Изменение хемилюминесцентных показателей крови кроликов при тритиевом воздействии

Автор: Федотова А.С., Жигарев А.А., Макарская Г.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Зоотехния и ветеринария
Статья в выпуске: 4, 2026 года.
Бесплатный доступ
Цель исследования – оценить изменение хемилюминесцентных показателей крови кроликов при воздействии трития в субклинических дозах. Задачи: сформировать субклинические поглощенные дозы у кроликов; оценить время достижения и максимальную интенсивность хемилю-минесцентной реакции, суммарную продукцию первичных и вторичных радикалов кислорода в периферической крови кроликов при воздействии трития; рассчитать индекс активации хемилюминесцентной реакции. Работа выполнена на кроликах калифорнийской породы в период 2021–2023 гг. в Институте прикладной биотехнологии и ветеринарной медицины ФГБОУ ВО Красноярский ГАУ и лаборатории Международного научного центра исследований экстремальных состояний организма при Президиуме Федерального исследовательского центра КНЦ СО РАН. Для исследования сформированы поглощенные дозы: до 2,6 мГр; 3,3–4,2; 6,4–14,0; 16,6–21,4; 31,9; 50,9–52,2; 62,3–63,2; 92,6 и 355,6 мГр. Оценивали ключевые параметры продукции АФК: время достижения максимума при формировании пика хемилюминесцентной кривой; максимальную интенсивность пика генерации АФК; суммарную продукцию радикалов; индекс активации. Динамика суммарного количества активированных первичных АФК имела бимодальный характер с модой при дозах 3,3–4,2 и 62,3 мГр. При воздействии трития на организм кроликов в диапазоне от 2,6 до 62,3 мГр и при дозе 355,6 мГр увеличивалось количество спонтанных первичных радикалов кислорода относительно значений интактных групп. При воздействии трития в дозе 63 мГр увеличивалась спонтанная и активированная продукция первичных АФК и спонтанная генерация вторичных радикалов кислорода, при этом наблюдалось снижение ИА первичных и вторичных радикалов, что нами определяется как негативный фактор тритиевого воздействия. Установленные изменения подтверждают теорию о бимодальной зависимости показателей при воздействии радиации в малых дозах.
Периферическая кровь, кролики, свободные радикалы кислорода, хемилюминесценция, максимальная интенсивность реакции
Короткий адрес: https://sciup.org/140315519
IDR: 140315519 | УДК: 619:616.15:636.92 | DOI: 10.36718/1819-4036-2026-4-162-173
Changes in chemiluminescent blood parameters in rabbits upon tritium exposure
The aim of the study is to evaluate changes in chemiluminescent parameters of rabbit blood when ex-posed to subclinical doses of tritium. Objectives: to form subclinical absorbed doses in rabbits; to estimate the time to reach and the maximum intensity of the chemiluminescent reaction, the total production of pri-mary and secondary oxygen radicals in the peripheral blood of rabbits when exposed to tritium; to calculate the activation index of the chemiluminescent reaction. The work was carried out on Californian rabbits in the period 2021–2023 at the Institute of Applied Biotechnology and Veterinary Medicine of the Kras¬noyarsk State Agrarian University and the laboratory of the International Scientific Center for Research of Extreme States of the Body under the Presidium of the Federal Research Center of the Krasnoyarsk Scientific Center of the Siberian Branch of the Russian Academy of Sciences. The absorbed doses formed for the study were: up to 2.6 mGy; 3.3–4.2; 6.4–14.0; 16.6–21.4; 50.9–52.2; 62.3–63.2; 92.6 and 355.6 mGy. The key parameters of ROS production were assessed: time to reach the maximum during the formation of the chemiluminescence curve peak; maximum intensity of the ROS generation peak; total ra-dical production; activation index. The dynamics of the total number of activated primary ROS were bi-modal, with a mode at doses of 3.3–4.2 and 62.3 mGy. When rabbits were exposed to tritium in the range from 2.6 to 62.3 mGy and at a dose of 355.6 mGy, the number of spontaneous primary oxygen radicals increased relative to intact groups. When exposed to tritium at a dose of 63 mGy, spontaneous and activated production of primary ROS and spontaneous generation of secondary oxygen radicals increased, while a decrease in the IA of primary and secondary radicals was observed, which we define as a negative factor of tritium exposure. The observed changes confirm the theory of a bimodal relationship between parameters upon exposure to low doses of radiation.
Текст научной статьи Изменение хемилюминесцентных показателей крови кроликов при тритиевом воздействии
Введение. Составляющие биосферы на протяжении всей эволюции испытывают воздействие внешних факторов, одним из которых является ионизирующее излучение. На элементы биосферы постоянно действуют космическая радиация, бета-, гамма-излучение естественных радионуклидов, находящихся в горных породах, почве, воде, в атмосферном воздухе, в воздухе помещений и входящих в состав живых организмов, все перечисленные источники формируют среднегодовую поглощенную дозу от природных источников. Величина среднегодовой поглощенной дозы от природных источников для сельскохозяйственных животных различна на территории РФ и определяется географической широтой и высотой над уровнем моря. Три- тий (3H), его изотопы 1Н и 2Н относятся к аэрогенным природным изотопам, присутствуют во внутренней среде организма сельскохозяйственных животных и участвуют в формировании суммарной поглощенной природной дозы [1]. В докладе Научного комитета ООН по действию атомной радиации (2000 г). определено, что суммарное количество природного 3H составляет 1,3 · 1018 Бк. В биосфере 99 % от общего количества природного 3H находится в виде оксида трития (тритиевой воды (НТО)) - 3Н20 или Т2О и в таком виде участвует в глобальном круговороте воды [1]. Совокупность природных радионуклидов в объектах биосферы формирует естественный радиационный фон. Значение природной среднегодовой поглощенной дозы не вызывает в организме сельскохозяйственных животных патологических изменений и воспринимается системами организма как норма [2]. В формировании среднегодовой поглощенной дозы природные источники составляют 99,98 %, дополнительный вклад в среднегодовую поглощенную дозу продуктивных животных вносят антропогенные источники ионизирующего излучения.
Тритий (Т, 3H) – радиоактивный изотоп водорода с массовым числом 3, ядро изотопа представлено одним протоном и двумя нейтронами. Период полураспада (Т1/2) – 12,26 года, бета-излучатель с энергией β-частиц – 5,7 кэВ, максимальная энергия β-частиц – 18,591 кэВ, пробег β-частиц в воздухе – 4,5–6,0 мм, в тканях – около 1 мк, удельная активность – 3,6 · 1014 Бк/кг [3, 4]. 3H присутствует в природе как естественный и антропогенный изотоп. Природный 3H образуется в результате взаимодействия протонов и нейтронов вторичного космического излучения с атомами атмосферы (N, O 2 , Ar) [4, 5]. Согласно нормам агентства по охране окружающей среды, предельно допустимая концентрация (ПДК) для изотопа – 740 Бк/л, уровень вмешательства для поверхностных вод – 15 Бк/л, в странах Евросоюза ПДК трития в воде – 100 Бк/л, в России уровень вмешательства – 7600 Бк/л, ПДК 3H – 45 000 Бк/л [4]. С учетом периода полураспада общее количество 3H, выброшенного в биосферу с 1954 г. при испытании термоядерного оружия, составляло 1200 кг. В настоящее время содержание 3H равно 300 кг. Основная часть 3H сосредоточена в пресных водах и биосфере, концентрация 3H в дождевой воде увеличилась от 0,5–5,0 до 500 ТE [4, 6].
В настоящее время основным источником техногенного трития является атомная промышленность. Получение энергии в условиях термоядерных реакторов увеличит накопление трития в биосфере, ожидается, что выделение трития в термоядерном реакторе в 104–105 раз больше по сравнению с работой АЭС при эквивалентной мощности, таким образом, к 2100 г. равновесное содержание трития в биосфере Земли составит около 2,5 · 1022 Бк [1].
При выбросе в атмосферу 3H быстро мигрирует в объекты окружающей среды [4]. 3H в водоемы попадает в результате прямого сброса и обмена парами 3H атмосферы и поверхности водоема и с осадками [7]. После этого он включается в цепь миграции: вода, почва – расте- ние – животные – агропродукция. Количество трития в окружающей среде – основной фактор, определяющий его содержание в звеньях экологической системы. Вблизи радиационно-опасных объектов со временем 3H хорошо накапливается в объектах окружающей среды [3].
Тритий при попадании внутрь организма активно инкорпорируется в состав биологической ткани, вызывая мутагенные нарушения. При распаде образует гелий (Не) и β-излучение, низкой энергией, в связи с этим при внешнем облучении организма не представляет угрозы для организма. При внутреннем облучении (поступление аэрогенно, алиментарно) изотоп представляет серьезную опасность для здоровья животных, изотоп в химическом вступает в соединения с кислородом, серой, азотом, легко проникает через мембрану клетки, активно воздействует на органеллы, нарушает их функционирование [1].
Таким образом, содержание 3H в объектах окружающей среды в настоящее время достаточное, изотоп относится к группе биологически активных радионуклидов. 3H влияет на все уровни организации биологического организма. Необходимо усилить контроль за выбросом 3H в окружающую среду, проводить постоянный мониторинг компонентов агробиоценозов, подверженных загрязнению 3H. Необходимо оценить радиобиологические эффекты в организме сельскохозяйственных животных, возникающие при действии 3H в разных дозах.
Ранее авторами проведена работа по оценке степени воздействия низких доз трития: 16 мкГр, 31 и 47 мкГр на фагоцитарную активность клеток крови, гематологические параметры и образование свободных кислородных радикалов в венозной крови кроликов [8]. Оценена генерация активных форм кислорода при тритиевом воздействии в дозе 6,78 мГр на организм кроликов, определена эффективность применения радиопротектора «Гумат калия 80». Установлено, что радиопротекторное действие препарата «Гумат калия 80» обусловлено снижением суммарного количества спонтанных вторичных АФК и увеличением количества антиген активированных АФК. В заключении авторы рекомендуют при тритиевом воздействии качестве радиопротектора применение «Гумат калия 80» в дозе 10–4 г/л [9]. Исследование является продолжением работы по гранту РНФ 23-26-10018 «Прогнозирование реакции сельскохозяйственных животных на низ- коинтенсивную радиацию и применение радиопротекторов. Экспрессный биолюминесцентный скрининг радиобиологических эффектов».
Цель исследования – оценить изменение хемилюминесцентных показателей крови кроликов при воздействии трития в субклинических дозах.
Задачи: сформировать субклинические поглощенные дозы у кроликов; оценить время достижения и максимальную интенсивность хемилюминесцентной реакции, суммарную продукцию первичных и вторичных радикалов кислорода в периферической крови кроликов при воздействии трития; рассчитать индекс активации хемилюминесцентной реакции.
Материалы и методы. Исследование проведено в период 2021–2023 гг. в Институте прикладной биотехнологии и ветеринарной медицины ФГБУ ВО «Красноярский государственный аграрный университет» и лаборатории Международного научного центра исследований экстремальных состояний организма при Президиуме Федерального исследовательского центра КНЦ СО РАН.
Работа выполнена на кроликах калифорнийской породы (возраст (7 ± 0,3), (12 ± 0,3) мес.), созданы 9 опытных групп (10 животных в каждой группе), 3 интактных группы (10 голов), вес кроликов (4 ± 0,5) кг. Клиническое исследование проводили посредством общего клинического осмотра, термометрии, аускультации, исследования выполнены по общепринятой методике. Кролики опытных групп получали тритиевую воду, в результате в 9 опытных группах сформированы поглощенные дозы: до 2,6 мГр; 3,3– 4,2; 6,4–14,0; 16,6–21,4; 31,9; 50,9–52,2; 62,3– 63,2; 92,6 и 355,6 мГр. Интактная (контрольная) группа животных получала воду из централизованной системы водоснабжения. Кроликов всех групп подвергали клиническому обследованию, оценивали гематологические и хемилюминесцентные показатели периферической крови. Рацион кормления и система содержания кроликов всех групп не отличались, рацион состоял из полнорационного комбикорма ПК-90 и сена разнотравного. Кролики содержались в индивидуальных клетках в условиях зоофермы ИП-БиВМ ФГБОУ ВО «Красноярский ГАУ». Отбор проб крови осуществлялся у кроликов в утренние часы, отбирались образцы крови из краевой ушной вены в вакуумные пробирки с добавлением натрия гепарина.
Результаты и их обсуждение. Уровень свободно радикальных процессов в клетках периферической крови кроликов при субклиническом воздействии трития определен хемилюминесцентным (ХЛ) методом. Свободно радикальные процессы в клетках периферической крови кроликов оценивали по спонтанной и антиген активированной in vitro частицами латекса продукции активных форм кислорода (АФК). В работе оценивали ключевые параметры продукции АФК: время (Тmax, мин) достижения максимума при формировании пика ХЛ кривой; максимальную интенсивность (Imax, имп/с) пика генерации АФК; суммарную продукцию радикалов (S, млн имп. за 90 мин); индекс активации (ИА, усл. ед.) – отношение суммарной продукции АФК активированной и спонтанной ХЛ [10].
Время достижения максимума ХЛ (Тmax, мин). При воздействии трития в субклинических дозах время формирования пиков хемилюминесцентной реакции носило изменчивый характер. В работе установлено сокращение времени на формирование пика спонтанной генерации первичных АФК: в группе № 2 (3,3–4,2 мГр) уменьшение в 1,26 раза (P ≤ 0,01); в группе № 3 (6,4–14,0 мГр) – в 1,29 раза (P ≤ 0,001) по сравнению с интактным контролем № 3. Сокращение времени на формирование пика спонтанной генерации первичных радикалов в группе № 5 (31,9 мГр) – в 1,52 раза (P ≤ 0,01) по сравнению со интактным контролем № 2 и в группе № 8 (92,6 мГр) – в 4,72 раза (P ≤ 0,001) по отношению к интактному контролю № 3 (табл. 1). Установлено, что время формирования пика ХЛ кривой при спонтанном образовании первичных радикалов увеличивалось в группе № 4 (16,0– 21,4 мГр) в 1,47 раза (P ≤ 0,001); в группе № 7 (62,3 мГр) – в 1,67 раза (P ≤ 0,001) по сравнению с интактным контролем № 2 и№ 3 соответственно.
Время формирования пика первичных (лю-цигенин-зависимых) радикалов при стимуляции ХЛ-реакции латексом возрастало в группе № 4 (16,0–21,4 мГр) в 1,66 раза (P ≤ 0,001); в группе № 7 (62,3–63,2 мГр) – в 1,50 раза (P ≤ 0,001) относительно интактного контроля № 2 и № 3 соответственно. Установлено сокращение времени формирования пика ХЛ-реакции в группе № 3 (6,4–14,0 мГр) в 1,34 раза (P ≤ 0,01); в группе № 5 (31,9 мГр) – в 1,74 раза (P ≤ 0,001); в группе № 8 (92,6 мГр) – в 3,67 раза (P ≤ 0,001) по сравнению с контрольными значениями.
Аналогичная тенденция прослеживалась при генерации вторичных радикалов (люминол-зависимых). Время формирования пика при спонтанном образовании вторичных АФК сокращалось: в группе № 2 (дозы 3,3–4,2 мГр) – в 1,52 раза (P ≤ 0,001); в группе № 3 (6,4– 14,0 мГр) – в 1,66 раза (P ≤ 0,001) по сравнению с интактным контролем № 1. Время образования пика спонтанной генерации вторичных радикалов уменьшалось в группе № 6 (50,9– 52,2 мГр) – в 2,12 раза (P ≤ 0,01); в группе № 8 (92,6 мГр) – в 3,98 раза (P ≤ 0,001) относительно данных интактного контроля № 3. Увеличение времени формирования пика вторичных АФК отмечалось: в группе № 4 (16,0–21,4 мГр) – в 1,28 раза (P ≤ 0,01); в группе № 7 (62,3–
63,2 мГр) – в 2,88 раза (P ≤ 0,001); в группе № 9 (355,6 мГр) – в 1,6 раза (P ≤ 0,001) по сравнению с интактным контролем № 2 и № 3 соответственно. При антиген активированной продукции вторичных радикалов время формирования пика первичных АФК увеличивалось в группе № 4 (поглощенная доза 16,0–21,4 мГр) в 1,77 раза (P ≤ 0,001); в группе № 7 (62,3– 63,2 мГр) – в 1,50 раза (P ≤ 0,001) относительно интактного контроля № 2 и № 3 соответственно (табл. 1). Установлено сокращение времени формирования пиков ХЛ-реакции в группе № 3 (6,4–14,0 мГр) в 1,34 раза (P ≤ 0,01); в группе №5 (31,9 мГр) – в 1,74 раза (P ≤ 0,001); в группе № 8 (92,6 мГр) – в 3,67 раза (P ≤ 0,001) по сравнению с контрольными значениями.
Таблица 1
|
Группа (поглощенная доза) |
Первичные радикалы |
Вторичные радикалы |
||
|
Спонтанная ХЛ |
Активированная ХЛ |
Спонтанная ХЛ |
Активированная ХЛ |
|
|
Интакт. конт. № 1 |
50,65 ± 2,37 |
38,71 ± 2,19 |
42,47 ± 2,73 |
21,53 ± 0,75 |
|
№ 1 (до 2,6 мГр) |
46,51 ± 2,66 |
38,92 ± 1,44 |
42,38 ± 0,99 |
22,04 ± 0,72 |
|
№ 2 (3,3–4,2 мГр) |
40,35 ± 2,17** |
33,00 ± 3,11 |
27,90 ± 1,59*** |
21,05 ± 1,32 |
|
№ 3 (6,4–14,0 мГр) |
39,28 ± 2,02*** |
28,91 ± 2,28** |
25,59 ± 3,22*** |
11,63 ± 0,46** |
|
Интакт. конт. № 2 |
47,40 ± 3,30 |
34,16 ± 1,54 |
38,92 ± 1,29 |
19,40 ± 0,92 |
|
№ 4 (16,0–21,4 мГр) |
69,79 ± 4,10*** |
56,71 ± 2,24*** |
50,00 ± 3,29** |
38,14 ± 4,34*** |
|
№ 5 (31,9 мГр) |
31,13 ± 1,84** |
19,63 ± 1,28*** |
37,88 ± 1,48 |
16,25 ± 2,09 |
|
Интакт. конт. № 3 |
47,83 ± 2,27 |
40,33 ± 2,14 |
23,88 ± 1,74 |
19,48 ± 1,84 |
|
№ 6 (50,9–52,2 мГр) |
51,87 ± 3,08 |
36,80 ± 2,27 |
11,27 ± 3,72** |
20,60 ± 5,06 |
|
№ 7 (62,3–63,2 мГр) |
80,00 ± 2,02*** |
60,67 ± 2,42*** |
68,67 ± 2,13*** |
55,00 ± 2,50*** |
|
№ 8 (92,6 мГр) |
10,13 ± 1,31*** |
11,00 ± 1,16*** |
6,00 ± 0*** |
6,75 ± 0,41*** |
|
№ 9 (355,6 мГр) |
49,75 ± 2,39 |
40,75 ± 1,96 |
38,25 ± 2,12*** |
18,75 ± 1,78 |
Примечание: * – P ≤ 0,05; ** – P ≤ 0,01; *** – P ≤ 0,001 по отношению к интактному контролю.
Время формирования пика при ХЛ-реакции Peak formation time in CL reaction
Максимальная интенсивность ХЛ (Imax, имп/с). При воздействии субклинических доз трития выявлены изменения в максимальной интенсивности пиков спонтанной и активированной ХЛ-реакции клеток периферической крови (табл. 2). Установлено увеличение максимальной интенсивности спонтанной продукции первичных радикалов в группе № 1 (до 2,61 мГр) в 5,17 раза (P ≤ 0,001); в группе № 2 (3,3–4,2 мГр) – в 4,17 раза (P ≤ 0,001); в группе № 3 (6,4– 14,0 мГр) – в 4,79 раза (P ≤ 0,001) по сравнению с интактным контролем № 1 и в группе № 7 (62,3– 63,2 мГр) – в 1,78 раза (P ≤ 0,001) относительно значений интактного контроля № 3. Максимальная интенсивность ХЛ-реакции при спонтанной продукции люцигенин зависимых радикалов снижалась в группе № 5 (31,9 мГр) в 2,63 раза (P ≤ 0,001); в группе № 9 (355,6 мГр) – в 1,78 раза (P ≤ 0,001) относительно данных интактного контроля № 3.
При антиген активированной продукции лю-цигенин зависимых радикалов максимальная интенсивность реакции повышалась во всех группах. Увеличение максимальной интенсивности составило: в группе № 1 (до 2,61 мГр) – в 2,76 раза (P ≤ 0,001); в группе № 2 (3,3– 4,2 мГр) – в 2,23 раза (P ≤ 0,001); в группе № 3 (6,4–14,0 мГр) – в 2,14 раза (P ≤ 0,001) по сравнению с интактным контролем № 1. Максимальная интенсивность ХЛ-реакции повысилась в группе № 4 (16,0–21,4 мГр) – в 1,54 раза (P ≤ 0,01); в группе № 5 (31,9 мГр) – в 2,1 раз (P ≤ 0,001) по сравнению со интактным контролем № 2. Максимальная интенсивность ХЛ-реак-ции увеличилась в группе № 6 (50,9–52,2 мГр) – в 1,33 раза (P ≤ 0,01); в группе № 7 (62,3– 63,2 мГр) – в 1,21 раза (P ≤ 0,05); в группе № 8 (92,6 мГр) – в 1,51 раза (P ≤ 0,001); в группе № 9 (355,6 мГр) – в 1,22 раза (P ≤ 0,05) по сравнению с интактным контролем № 3 (табл. 2).
Таблица 2
Максимальная интенсивность ХЛ-реакции крови при действии субклинических доз трития, Imax, имп/с Maximum intensity of the CL reaction of blood under the influence
\of subclinical doses of tritium, Imax, imp/s
|
Группа (поглощенная доза) |
Первичные АФК |
Вторичные АФК |
||
|
Спонтанная ХЛ |
Активированная ХЛ |
Спонтанная ХЛ |
Активированная ХЛ |
|
|
Интакт. конт. № 1 |
101,13 ± 22,18 |
373,71 ± 36,46 |
200,88 ± 43,68 |
875,06 ± 43,07 |
|
№ 1 (до 2,6 мГр) |
522,44 ± 102,45*** |
1033,01 ± 76,92*** |
242,25 ± 16,96 |
793,43 ± 43,78 |
|
№ 2 (3,3–4,2 мГр) |
422,05 ± 37,83*** |
832,35 ± 60,27*** |
385,00 ± 20,52*** |
831,6 ± 61,69 |
|
№ 3 (6,4–14,0 мГр) |
484,84 ± 90,42*** |
798,72 ± 75,36*** |
282,91 ± 37,68 |
712,69 ± 63,38* |
|
Интакт. конт. № 2 |
401,28 ± 33,49 |
463,72 ± 50,59 |
421,84 ± 26,78 |
944,16 ± 47,13 |
|
№ 4 (16,0–21,4 мГр) |
324,93 ± 37,73 |
712,71 ± 55,74** |
254,71 ± 32,34** |
511,29 ± 75,29*** |
|
№ 5 (31,9 мГр) |
152,38 ± 26,56*** |
973, 00 ± 59,55*** |
169,88 ± 17,17*** |
1350,25 ± 59,99*** |
|
Интакт. конт. № 3 |
588,50 ± 31,40 |
892,28 ± 47,91 |
320,90 ± 26,36 |
827,23 ± 52,57 |
|
№ 6 (50,9–52,2 мГр) |
581,47 ± 64,76 |
1187,27 ± 81,81** |
232,07 ± 32,97* |
735,43 ± 52,57** |
|
№ 7 (62,3–63,2 мГр) |
1046,31 ± 100,93*** |
1079,31 ± 73,98* |
375,77 ± 37,20 |
946,54 ± 42,36 |
|
№ 8 (92,6 мГр) |
575,75 ± 34,35 |
1349,50 ± 82,57*** |
698,75 ± 27,55*** |
1784,88 ± 44,25*** |
|
№ 9 (355,6 мГр) |
331,00 ± 23,56*** |
1088,75 ± 68,06* |
392,75 ± 24,95 |
760,75 ± 47,02 |
Примечание: * – P ≤ 0,05; ** – P ≤ 0,01; *** – P ≤ 0,001 по отношению к интактному контролю.
Схожая картина выявлена в ХЛ-реакции при продукции вторичных АФК. Максимальная интенсивность ХЛ-реакции при спонтанной продукции вторичных радикалов кислорода снижалась в группе № 4 (16,0–21,4 мГр) в 1,66 раза (P ≤ 0,01); в группе № 5 (31,9 мГр) – в 2,48 раза (P ≤ 0,001); в группе № 6 (50,9–52,2 мГр ) – в 1,38 раза (P ≤ 0,05) относительно контрольных интактных групп. Однако в некоторых группах установлено увеличение максимальной интенсивности ХЛ-реакции при спонтанной продукции вторичных АФК. Выявлено усиление максимальной интенсивности в группе № 2 (3,3– 4,2 мГр) в 1,92 раза (P ≤ 0,001); в группе № 8 (92,6 мГр) – в 1,38 раза (P ≤ 0,001) относительно значений интактного контроля № 1 и № 3 соответственно. При антигенактивированной генерации 2 радикалов максимальная интенсивность продукции вторичных АФК снижалась в группе № 3 (6,4–14,0 мГр) в 1,23 раза (P ≤ 0,05); в группе № 4 (16,0–21,4 мГр) – в 1,85 раз (P ≤ 0,001); в группе № 6 (50,9– 52,2 мГр) – в 1,12 раза (P ≤ 0,01) по сравнению с интактными контролями. Выявлено увеличе- ние максимальной интенсивности продукции в группе № 5 (31,9 мГр) в 1,43 раза (P ≤ 0,001) и в группе № 8 (92,6 мГр) в 2,16 раза (P ≤ 0,001) относительно интактного контроля № 2 и № 3 соответственно.
Суммарная генерация АФК (S, млн имп. за 90 мин). При воздействии трития на организм кроликов в диапазоне от 2,6 до 62,3 мГр и при дозе 355,6 мГр увеличивается количество спонтанных первичных радикалов кислорода относительно значений интактных групп. Динамика суммарного количество активированных первичных АФК имеет бимодальный характер с модой при дозах 3,3–4,2 и 62,3 мГр (рис. 1).
В работе установлен рост суммарной спонтанной продукции первичных радикалов в периферической крови. При спонтанной продукции первичных радикалов установлено увеличение суммарной генерации АФК в группе № 1 в 1,67 раза (P ≤ 0,001); № 2 и № 3 – в 1,89 раза (P ≤ 0,001) соответственно относительно значений интактного контроля № 1 (рис. 1). Выявлено увеличение спонтанной продукции первичных радикалов при дозах 62,3–63,2 мГр (группа № 7) – в 1,73 раза (P ≤ 0,001) относительно интактного контроля № 3. Одновременно наблюдалось снижение спонтанной продукции первичных АФК в группе № 5 количество АФК уменьшилось в
2,4 раза (P ≤ 0,001), в группе № 8 в – 1,38 раза (P ≤ 0,001), в группе № 9 в – 1,57 раза (P ≤ 0,001) по сравнению с данными интактного контроля № 3 (рис. 1).
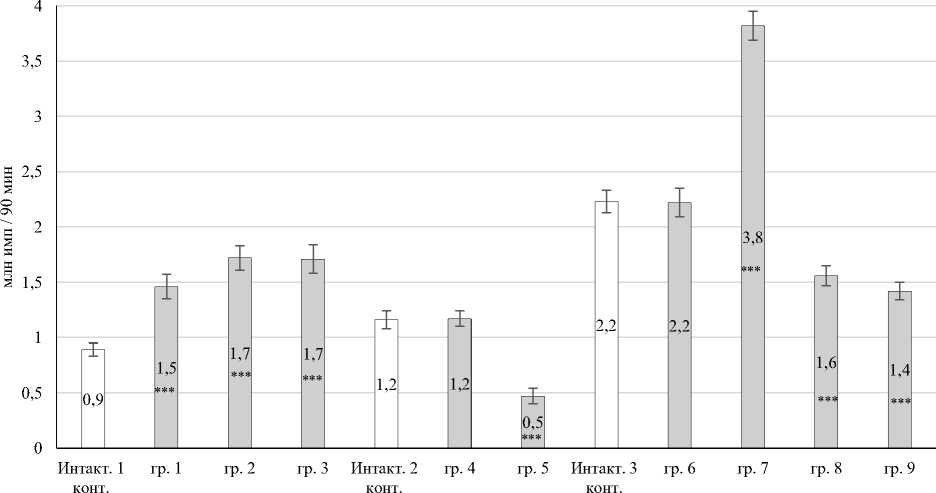
Рис. 1. Суммарная спонтанная продукция первичных АФК в периферической крови кроликов: * – Р < 0,05; ** – Р < 0,01; *** – Р < 0,001 по отношению к инактному контролю Total spontaneous production of primary ROS in the peripheral blood of rabbits:
* – P < 0.05; ** – P < 0.01; *** – P < 0.001 relative to the inactive control
Количество первичных антиген активированных АФК увеличивалось во всех группах (рис. 2). Суммарная продукция первичных антиген активированных радикалов возрастала в группе № 1 в 2,15 раза (P ≤ 0,001); в группе № 2 – в 2,17 раза (P ≤ 0,001); в группе № 3 – в 1,88 раза (P ≤ 0,001) относительно интактного контроля № 1. Число первичных антиген активированных радикалов кислорода увеличивалось в группе № 4 – в 1,51 раза (P ≤ 0,001); в группе № 5 – в 1,49 раза (P ≤ 0,01) по сравнению со интактным контролем № 2; в группе № 6 – в 1,22 раза (P ≤ 0,001); в группе № 7 – в 1,16 раза (P ≤ 0,01) и группе № 9 – в 1,24 раза (P ≤ 0,001) относительно значений интактного контроля № 3.
Выявлено снижение суммарного количества активированных первичных радикалов в группе № 8 в 1,13 раза (P ≤ 0,05) относительно значений интактного контроля № 3 (см. рис. 2).
Динамика суммарной спонтанной продукции вторичных (люминолзависимых) радикалов за 90 мин демонстрировала бимодальную зависимость от поглощенной дозы (рис. 3). Во второй опытной группе количество радикалов кислорода превышало в 1,95 раза (P ≤ 0,001) данные интактного контроля № 1. Суммарная спонтанная продукция вторичных АФК превышала в группе № 7 – в 1,22 раза (P ≤ 0,01); в группе № 9 – в 1,27 раза (P ≤ 0,01) значение интактного контроля № 3. Снижение суммарной спонтанной продукции установлено в группе № 4 – уменьшение в 1,7 раза (P ≤ 0,001); в группе № 5 – в 2,57 раза (P ≤ 0,001); в группе № 6 – в 1,74 раза (P ≤ 0,001); – в группе № 8 в 1,62 раза (P ≤ 0,001) относительно значений интактного контроля № 2 и № 3 (см. рис. 3).
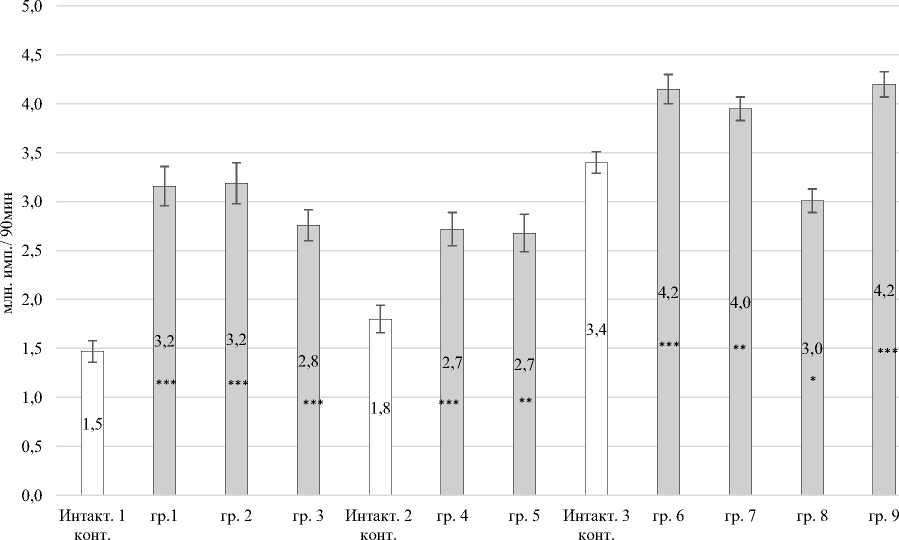
Рис. 2. Суммарная активированная продукция первичных АФК в периферической крови кроликов: * – Р < 0,05; ** – Р < 0,01; *** – Р < 0,001 по отношению к интактному контролю Total activated production of primary ROS in the peripheral blood of rabbits:
* – P < 0.05; ** – P < 0.01; *** – P < 0.001 relative to the intact control
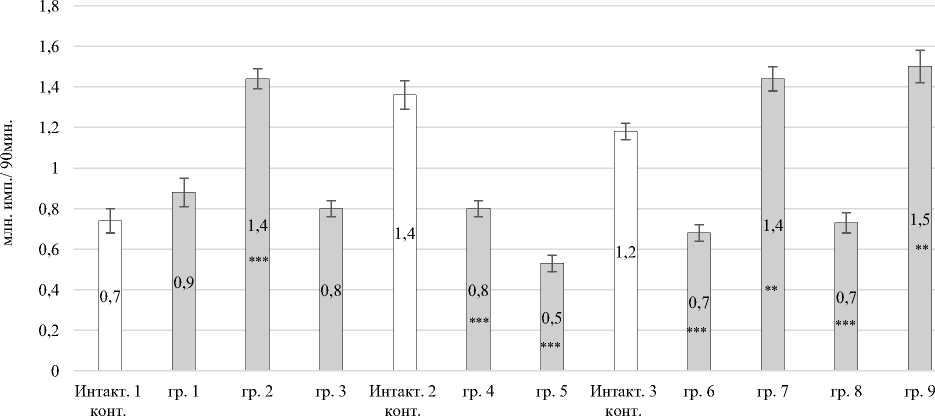
Рис. 3. Суммарная спонтанная продукция вторичных АФК в периферической крови кроликов:
* – Р < 0,05; ** – Р < 0,01; *** – Р < 0,001 по отношению к интактному контролю Total spontaneous production of secondary ROS in the peripheral blood of rabbits:
* – P < 0.05; ** – P < 0.01; *** – P < 0.001 relative to the intact control
Установлено достоверное снижение суммар- 1,44 раза (P ≤ 0,001); в группе № 8 – в 1,16 раза ной продукции активированных люминол зави- (P ≤ 0,01) относительно контрольных значений симых радикалов в группах № 3 и № 4 в (рис. 4).
гр. 3

1,6
***
2,5
Интакт. 2 конт.

1,7
***
гр. 4
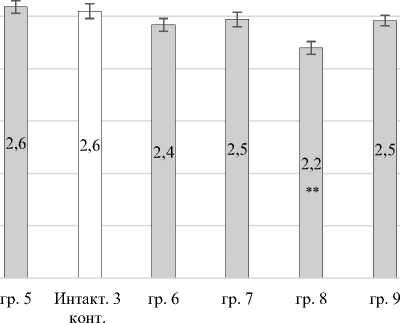
Рис. 4. Суммарная активированная продукция вторичных АФК в периферической крови кроликов: * – Р < 0,05; ** – Р < 0,01; *** – Р < 0,001 по отношению к итнактному контролю Total activated production of secondary ROS in the peripheral blood of rabbits:
* – P < 0.05; ** – P < 0.01; *** – P < 0.001 relative to the inactivated control
Индекс активации (ИА, усл. ед.). На основании данных по активированной и спонтанной продукции АФК рассчитан индекс активации ХЛ-реакции. Индекс активации люцигенин зависимых радикалов в группе № 1 превышал в 1,66 раза (P ≤ 0,001), во 2-й опытной группе – в 1,17 раза (P ≤ 0,05), значение интактной группы № 1 (рис. 5). В группе № 5 индекс активации превышал значение интактного контроля № 2 в 2,48 раза (P ≤ 0,001). В опытных группах № 6 и № 7 установлено снижение ИА относительно интактного контроля № 3 в 1,32 раза (P ≤ 0,01) и в 1,8 раз (P ≤ 0,001) соответственно. В группе № 9 установлено увеличение ИА в 1,19 раз (P ≤ 0,01) по сравнению с интактным контролем № 3.
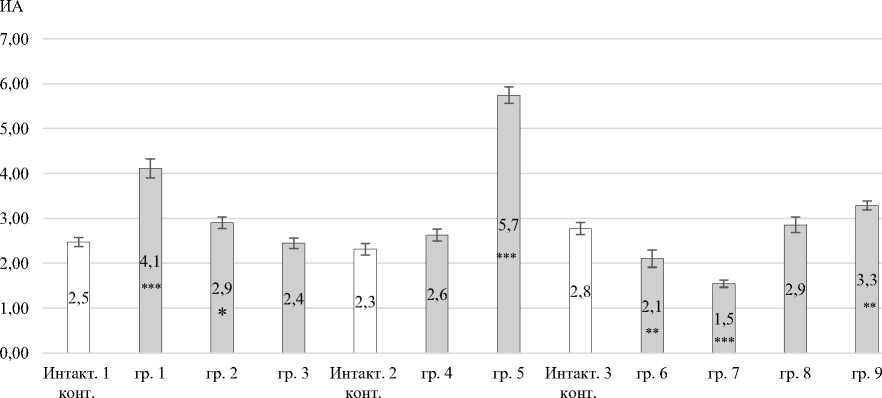
Рис. 5. Индекс активации первичных АФК клетками периферической крови кроликов: * – Р < 0,05; ** – Р < 0,01; *** Р < 0,001 по отношению к интактному контролю Index of primary ROS activation by rabbit peripheral blood cells:
* – P < 0.05; ** – P < 0.01; *** P < 0.001 relative to the intact control
Индекс активации люминол зависимых радикалов снижался в группе № 1 (до 2,61 мГр) в 1,31 раза (Р < 0,001); в группе № 2 (3,3–4,2 мГр) – в 1,42 раза (Р < 0,001); в группе № 3 – в 1,59 раза (Р < 0,001) относительно интактного контроля № 1 (рис. 6).
В группе № 4 (16,6–21,4 мГр) установлено уменьшение ИА в 1,17 раза (Р < 0,05) относительно интактного контроля № 2. Однако при дозе 31,9 мГр (группа № 5) выявлено увеличение индекса активации в 1,84 раза (Р < 0,001) относительно интактного контроля № 2. Погло- щенные дозы 50,9–92,6 мГр увеличивали индекс активации, в работе установлен рост индекса активации в группе № 6 в 2,37 раз (Р < 0,001); в группе № 7 – в 1,22 раза (Р < 0,05); в группе № 8 – в 1,91 раза (Р < 0,001) относительно интактного контроля № 3.
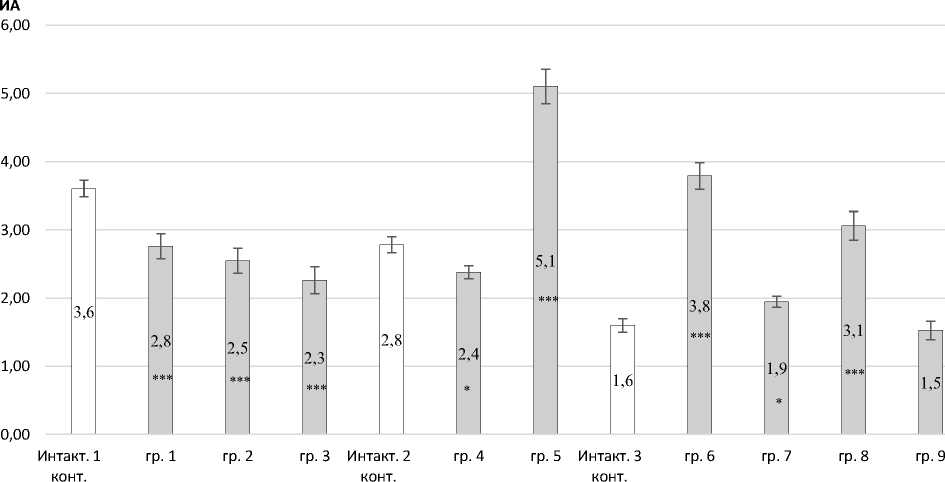
Рис. 6. Индекс активации вторичных АФК клетками периферической крови кроликов: * – Р < 0,05; ** – Р < 0,01; *** – Р < 0,001 по отношению к интактным контролям Index of secondary ROS activation by rabbit peripheral blood cells: * – P < 0.05; ** – P < 0.01; *** – P < 0.001 relative to intact controls
Воздействие радиации в дозе 355,6 мГр не приводило к изменениям индекса активации хемилюминесцентной реакции, значение находилось в диапазоне интактной группы № 3. Изменение индекса активации носило бимодальный характер, как и в других показателях.
Заключение. В хемилюминесцентных характеристиках крови кроликов, полученной после воздействия трития, замечена нестабильная динамика хемилюминесцентных показателей: времени достижения максимума, максимальной интенсивности, суммарной генерации АФК, индекса активации первичных и вторичных радикалов. Такие изменения подтверждают теорию о бимодальной зависимости показателей при воздействии радиации в малых дозах.
Динамика суммарного количества активированных первичных АФК имеет бимодальный характер с модой при дозах 3,3–4,2 и 62,3 мГр. При воздействии трития на организм кроликов в диапазоне от 2,6 до 62,3 мГр и при дозе 355,6 мГр увеличивается количество спонтанных первичных радикалов кислорода относительно значений интактных групп. Установлено, что при воздействии трития в дозе 63 мГр увеличивалось спонтанная и активированная продукция первичных АФК и спонтанная генерация вторичных радикалов кислорода, при этом наблюдалось снижение ИА первичных и вторичных радикалов, что нами определяется как негативный фактор тритиевого воздействия.
