Изменение пространственной структуры популяции разноцветной ящурки Eremias arguta Deserti (Gmelin, 1789) в полупустынях Астраханской области
 в полупустынях Астраханской области")
Автор: Полынова Галина Вячеславовна, Мишустин Станислав Сергеевич
Журнал: Принципы экологии @ecopri
Рубрика: Оригинальные исследования
Статья в выпуске: 2 (36), 2020 года.
Бесплатный доступ
Материалы по пространственной структуре разноцветной ящурки Eremias arguta deserti (Gmelin, 1789) собраны в полевые сезона 2017-2019 гг. в песчаных полупустынях Астраханской области. Трехлетние исследования на уровне внутрипопуляционной группировки показывают поэтапное сокращение численности популяции от сезона к сезону. В брачный сезон из системы перекрывающихся участков самцов и самок (2017 г.) пространственная структура группировки сначала трансформируется в систему изолированных участков половых партнеров (2018 г.), а затем в малочисленную группировку изолированных участков самок (2019 г.). Это происходит из-за того, что ящерицы рассредоточиваются на уменьшающихся по площади присущих им открытых биотопах. Уменьшение характерно и для кочующей части поселения, что подтверждает общее снижение численности популяции на окружающей территории. Группировка постепенно перестает размножаться, о чем свидетельствует отсутствие неполовозрелых особей во все весенние сезоны и снижение численности сеголеток в конце лета 2018 г. по сравнению с этим же периодом 2017 г. Основной причиной описанного процесса является изменение характерного для вида биоценоза - зарастание песчаных массивов, отмеченное в последние годы на многих территориях пустынных и полупустынных экосистем. Наблюдения показывают, что негативными последствиями зарастания биотопа являются ухудшения условий для передвижения и коммуникации. Измерение проективного покрытия и видового разнообразия фитоценоза, проведенное на пробных геоботанических площадках в 2011, 2014 и 2017 гг., говорит об остепнении территории. Основной причиной остепнения является увеличение общей суммы осадков в течение последнего десятилетия с 2010 по 2019 г. Последнее подтверждается положительной корреляцией увеличения проективного покрытия и общей суммы осадков за этот период.
Пространственная структура, популяция, разноцветная ящурка eremias arguta deserti, общее проективное покрытие, зарастание песков
Короткий адрес: https://sciup.org/147231299
IDR: 147231299 | УДК: 591.525
Changes in the spatial structure of the Eremias arguta Deserti population in semi-deserts of the Astrakhan region
Materials on the spatial structure of the lizard Eremias arguta deserti (Gmelin, 1789) were collected in the sandy semi-deserts of the Astrakhan region during the 2017-2019 field seasons. Three-year studies at the level of intra-population grouping show a gradual reduction in the population size from season to season. During the mating season, the spatial structure of the grouping is first transformed from a system of overlapping individual plots of males and females (2017) into a system of isolated territories of sexual partners (2018), and then into a small grouping of isolated home ranges of females (2019). This is due to the fact that lizards are dispersed in the area of decreasing inherent open biotopes. The decrease is also typical for the nomadic part of the grouping, which confirms the general decline in the population in the surrounding area. The group is gradually ceasing to reproduce, as evidenced by the absence of immature individuals in all spring seasons and a decrease in the number of current year’s young at the end of summer 2018 compared to the same period in 2017. The main reason for the described process of population degradation is a change in the biotope characteristic of the species - the overgrowth of sand massifs, which has been observed in recent years in many areas of desert and semi-desert ecosystems. Observations show that the negative consequences of biotope overgrowth are the deterioration of conditions for movement and communication. The measurement of the projective cover and plant species diversity of phytocenosis, carried out on sample geobotanical plots in 2011, 2014 and 2017, indicates that the territory is being transformed into steppe. The main reason for steppification is the increase in total precipitation over the past decade from 2010 to 2019. The latter is confirmed by a positive correlation between the increase in the projective cover and the total amount of precipitation over this period.
Текст научной статьи Изменение пространственной структуры популяции разноцветной ящурки Eremias arguta Deserti (Gmelin, 1789) в полупустынях Астраханской области
У11ИВЕРСИТЕТ
Изучение пространственной структуры является одним из наиболее актуальных направлений в популяционной экологии позвоночных животных. Именно эта составляющая популяционной структуры служит, с одной стороны, основой всех форм нормальной жизнедеятельности вида, а с другой – самым лабильным и действенным механизмом приспособления к меняющимся условиям окружающей среды. Пространственная структура популяции объединяет широкий спектр особенностей использования территории и является показателем благополучия вида (Шилов, 1977).
Одним из наиболее удобных объектов исследований в этой области служат открыто живущие виды пустынных и полупустынных экосистем, в частности ящерицы. Их изучение становится все более насущной задачей в связи с зарастанием песков, которое встречается повсеместно на аридных территориях и влияет на популяции рептилий. В русле изучения динамики герпетокомплексов песчаных полупустынь Астраханской области (Полынова и др., 2019) одним из основных объектов наших исследований оказалась разноцветная ящурка Eremias arguta deserti (Gmelin, 1789) – фоновый вид соответствующих экосистем этого региона.
Численность разноцветной ящурки в разных частях ареала и различных биотопах колеблется в широком диапазоне от 1.2–3 до 550–640 ос/га (Разноцветная ящурка, 1993). Географически близкие материалы по Калмыкии 70-х – начала 80-х гг. прошлого века показывают относительную стабильность плотности населения ящурки в большинстве стаций (Бадмаева, 1983). В Астраханской области на данный момент происходит значительное сокращение численности вида.
Выяснение причин снижения численности разноцветной ящурки и изучение пространственной структуры как механизма приспособления к меняющимся условиям среды – цель нашего исследования. Объектом исследования является внутрипопуля-ционная группировка – уровень, на котором проходят начальные процессы авторегуляции в популяции.
Материалы
Материалы по пространственной структуре разноцветной ящурки собраны в течение трех полевых сезонов: в первые две декады мая 2017, 2018 гг. и в первой половине мая 2019 г. Вне сезона размножения – с 27 авгу- ста по 16 сентября 2017 г. и с 19 по 29 августа 2018 г. Место исследований – окрестности поселка Досанг Красноярского района Астраханской области (N 46° 54'08.7264" E 47° 54' 52.5312").
Выбор поселения вида стал результатом маршрутного обследования территории вокруг поселка (~7 км2). Выбранное поселение соответствует уровню внутрипопуляцион-ной группировки или элементарной популяции (Наумов, 1963; Шилов, 1977) и расположено на отдельном участке полузакреплен-ного песка площадью 0.4 га. Фитоценоз – джузгуно-полынное сообщество ( Calligonum aphyllum Litv. и Artemisia arenaria DC.) с наличием признаков сукцессии в сторону степного сообщества (Полынова и др., 2019).
Во время исследований на территории поселения отловлены, промерены и помечены все встреченные особи разноцветной ящурки: в мае и сентябре 2017 г. – 76 и 25 особей, в мае и конце августа 2018 г. – 38 и 14 особей, в мае 2019 г. – 12 ящериц.
Статья не содержит сведений по размерам индивидуальных участков, поскольку цель работы не требует обсуждения этого параметра пространственной структуры. Кроме того, представленные материалы в большинстве случаев характеризуют только центры активности участков обитания. У данного вида центры активности – это знакомые естественные убежища: участок с кустами и норами под ними. Центры активности легко выявляются за небольшой период наблюдений и дают представление о плотности поселения.
Методы
В работе использовался набор методов, подробно описанный нами в ранее опубликованных исследованиях (Полынова, Бажи-нова, 2012), поэтому здесь приведен лишь их краткий перечень.
Маркировку производили временной (спиртовой маркер) и постоянной (отрезание кончиков фаланг пальцев по классической схеме) меткой. У меченых животных измеряли длину тела и хвоста (мм). Описание пространственной структуры осуществляли на основе картирования встреч и перемещений, построения карт индивидуальных участков по методу выпуклого многоугольника, методов тропления и осторожного преследования. Обработка полевых данных проходила в программе Adobe llustrator (для визуализации участка). На основе обработанных материалов выявлена оседлая и мигрирующая части популяции. Характеристика численности дана в ос/га, а оценка величины потока мигрантов – в ос/сутки. Проведено геоботаническое исследование динамики фитоценоза (Полынова и др., 2019). Оценка зависимости экологических показателей дана на основе коэффициента корреляции Пирсона.
Результаты
Согласно опубликованным сведениям общий характер пространственного распределения популяций разноцветной ящурки неравномерен. На Прикаспийской низменности размещение мозаичное (Табачишин и др., 2006), а в Центральном Предкавказье и Харьковской области - ленточное (Разноцветная ящурка, 1993). В нашем случае поселение представляет элемент мозаичного распределения, типичного для региона.
Трехлетние наблюдения, проведенные на уровне внутрипопуляционной группировки, свидетельствуют о направленном снижении численности вида. Значительно уменьшается как оседлая часть поселения, так и поток мигрантов (табл. 1).
Таблица 1. Динамика плотности населения группировки разноцветной ящурки
Eremias arguta deserti , май 2017-2019 гг.
|
Сезон |
Общая плотность, ос/га |
Плотность оседлых особей, ос/га |
Поток мигрантов, ос/сутки |
|
Май 2017 |
190 |
85 |
2.1 |
|
Май 2018 |
95 |
17.5 |
1.5 |
|
Май 2019 |
30 |
7.5 |
0.5 |
Процесс затрагивает не только количественную, но и качественную характеристику группировки: происходят принципиальные перемены в пространственной структуре и половозрастном составе.
Разноцветная ящурка, как большинство ящериц (Stamps, 1977; Vitt et al., 1974 и др.), относится к животным с интенсивным типом использования территории. Животные образуют систему групп перекрывающихся участков, размер которых колеблется в широком диапазоне (Тертышников, 1970). Главной функцией подобной пространственной структуры в сезон размножения является поддержание достаточного уровня контактов между брачными партнерами. Такую пространственную структуру группировки мы наблюдаем у разноцветной ящурки весной 2017 г. Это система перекрывающихся участков брачных партнеров (рис. 1) при соотношении половых групп 1 : 1.1 (16 самцов : 18 самок).
Система участков самок объединяет между собой всю внутрипопуляционную группировку. Неполовозрелых особей в группировке нет. Поток мигрантов, проходящий по территории поселения (см. табл. 1), также состоит только из половозрелых ящериц (18 самцов, 24 самки). Под потоком мигрантов мы понимаем не направленно движущихся животных, а особей, широко кочующих по территории популяции и не имеющих в дан- ном месте постоянного участка.
Весной 2018 г. пространственная структура поселения разноцветной ящурки существенно меняется. Оседлая часть группировки также состоит из самцов и самок (2 и 5 соответственно), но уменьшается в 4.8 раза (см. табл. 1). Кроме того, участки половых партнеров располагаются изолированно (рис. 2). Подобная пространственная структура гораздо меньше способствует контакту брачных партнеров. Как и весной предыдущего года, неполовозрелых ящериц среди оседлых особей нет. В небольшом числе они встречаются среди кочующих животных, составляя 10 % от общего числа мигрантов (11 самцов, 17 самок, 3 неполовозрелые особи). Поток мигрантов также снижается (см. табл. 1), что подтверждает снижение численности популяции в целом.
Весной 2019 г. от группировки разноцветной ящурки на данной территории остается 3 изолированных участка самок (рис. 3). Оседлых самцов нет. При таком составе функцию поддержания достаточного уровня брачных контактов пространственная структура, очевидно, уже не выполняет. Животных значительно меньше и в кочующей части поселения (табл. 1), что является показателем продолжающегося снижения общей численности популяции. Неполовозрелые ящерицы на территории поселения по-прежнему отсутствуют.
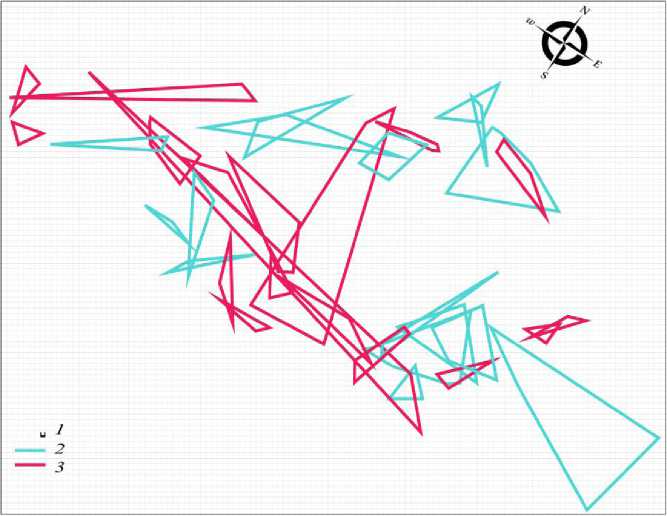
Рис. 1. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti , май 2017 г. 1 – размер ячейки 1 м, 2 – граница участка самца, 3 – граница участка самки Fig. 1. Spatial structure of intra-population group of the stepperunner Eremias arguta deserti , May 2017. 1 – size of cell is 1 m, 2 – boundary of the male area, 3 – boundary of the female area
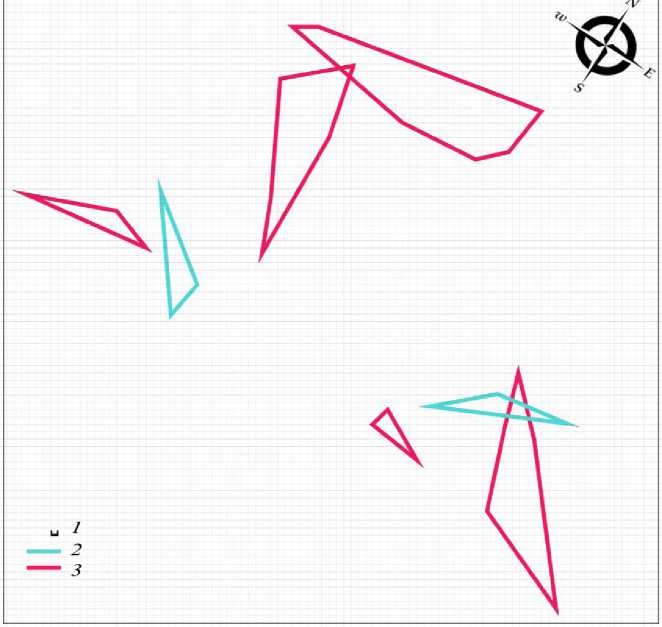
Рис. 2. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti , май 2018 г. 1 – размер ячейки 1 м, 2 – граница участка самца, 3 – граница участка самки Fig. 2. Spatial structure of intra-population group of the stepperunner Eremias arguta deserti , May 2018. 1 – size of cell is 1 m, 2 – boundary of the male area, 3 – boundary of the female area
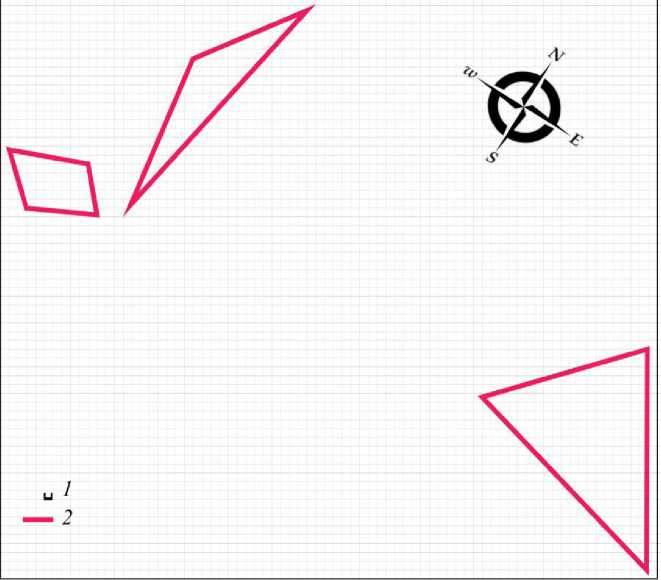
Рис. 3. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti, май 2019 г. 1 – размер ячейки 1 м, 2 – граница участка самки
Fig. 3. Spatial structure of intra-population group of stepperunner Eremias arguta deserti , May 2019. 1 – size of cell is 1 m, 2 – boundaryof the female area
Показателем успешности брачного сезона сокращающейся группировки служит выход сеголеток в конце лета – начале осени. В 2017 г. на территории группировки мы встречаем 18 сеголеток, а в 2018 г. – 10, т. е. в пересчете на плотность населения – 45 и 25 ос/ га соответственно. Снижение численности и в этой возрастной группе очевидно.
Основной причиной уменьшения численности вида, на наш взгляд, являются изменения характерного биотопа. По нашим наблюдениям, которые проходили в полевые сезоны 2010–2014 и 2017–2019 гг., происходит зарастание местообитания внутрипопу-ляционной группировки и примыкающей к ней территории популяции. В упрощенном варианте геоботанического исследования использованы два параметра: видовое разнообразие фитоценоза и общее проективное покрытие травяного яруса. Проведенное в 2011, 2014 и 2017 гг. геоботаническое описание площадок достоверно показывает, что за рассмотренный период слабо закрепленная территория превращается в полузакреплен-ную, полузакрепленная – в закрепленную, а общее проективное покрытие травяного яруса закрепленного участка увеличивается в 1.6 раза (Полынова и др., 2019). В 2018 и 2019 гг. во флоре к 13 постоянно встречаю- щимся видам прибавляется еще 10 новых однолетников и двулетников (Мишустин, Полынова, 2019а, б). С 2011 г. мы также наблюдаем за распространением колосняка гигантского Leymus racemosus (Lam.) Tzvelev – основного участника закрепления песчаных экосистем (Цвелев, 1976). При геоботаническом описании территории весной 2011 г. растение фиксируется на 1/4 всех заложенных площадок, весной 2014 – на 1/3, а весной 2017 г. колосняк присутствует уже на половине площадок. Сукцессионный процесс приводит к изменению джузгуно-полынного сообщества на джузгуно-разнотравное, т. е. идет в сторону остепнения биоценоза (Полы-нова и др., 2019).
Анализ динамики основных климатических факторов исследуемой территории показывает, что зарастание песков связано с увеличением общей суммы осадков ( Архив погоды в Досанге, 2020 ). Результат расчета линейного коэффициента корреляции Пирсона свидетельствует ( r = 0.82; p ≤ 0.01), что в течение последнего десятилетия увеличение общего проективного покрытия растительного яруса имеет сильную положительную зависимость от увеличения общей суммы осадков (рис. 4).
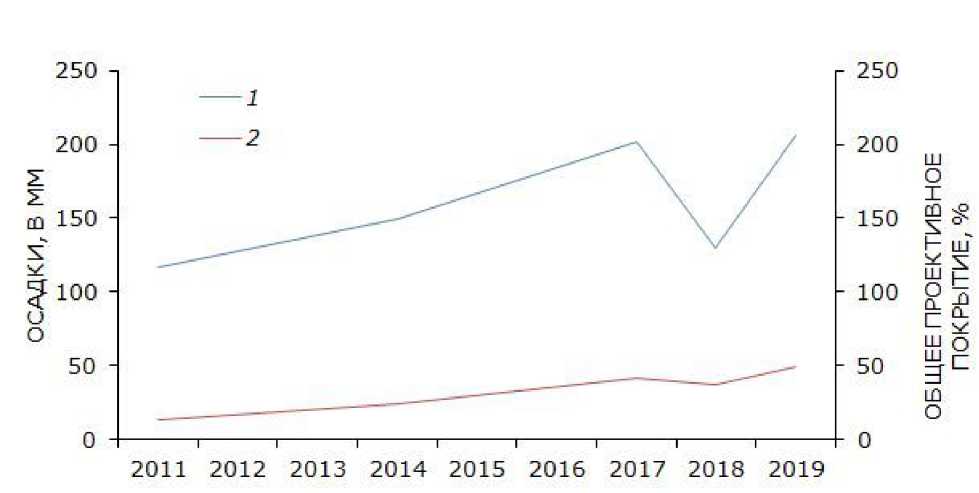
Рис. 4. Многолетняя динамика общей суммы осадков и проективного покрытия. 1 – общая сумма осадков (мм), 2 – общее проективное покрытие травяного яруса (%)
Fig. 4. Long-term dynamics of the total amount of precipitation and total projective cover of vegetation layer. 1 – total amount of precipitation (mm); 2 – total projective cover of vegetation layer (%)
Обсуждение
Снижение численности многих видов рептилий является широко распространенным явлением на зарастающих территориях песчаных пустынь и полупустынь нашей страны и сопредельных государств: Поволжья (Табачишин и др., 2006), Казахстана (Сараев, Пестов, 2010), Восточного Предкавказья (Лотиев, Батхиев, 2019), Узбекистана (Бондаренко и др., 2010). Подобный процесс отмечен и в условиях зарастания биотопов с каменистым грунтом (Galan, 2004). Процесс зарастания зависит прежде всего от климатических условий (Колесниченко, 1971). Наши материалы показывают его положительную корреляцию с увеличением общей суммы осадков за последнее десятилетие. Закреплению песков также способствует повсеместное сокращение поголовья скота и исчезновение диких копытных, отмеченное и для исследованной территории.
Изменение характерного биотопа негативно влияет прежде всего на виды псаммофилы (Полынова и др., 2019), также оказывая отрицательное воздействие на экологически пластичную разноцветную ящурку. Разноцветная ящурка относится к эвриби-онтным видам, однако имеет определенные стации предпочтения в разных зонах, что приводит к сложной картине биотопиче-ского распределения (Разноцветная ящурка, 1993). Выбор биотопа у разноцветной ящур- ки зависит от целого ряда факторов. Важную роль играет освещенность, что определяется структурой растительного покрова. На песчаных почвах предпочитает горизонтальные поверхности и участки с относительно разреженным травостоем (Котенко, 1983). Высокая численность ящерицы установлена в биотопах с проективным покрытием от нескольких до 30–40 % (Разноцветная ящурка, 1993). По нашим наблюдениям, негативными последствиями зарастания биотопа для вида являются ухудшения условий передвижения и коммуникации. Наиболее предпочитаемые стации вида имеют низкую влажность, песчаный или каменисто-песчаный грунт, высокую освещенность, возможность передвигаться и достаточное количество убежищ (Разноцветная ящурка, 1993).
На территории исследований вид селится на относительно открытых участках, образуя на них систему внутрипопуляционных группировок, пространственная и половозрастная структура которых реагирует на зарастание биотопа. В начале исследований, весной 2017 г., мы наблюдаем на такой открытой территории многочисленную группировку. Значительное перекрывание участков самцов и самок и их соотношение, близкое к 1 : 1, создают основу для процесса размножения. Подобное соотношение полов характерно для популяций разноцветной ящур-ки из разных частей ареала (Разноцветная ящурка, 1993) и свойственно многим видам ящериц в сезон размножения (Alcala, Brown, 1967; Bustard, 1970; Ballinger, 1973 и др.).
В следующий брачный сезон 2018 г. группировка продолжает свое существование на той же территории, адаптируясь к изменившимся условиям. Число оседлых животных резко уменьшается. Стремясь занять пространство с разреженной растительностью, животные образуют структуру с почти не перекрывающимися индивидуальными участками. Контакты брачных партнеров в этой ситуации становятся более редкими, что снижает эффективность процесса размножения. В результате численность сеголеток в конце августа уменьшается по сравнению с предыдущим годом в 1.8 раза. Снижение потока мигрантов свидетельствует о падении численности популяции в целом.
В весенний сезон 2019 г. способной к размножению группировки на территории поселения нет. Оседлы только немногочисленные изолированно живущие самки, занявшие участки с разреженной растительностью. Неоседлая часть поселения продолжает сокращаться. Популяция постепенно перестает размножаться на данной территории, что подтверждается отсутствием неполовозрелых особей во все весенние сезоны и снижением численности сеголеток в 2018 г. по сравнению с 2017 г.
Таким образом, на зарастание характерного биотопа группировка отвечает поэтапным снижением численности за счет постепенного сокращения размножения в измененных условиях. Снижение численности вида на данной территории подтверждено и наблюдениями наших коллег (Литвинов Н. А., устное сообщение).
Заключение
-
1. Трехлетнее исследование пространственной структуры разноцветной ящур-ки Eremias arguta deserti (Gmelin, 1789) показывает направленное сокращение численности популяции, которое наглядно проявляется на уровне внутрипопуля-ционной группировки.
-
2. Пространственная структура группировки в брачный сезон упрощается от сезона к сезону. Из системы перекрывающихся участков самцов и самок она сначала трансформируется в систему изолированных участков половых партнеров, а затем в малочисленную группировку изолированных участков самок. Это свидетельствует о том, что размножение на данной территории постепенно сокращается. Уменьшение мигрирующей части поселения подтверждает общее сокращение численности популяции.
-
3. Основной причиной описанного процесса является изменение характерного биоценоза: происходит увеличение общего проективного покрытия травяного яруса, разрастание закрепляющих пески видов растений и общее остепне-ние растительных сообществ. Протекающие в экосистеме изменения связаны с увеличением общей суммы осадков на данной территории за последнее десятилетие. Негативными последствиями зарастания биотопа для вида являются ухудшения условий передвижения и коммуникации.
Список литературы Изменение пространственной структуры популяции разноцветной ящурки Eremias arguta Deserti (Gmelin, 1789) в полупустынях Астраханской области
- Архив погоды в Досанге . URL: l^ttps://rp5.m/Архив погоды в Досанге (дата обращения: 23.01.2020).
- Бадмаева В. И. Ящерицы Калмыкии : Дис. ... канд. биол. наук. Киев, 1983. 227 с.
- Бондаренко Д. А., Перегонцев Е. А., Абдуназаров Б. Б., Сударев В. О. О современном состоянии гер-петофауны песчаных массивов Ферганской долины (Узбекистан) // Герпетологические исследования в Казахстане и сопредельных странах: Сборник научных статей / Под ред. Т. Н. Дуйсебаевой. Алматы: АСБК СОПК, 2010. С. 246-248.
- Котенко Т. И. Пресмыкающиеся левобережной степи Украины : Дис. ... канд. биол. наук. Киев, 1983. 555 с.
- Лотиев К. Ю., Батхиев А. М. О деградации туранского герпетофаунистического комплекса в Терском песчаном массиве (Восточное Предкавказье) // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 115-128. DOI: 10.21685/2307-9150-20192-12.
- Мишустин С. С., Полынова Г. В. Влияние абиотических факторов на популяции ящериц в условиях Астраханских полупустынь // Современная герпетология: проблемы и пути их решения. СПб.: Зоологический институт РАН, 2019а. С. 61.
- Мишустин С. С., Полынова Г. В. Динамика фитоценозов в полупустынях юго-восточной части Нижнего Поволжья // Вестник института ИКИАТ. 2019б. № 1 (38). С. 10-11. DOI: 10.24411/20717830-2019-10002.
- Наумов Н. П. Экология животных . М.: Высшая школа, 1963. 618 с.
- Полынова Г. В., Бажинова А. В. Учет особенностей активности при оценке численности популяции круглоголовки-вертихвостки (Phrynocephalus guttatus guttatus) // Зоологический журнал. 2012. Т. 91. № 11. С. 1411-1414.
- Полынова Г. В., Мишустин С. С., Полынова О. Е. Динамика герпетокомплекса песчаных пустынь Астраханской области // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 150-163. DOI: 10.21685/2307-9150-2019-2-15.
- Разноцветная ящурка : Коллективная монография / Под ред. Н. Н. Щербака. Киев: Наукова думка, 1993. 240 с.
- Сараев Ф. А., Пестов М. В. К кадастру рептилий Северного и Северо-Восточного Прикаспия // Герпетологические исследования в Казахстане и сопредельных странах: Сборник научных статей / Под ред. Т. Н. Дуйсебаевой. Алматы: АСБК - СОПК, 2010. С. 172-191.
- Табачишин В. Г., Завьялов Е. В., Табачишина Е. И. Пространственное размещение разноцветной ящурки - Eremias arguta (Pallas, 1776) на севере ареала в Поволжье // Современная герпетология. 2006. Т. 5/6. С. 117-124.
- Тертышников М. Ф. Индивидуальная территория прыткой ящерицы (Lacerta agilis exigua Eichw., 1831) и разноцветной ящурки (Eremias arguta deserti Gmel., 1876J и особенности ее использования // Зоологический журнал. 1970. Т. 49. Вып. 9. С. 1377-1385.
- Цвелев Н. Н. Злаки СССР . Л.: Наука, 1976. 788 с.
- Шилов И. А. Эколого-физиологические основы популяционных отношений у животных . М.: Московский университет, 1977. 261 с.
- Alcala A. C., Brown W. C. Population ecology of the tropical scincoid lizard, Emoia atrocostata in the Philippines // Copeia. 1967. No 3. P. 596-604.
- Ballinger R. E. Comparative demography of two viviparous iguanid lizards (Sceloporus jarrovi and Sceloporus poinsetti) // Ecology. 1973. Vol. 54. No 2. P. 269-283.
- Bustard H. R. The population ecology of the Australian gekkonid lizard Heteronica binoei in an exploited forest // Journal of zoology. 1970. Vol. 162. No 1. P. 31-42.
- Galan P. Structure of a population of the lizard Podarcis bocagei in northwest Spain: variations in age distribution, size distribution and sex ratio // Animal biology. 2004. Vol. 54. No 1. P. 57-75.
- Stamps J. F. Social behavior and special pattern in lizards // Biology of the reptilian. New York: Akad. Press, 1977. Vol. 7. P. 265-334.
- Vitt L. J., Congdon J. D., Hulse A. C., Platz J. K. Territorial aggression encounters and tail breaks in the lizard Sceloporus magister // Copeia. 1974. No 4. P. 990-993.
