Изучение регуляторных механизмов биосинтеза вторичных метаболитов Aspergillus Flavus с использованием инструментов биоинформатики

Автор: Потороко И.Ю., Руськина А.А., Малинин А.В.
Рубрика: Биохимический и пищевой инжиниринг
Статья в выпуске: 4 т.13, 2025 года.
Бесплатный доступ
На сегодняшний день исследование рисков контаминации зернового сырья токсигенной микрофлорой, в частности микромицетами вида Aspergillus flavus и его вторичными метаболитами, в числе которых Афлатоксин В1, является актуальным направлением для глубинного изучения возможности минимизации рисков продуктов переработки зерна. Возможности инструментов биоинформатики позволяют на уровне прогнозирования раскрыть механизмы технологий обеззараживания и определить уровень воздействия процессов обеззараживания для блокирования рисков. Цель данного исследования заключалась в изучении генов (aflR, aflJ, aflS, LaeA, VeA, rtfA) вторичных метаболитов токсигенного микромицета вида Aspergillus flavus для оценки основных регуляторных механизмов биосинтеза афлатоксинов и влияния на экспрессию генов активных форм кислорода (АФК) с применением инструментов биоинформатики (UGENE, AlphaFold Server, ProtParam). Было установлено, что исследование влияния активных форм кислорода (АФК) на экспрессию генов применимо для понимания механизмов устойчивости зернового сырья к риску контаминации плесневыми грибами и накоплению афлатоксинов при экстремальных климатических условиях. При анализе генома Aspergillus flavus было выявлено 56 генкластеров, которые кодируют различные ферменты, необходимые для биосинтеза вторичных метаболитов, включая афлатоксины, циклопиазоновую кислоту и афлатрем. Взаимосвязь геномной и структурной информации с физикохимическими свойствами критически важны для понимания патогенеза, устойчивости Aspergillus flavus и разработки методов контроля и создания эффективных способов обеззараживания зернового сырья, что имеет прямое значение для обеспечения продовольственной безопасности и здоровья потребителя применительно к продовольственному и фуражному зерну.
Aspergillus flavus, гены вторичных метаболитов, АФК, экспрессия генов, геномная аннотация, зерно пшеницы
Короткий адрес: https://sciup.org/147252431
IDR: 147252431 | УДК: 664.696.9 | DOI: 10.14529/food250409
Study of regulatory mechanisms of biosynthesis of secondary metabolites of Aspergillus Flavus using bioinformatics tools
Currently, the study of the risks of contamination of grain raw materials with toxigenic microflora, in particular micromycetes of the Aspergillus flavus species and its secondary metabolites, including Aflatoxin B1, is a relevant area for an indepth study of the possibility of minimizing the risks of grain processing products. Over time, opportunities have emerged to use bioinformatics tools that allow us to predictively uncover the mechanisms of disinfection technologies and determine the level of impact of disinfection processes to mitigate risks. The aim of this study was to examine the genes (aflR, aflJ, aflS, LaeA, VeA, rtfA) of secondary metabolites of the toxigenic micromycete Aspergillus flavus to assess the key regulatory mechanisms of aflatoxin biosynthesis and the influence of reactive oxygen species (ROS) on gene expression using bioinformatics tools (UGENE, AlphaFold Server, ProtParam). It was found that studying the influence of reactive oxygen species (ROS) on gene expression is applicable to understanding the mechanisms of grain raw material resistance to mold contamination and aflatoxin accumulation under extreme climatic conditions. Analysis of the Aspergillus flavus genome revealed 56 gene clusters encoding various enzymes required for the biosynthesis of secondary metabolites, including aflatoxins, cyclopiazonic acid, and aflatrem. The relationship between genomic and structural information and physicochemical properties is critical for understanding the pathogenesis and resistance of Aspergillus flavus, developing control methods, and creating effective methods for disinfecting grain raw materials, which is of direct importance for ensuring food security and consumer health in relation to food and feed grain.
Текст научной статьи Изучение регуляторных механизмов биосинтеза вторичных метаболитов Aspergillus Flavus с использованием инструментов биоинформатики
I .Yu. Potoroko, , А.А. Ruskina, , Malinin, ,
На сегодняшний день Aspergillus flavus (А. flavus) и его вторичные метаболиты чрезвычайно канцерогенные афлатоксины представляют значимую угрозу для сельского хозяйства и безопасности пищевых продуктов. A. flavus имеет высокий уровень генетического разнообразия, который адаптирован к различным условиям окружающей среды, что особенно значимо для регионов, подверженных глобальным изменениям климата. Помимо афлатоксинов A. Flavus также метаболиру- ет циклопиазоновую кислоту (ЦКП), которая повреждает печень, почки и ЖКТ, но немаловажным является способность разлагать многочисленные сложные органические полимеры в биотопливо [4, 5].
Особое место в современных исследованиях занимает изучение A. flavus на генном уровне, что позволяет найти решение проблем, связанных с выработкой афлатоксинов на основе изучения молекулярных механизмов, лежащих в основе биосинтеза афлатоксина. Синтез афлатоксина (конверсия из ме- тилстеригматоцистина) требует участия белка OrdA, кодируемого геном aflQ (ordA). Гены A. flavus, участвующие в биосинтезе вторичных метаболитов, собраны в кластеры [2, 3], было идентифицировано в общей сложности 63 кластера генов вторичных метаболитов, продуцирующих афлатоксин. Среди них 20 были идентифицированы в основном геноме, 10 – в мягком ядре, 21 – в геноме оболочки, 9 – в облачном геноме и 3 – в виде синглетонов. Кластеры генов, продуцирующих афлатоксин, равномерно распределены в основном и дополнительных геномах AflaPan. Кроме того, был проведен анализ Pan-GWAS с использованием данных фенотипирования афлатоксина, продуцируемого 225 изолятами, который выявил 391 ортогруппу, связанную с продукцией токсина, включая эти 63 кластера генов вторичных метаболитов, продуцирующих афлатоксин [7].
Внешний вид колоний плесневого гриба вида A. flavus на питательной среде Чапека и его микроскопии представлен на рисунке.
Вторичные метаболиты могут вырабатываться как часть скоординированных реакций на окислительный стресс микромицета, наряду с экспрессией генов антиоксидантных ферментов. Гены биосинтеза афлатоксина и гены антиоксидантных ферментов совместно экспрессируются и высоко коррелируют с биомассой микромицетов в условиях стресса. Окислительный стресс приводит к изменениям метаболического профиля A. flavus в отношении первичного метаболизма и антиоксидантных механизмов после индукции либо АФК, либо АФК-генерирующих соединений [4].
Молекулярные механизмы вторичного метаболизма и развития плесневого гриба
-
A. flavus включают следующие процессы, требующие прогностического изучения:
– Биосинтез вторичных метаболитов. На данный биосинтез влияют различные сигналы окружающей среды и ряд белков на нескольких уровнях, включая хромосомные, транскрипционные и посттрансляционные модификации.
– Кластеры вторичных метаболитов. При анализе генома Aspergillus flavus было выявлено 56 ген-кластеров, которые кодируют различные ферменты, необходимые для биосинтеза вторичных метаболитов, включая афлатоксины, циклопиазоновую кислоту и аф-латрем.
– Механизмы ответа на окружающую среду. К ним относятся пути ответа на стресс, кворум-сенсинг и сигнальные пути G-белков.
– Функция транскрипционной регуляторной единицы группы белков Velvet Complex .
Целью данного исследования является изучение отдельных генов вторичных метаболитов плесневого гриба вида A. flavus для понимания основных регуляторных механизмов биосинтеза афлатоксинов и влияния активных форм кислорода (АФК) на экспрессию генов с применением инструментов биоинформатики.
Объекты и методы исследования
Изучение геномов A. flavus станет ресурсом для комплексной оценки процесса биосинтеза афлатоксинов, что важно для разработки методов борьбы с их производством, например, при хранении зерна. В качестве объекта исследования для изучения геномной аннотации с применением инструментов биоинформатики использовали эталонный геном плесневого гриба A. flavus , полученный из базы данных генетических последовательно-
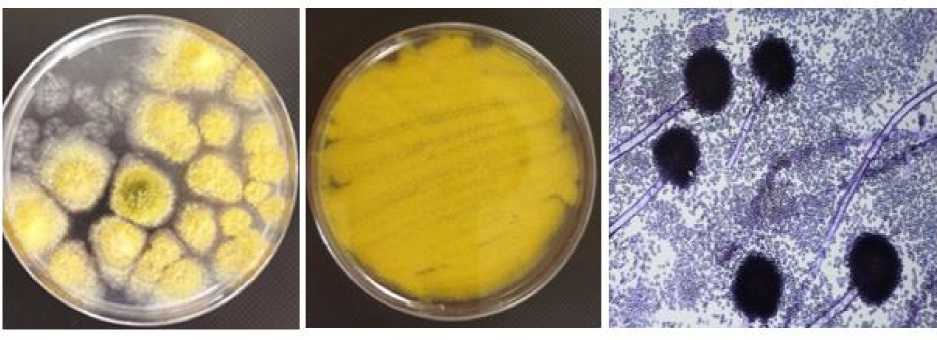
Внешний вид и микроскопия плесневого гриба вида A. flavus
стей GenBank (база данных, находящаяся в открытом доступе, содержащая все аннотированные последовательности ДНК и РНК, а также последовательности закодированных в них белков).
Для достижения поставленной цели были отобраны наиболее значимые гены вторичных метаболитов: aflR, aflJ, aflS, LaeA, VeA, rtfA. Для трансляции нуклеотидной последовательности гена в аминокислотную применяли инструменты программного обеспечения UGENE (программное обеспечение для био- информатики с открытым исходным кодом для Windows, macOS и Linux).
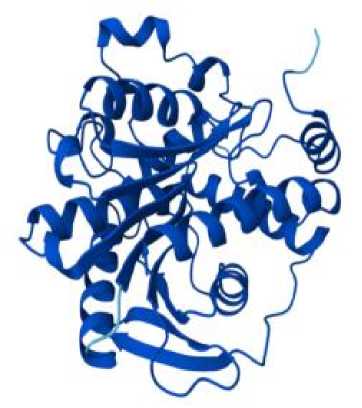
Для предсказания трёхмерной структуры генов по его аминокислотной последовательности использовалась программа на базе искусственного интеллекта AlphaFold Server с применением глубокого обучения. Модель обучается на больших наборах известных структур и последовательностей, выявляя корреляции между последовательностью аминокислот и их пространственным расположением [8, 11].
Ген aflR
MVDHISPRASPGPIRSSQTRRARKLRDSCTSCASSKVRCTKEKPACARCIERGLACQYMVSKR MGRNPRAPSPLDSTRRPSESLPSARSEQGLPAHNTYSTPHAHTQAHTHAHSHPQPHPQSHPQSN QPPHALPTPNGSSSVSAIFSHQSPPPPVETQGLGGDLAGQEQSTLSSLTVDSEFGGSLQSMEHGN HVDFLAESTGSLFDAFLEVGTPMIDPFLESAPLPPFQARYCCFSLALQTLTHLFPHAPLGCQLRL TDGEDSSCNLMTTDMVISGNKRATDAVRKILGCSCAQDGYLLSMVVLIVLKVLAWYAAAAG TQCTSTAAGGETNSGSCSNSPATVSSGCLTEERVLHLPSMMGEDCVDEEDQPRVAAQLVLSEL HRVQSLVNLLAKRLQEGGDDAAGIPAHHPASPFSLLGFSGLEANLRHRLRAVSSDIIDYLHRE
Ген aflJ
METPFAAPWHQFVEDLGQTPCLPGKDLDSILAGWGQLAGTLATRYGFPPPDESVTTEDVQLD GLWLRCYTPPNATGQEPVGLYFHGGGWVMGGVNEEDGFCRVISRQCQMRLVSVEYRKAPET RYPGALNDGVSAALWALSRYENQPLILMGTSAGGNLAFGTALRLIDQDMADKVSGVVALAPI TVHPDAVPEHLKEQYTAYEENAELTVNSRAAMQVFFDCYKAPVDDVYTSCLLHPRLLALPKV YIAELGLDTLRDDARLMKGALDTAKVPVMYDAYPGYPHCSFMFPFKSLGEHQRTFFGGVAK AVRWMS
Ген aflS
MTLTDLETCAEEIATAARTLARDGHSGGYSAGLPDHLRPVQRTLIANASQVLALASQPADLVR QLALYNQLLACLRWLGEFQVLACIPLDESVPFEDVADIAGVPECRLRRLVRPLFTIGFLCEPSPG HVAHSVMSKQFVTQPALLDAILFMSETLAPSASAMGTQTRRFGASEQAEDSAWNMAVGSDSP FAECLQQRPKVKRQLGAYLSYVSSSIDAGVEDTLTRMNWQNLGMATVVHVGAQSPSLVVAL APQFPSLRFLVQTEAKAESGGHQPCLDNHGISALKLASIPLHLRARITWGTRLSTATQPVLDAA VYLISIPFPSPQSPAMEITMRVAQALKAHVEVLRNNSDARLILTLPMSSATRSMDAAARAAVSL SDLSLLQLTNGGSLNMGEIRDLLRSRSDGLVVMREVRSPTNAVIAFEIQYRVDNDDNRY
Ген LaeA
MFGNGQTGQRLPAMASPPHDSYYSQSLASSRSRNNSDAMDIYAITDRDPPAREPSGYSQWYR NGSPSVNSIHSKSSEKQPFYEENGRMYHAYRKGVYMLPCDEQEQDRLDIFHKLFTVARVSDGL MYAPHPRNGRFLDLGCGTGIWAIDVANKYPDAFVVGVDLAPIQPSNHPKNCEFYAPFDFESPW AMGEDSWDLIHLQMGCGSVMGWPNLYRRIFAHLRPGAWFEQVEIDFEPRCDDRPLEGLAIRQ WYQYLKQATQDAMRPINHNSRDTIRDLQEAGFTDIDHQMVGLPLNPWHQDEHERKVARWY NLAVSESIESLSMAPFSRIFNWDLDRIRRISSEVKSEAFNKEIHAYNILHIYQARKPAN
Ген VeA
MATRAPLAPPPNETEASVSRITREGKKLTYKLNVMQQPERARACGAGAKSSADRRPVDPPPVV ELRVYESDPNDDLNKTDITFAYNANFFLYATLETARPMAQGRFAPNPTCPVLTGVPVAGVAYL DRPSQAGYFIFPDLSVRHEGVYRLNFHLYEETKESKDANENAPIQSMSNPMPSKPMAPKSFLEFR LEVVSVPFTVFSAKKFPGLATSTSLSRVIAEQGCRVRIRRDVRMRRRGEKRTDDYDYDEERVYR SSDRISTPDTHGYAGTPVERPRSTSTSTVDPSFPYGVDAQRRSSGATEYGFQGAQPYQRPLPPAP GPAPAAVSTPAPPAPPAPPSHNPGYQSHLSFGSTQTQYPAPQLPPTPQTASTLAAPYSPHPSYSHA RNPSTSAEYETPGYSYPPSRMSTERSSYPKNGLPPLRLEPPKPLNMPSGEPRSSDPNAYHSVAQS AAPRSQTPSSSLVPSLPPLKALSGDYPNNLSQSSSSTSQSPSHDLGAGKKFFWDTGASLSKRSYE DSFGHDDRPLYNGMRPDTESYPRRLSDASRNFYNETRDEMAYKRANGRMATKISPALQ
Ген rtfA
MALPEIQREQILSERAQEVDRHNQDLALRRLLASREREEARKAKKNKRKASAANLDEGSRKSS RQKTTLGGRKVGEASEAIEAYKRQREQKGKRDELRRRDTATKDHKSKSRVSDEDADGESEVE WDDRERSPTPPKDDPPAELRDIQRARVGRTNFAQVCFYPGFEDTMVGCYVRLNVGPNPNGVN EYRLAMITGESLTIGWRTNGSIRDQRRKEIRSGGSQRPDIHDRPVCCSRPWQNHTRVSFRCLLR LPVHRGMPLYFVHGLGHINRRNSTDGDRWQLRIAKCPRNPSLLKKLWTSTVSTISSPPKNMRN CVSKGCIPRQHFSNESTRRSSKLPKNSVMTMKLGDFKSSSQILAPAPNPVPKRSLLSMSAWPNT YGTKNLIMRTSVVRRRSERLVGKPPPPQRAEKGPTLSCASRLEQRLTTMPTSRRPNLRMEAPAP LLRVHQTQARPPGAALRPIHKISNPREVAERFVIGIWTTRTLPPWIWTSILKFRTIYSVKDAFILL SILILMAYFEYQPLRRCSIRRSGLTFDPSCTSLGMVFVWRYHQLATVCILYKNFKFGYTFENED NICRLRVGLSNHILMPKPG
Расчет физико-химических параметров белковых последовательностей генов объектов вторичного метаболизма осуществлялся при помощи веб-инструмента ProtParam .
Для представленных выше аминокислотных последовательностей вычислялись следующие параметры: количество аминокислот, теоретическая молекулярная масса, теоретическая изоэлектрическая точка, индекс нестабильности , а лифатический индекс, индекс GRAVY [6]. Далее проводилась статистическая обработка данных при помощи MS Excel.
Результаты и их обсуждение
На первом этапе исследования проводилось изучение геномной аннотации плесневого гриба A. Flavus, были отобраны наиболее важные гены вторичного метаболизма: aflR, aflJ, aflS, LaeA, VeA, rtfA. Затем с использованием аминокислотной последовательности была предсказана структура каждого гена, представленная в табл. 1. Таким образом, структура гена может быть полезна для изучения регуляторных механизмов биосинтеза афлатоксинов и влияния активных форм кислорода (АФК) на экспрессию генов через анализ регуляторных элементов и функций белков, вовлечённых в эти процессы. Структура гена определяет последовательность аминокислот в белке, а регуляторные элементы (промоторы, энхансеры, репрессорные элементы) влияют на экспрессию гена [4, 8].
Ген AflR является центральным узлом регуляторной сети, который интегрирует различные сигналы (включая АФК) для контроля биосинтеза афлатоксина. Структура белка данного гена влияет на экспрессию за счет наличия мотива кластера цинка в полипептиде aflR. При удалении гена aflR у Aspergillus flavus происходит полное прекращение синтеза афлатоксина B1, а транскрипция генов кластера (таких как aflD, aflO и др.) не обнаруживается. Это доказывает его незаменимую роль как главного переключателя пути биосинтеза.
Ген aflJ является регуляторным геном, который не кодирует белок. Он расположен рядом с геном aflR и необходим для экспрессии других генов в кластере афлатоксина. Данный ген дивергентно транскрибируется от aflR . Связывается с C-терминальным регионом гена aflR . Участвует в модуляции экспрессии генов, возможно, через взаимодействие с aflR . Делеция гена aflJ снижает уровень афлатоксинов. Данный ген кодирует эстеразу EstA , которая необходима для превращения гемиацеталя ацетата версиконала в версико-нал на одном из этапов афлатоксигенеза.
Ген aflS (ранее известен как aflJ ) является кодирующим белком. Он необходим для активации гена aflR . Переэкспрессия aflS приводит к увеличению биосинтеза афлатоксина. Соотношение регуляторных генов ( aflR/aflS ) прямо пропорционально продукции афлатоксина, например, афлатоксина B1. Температура окружающей среды влияет на уровни экспрессии регуляторных генов ( aflR и aflS ) и выработку афлатоксина.
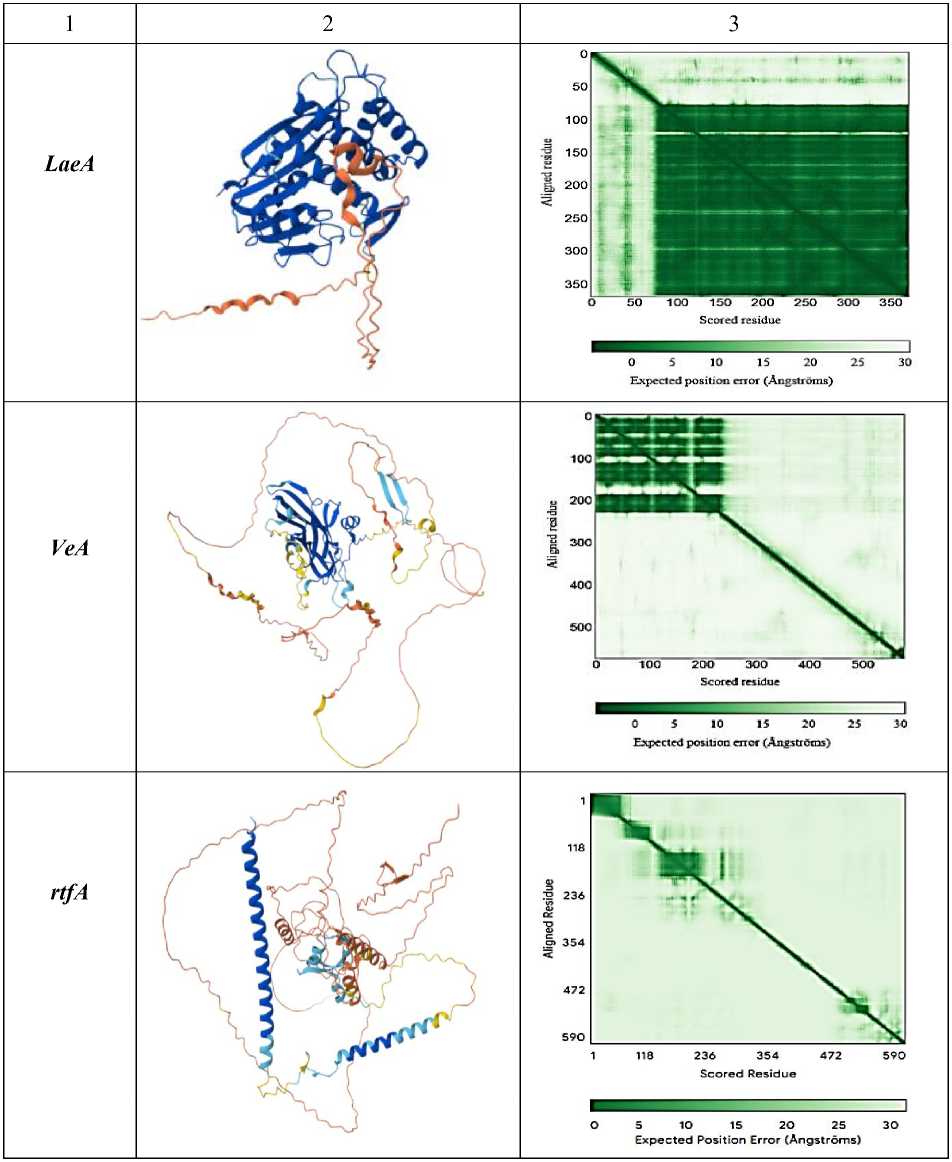
Ген LaeA необходим для транскрипционной активации гена aflR , который участвует в продукции афлатоксинов. Также LaeA регулирует кластеры генов, вовлечённые в синтез других вторичных метаболитов, например, циклопиазоновой кислоты и афлатрема. Также делеция гена LaeA у плесневого гриба может приводить к потере биосинтеза афлатоксинов.
Ген VeA может предотвращать связывание отрицательных регуляторов транскрипции с промоторами генов синтеза афлатоксинов. В некоторых случаях VeA участвует в формировании комплекса ДНК-белок, что влияет на экспрессию генов, вовлечённых в синтез афлатоксинов. Экспрессия VeA контролируется генами aflR и aflJ , которые явля-
Таблица 1
Результаты прогноза трёхмерной структуры генов биосинтеза вторичных метаболитов A. flavus с использованием AlphaFold Server
Код гена
3D структура
PAE
aflR
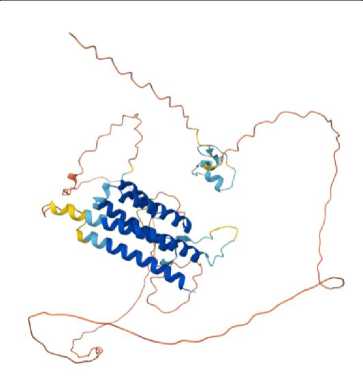
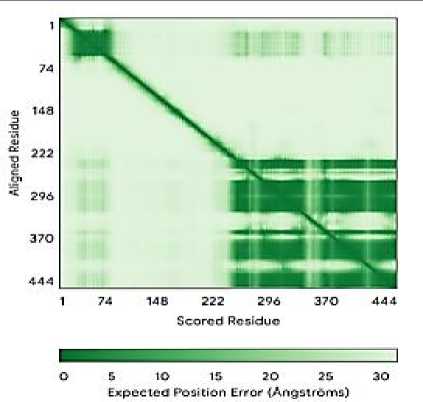
aflJ

1 39 78 117 156 195 234 273 312
Scored Residue

О 5 10 15 20 25 30
Expected Position Error (Angstroms)
aflS
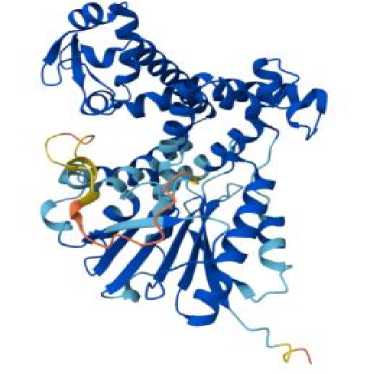
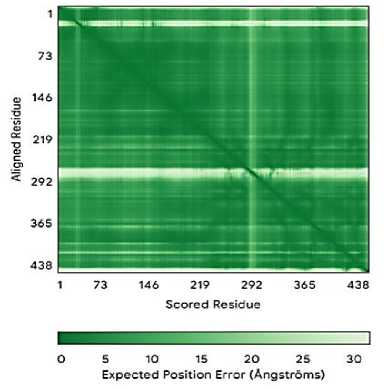
Окончание табл. 1
ются активаторами генов синтеза афлатоксинов [1, 3, 10].
Ген rtfA контролирует экспрессию генов, ответственных за синтез афлатоксинов а также влияет на экспрессию глобальных регуляторов, которые влияют на морфогенез и вто- ричный метаболизм у A. flavus, VeA и laeA. При этом удаление гена rtfA снижает выработку афлатоксина B1, но при этом изменяет выработку нескольких неизвестных вторичных метаболитов, что указывает на более широкий регуляторный эффект.
На заключительном этапе исследования проводился расчет физико-химических параметров белковых последовательностей генов вторичного метаболизма плесневого гриба Aspergillus flavus при помощи вебинструмента ProtParam. Результаты расчета представлены в табл. 2.
Полученные результаты физико-химических параметров белковой последовательности генов позволяют предсказать возможное поведение и характеристики в клетке. Для исследований белковых структур у плесневых грибов Aspergillus flavus в условиях АФК это особенно важно.
Теоретическая изоэлектрическая точка демонстрирует значение pH, при котором суммарный заряд белка равен нулю. Гены aflR, aflJ, aflS, LaeA <7 являются кислыми, в то время как гены VeA, rtfA >7 являются основными. Таким образом предсказывает, в какой среде белок будет менее растворим и может выпасть в осадок. Индекс нестабильности – это теоретическая оценка стабильности белка in vitro (в пробирке), а именно < 40: Предсказывает стабильный белок. > 40: Предсказывает нестабильный белок. Таким образом, программа предсказала, что ген aflJ является стабильным. Алифатический индекс определяет относительный объем, занятый алифатическими боковыми цепями (A, V, I, L). Высокий индекс (>70) указывает на потенциально повышенную термостабильность белка.
Высокий алифатический индекс наблюдается у генов aflR, aflJ, aflS, rtfA . Алифатический индекс может быть высоким, так как мембранные белки богаты алифатическими аминокислотами для взаимодействия с липидным бислоем. Также стоит отметить, что в ответ на АФК экспрессируется белок с высоким GRAVY и алифатическим индексом, он с высокой вероятностью может участвовать в перестройке клеточной стенки. Индекс GRAVY демонстрирует индекс, который рассчитывается, исходя из аминокислотной последовательности, и х арактеризует растворимость белка . Если GRAVY положителен, то белок является гидрофобным и, следовательно, плохо растворимым в воде (белковая структура гена aflS ) . Если GRAVY отрицателен, то белок является гидрофильным и, следовательно, хорошо растворимым в воде (белковые структуры генов aflR , aflJ, LaeA, VeA, rtfA ). Взаимосвязь параметров с функцией: белки, участвующие в ответе на АФК, часто должны быть стабильными и хорошо растворимыми в цитоплазме. Поэтому для них можно ожидать низкий индекс нестабильности (<40) и отрицательный индекс GRAVY [4, 6, 9, 12].
Выводы по результатам работы
Таким образом, применение инструментов биоинформатики позволяет осуществлять геномную аннотацию, которая важна для понимания структуры и функций генома, а также для выяснения биологической роли генов в
Таблица 2
Результаты расчета физико-химических параметров белковой последовательности генов продуктов вторичного метаболизма плесневого гриба A. flavus
Aspergillus flavus использует АФК как сигнал об изменении окружающей среды, чтобы перенастроить свой метаболизм, укрепить клеточные структуры и модулировать биосинтез чрезвычайно канцерогенных афлатоксинов, что в итоге повышает его шансы на выживание и успешную колонизацию субстрата. Понимание этого процесса с применением инструментов биоинформатики имеет практическое значение для поиска способов подавления вирулентности и токсинообразо-вания микромицета вида A. flavus.
Из 23 генов, выделенных в настоящее время, способных координировать белки, участвующие в биосинтезе афлатоксинов, только ген aflR кодирует фактор транскрипции, реагирующий на сигналы окружающей среды, и может регулировать экспрессию других генов.
