эффективным и экономически целесообразным является снижение этого показателя селекционными методами до биологически возможного уровня. Попытки создать низкоингибиторные сорта сои осуществлялись практически во всём мире. Однако дефицит доноров признака пониженной трипсинингибирующей активности в естественном генофонде культурной сои до последнего времени не позволял добиться положительных результатов.
Анализ данных, представленных на рис. 4, позволяет сделать вывод о полиморфизме признака трипсинингибирующей активности как в подроде Soja, так и в роде Glycine в целом.
Как следует из рис. 4, максимальными значениями ТИА отличались сорта вида G. max . Минимальным уровнем ТИА отличались формы G. soja , диапазон изменчивости которых в условиях 2001-2003 гг. составил от 5,96 до 12,17 мг/г. Сортообразцы сои изящной по этому показателю занимали промежуточное положение. Показатели ТИА у австралийских видов, в целом, также занимали промежуточное положение между уровнями ТИА уссурийской и культурной сои.
Полученные данные позволяют сделать благоприятный прогноз при развитии селекции на снижение ТИА до уровня дикорастущего предка – сои уссурийской ( G. soja ).
Одним из современных направлений в селекции масличных культур, в т.ч. и сои, является селекция на изменение жирно-кислотного состава масла. Разнонаправленные тре- бования различных отраслей промышленности вызывают необходимость создания сортов сои с различным жирно-кислотным составом масла.
Как следует из рис. 5, филогенетическая эволюция жирно-кислотного состава масла у видов сои в пределах подрода Soja шла в направлении снижения полиненасы-щенных кислот – линоленовой и линолевой и увеличения олеиновой кислоты.
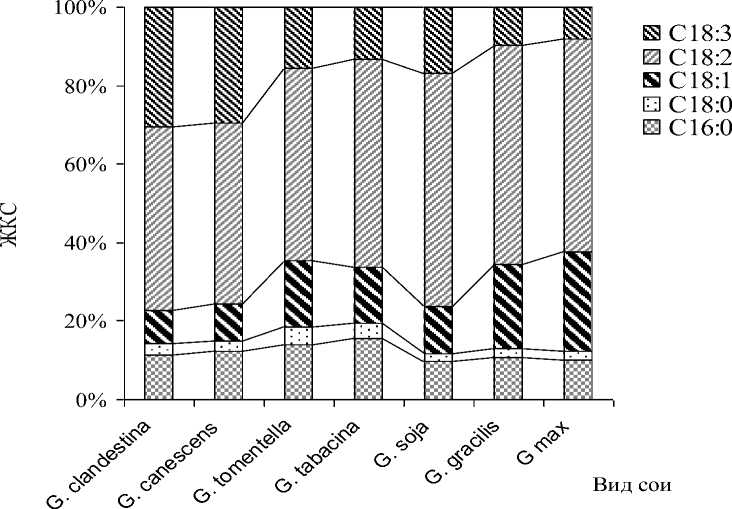
Рисунок 5 – Динамика эволюции жирно-кислотного состава масла в роде Glycine
При этом в наших исследованиях содержание в масле насыщенных пальмитиновой и стеариновой кислот в подроде Glycine варьировало в пределах 11,4-15,7 и 2,5-4,3 % соответственно. Доля этих кислот в масле видов подрода Soja практически не претерпела никаких изменений за всё время филогенеза рода Glycine . Австралийские виды сои отличались несколько большим содержанием насыщенных кислот, в среднем составляющим 11,4-15,7 % пальмитиновой кислоты и 2,5-4,8 % стеариновой кислоты.
Неодинаковый компонентный состав масла у различных видов сои предполагает возможность селекционно-генетического улучшения по жирно-кислотному составу, в первую очередь, по содержанию филогенетически лабильных олеиновой, линолевой и линоленовой кислот.
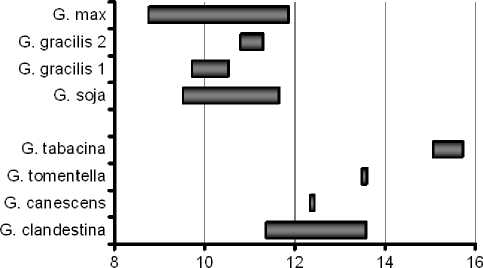
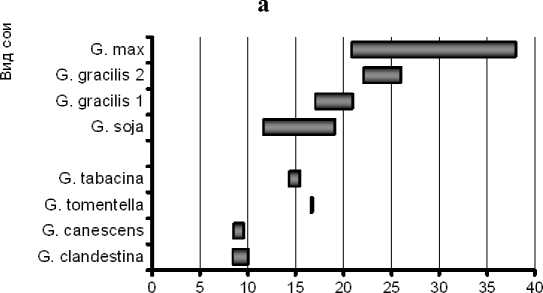
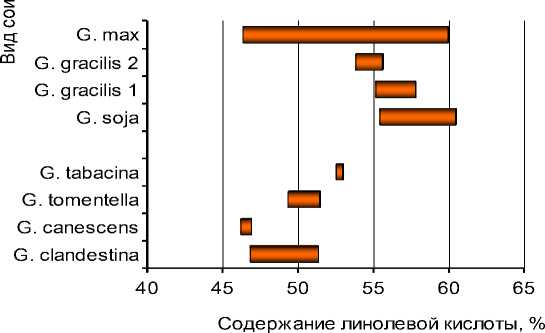
Подтверждением этому может служить наличие варьирования содержания основных жирных кислот в пределах вида (рис. 6).
Наличие в генофонде подрода Soja форм с повышенным до 60 % или пониженным до 47 % содержанием линолевой кислоты, определяет возможность успешной селекции сортов культурной сои в этих направлениях.
Содержание пальмитиновой кислоты, %
Содержание олеиновой кислоты, %
б
в
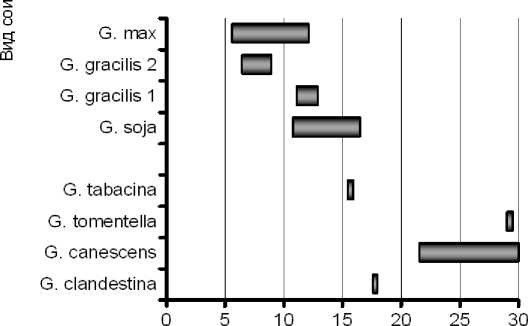
Содержание линоленовой кислоты, %
г
Рисунок 6 – Диапазоны варьирования 4-х жирных кислот сои у некоторых видов сои рода Glycine а – пальмитиновая кислота С16:0; б – олеиновая кислота С18:1;
в – линолевая кислота С18:2; г – линоленовая кислота С18:3.
Таким образом, в результате исследований полиморфизма биохимических признаков – содержания в семенах белка и масла, активности ингибиторов трипсина и жирно-кислотного состава масла у сортов культурной сои и 6 дикорастущих видов Китайского и Австралийского генетических центров. Наличие в гомологических рядах видов сои подрода Soja высокобелковых и низкоингибиторных форм открывает возможность выведения сортов культурной сои с высоким содержанием белка и пониженной трипсинингибирующей активностью.
