Энергетический обмен у молодняка крупного рогатого скота при нарушениях кислотно-щелочного баланса крови

Автор: Бусловская Л.К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обмен веществ и продуктивность
Статья в выпуске: 6 т.40, 2005 года.
Бесплатный доступ
Изучали особенности обмена энергии у бычков красной степной породы при воздействии факторов, вызывающих сдвиги кислотно-щелочного баланса крови. Определяли характер нарушений и компенсаторные реакции. Оценивали затраты энергии и эффективность производства продукции при адаптации крупного рогатого скота к стрессорам.
Короткий адрес: https://sciup.org/142133131
IDR: 142133131 | УДК: 636.2:591.05
Energy metabolism in young cattle during the disturbing of acid-base equilibrium of blood
The author studied the peculiarities of energy metabolism in young bulls of the Red Steppe breed under the influence of the factors caused the disturbing of acid-base equilibrium of blood. The character of the disturbing and the compensatory reactions were investigated. The energy consumption and effectiveness of production during adaptation of cattle to stress were estimated. It was shown, that energy consumption under the influence of the stress raises regardless of the disturbing character and compensation stage. The fraction of energy is required for the liveweight gain and the consumption of the metabolic energy increase and it causes the unproductive feed expenditure. These data necessitate a correction for energy rations during the cattle keeping.
Текст научной статьи Энергетический обмен у молодняка крупного рогатого скота при нарушениях кислотно-щелочного баланса крови
Какова «цена адаптации», как изменяются затраты энергии при нарушениях кислотно-щелочного баланса разного характера и при адаптации к последним, каковы при этом продуктивность животных и эффективность производства продукции — эти вопросы входили в задачу нашей работы.
Методика. Объектом исследования служили бычки красной степной породы живой массой 60,3 кг в возрасте 1 мес, подобранные по методу аналогов (по 6 гол. в каждой группе); животных содержали при температуре воздуха 18-20 оС. При создании экспериментальных моделей для оценки нарушений кислотно-щелочного баланса использовали метод функциональных дозированных нагрузок. В качестве нагрузки использовали температуру воздуха как фактор, к которому организм животных приспосабливается в естественных условиях.
В первой серии экспериментов (I опыт), проводимых в климатической камере, ацидотический сдвиг метаболической природы вызывали посредством воздействия в течение 2 ч температуры - 3... - 5 оС при относительной влажности воздуха 70 %. Во второй серии экспериментов (II опыт) (также в условиях климатической камеры) алкалозный сдвиг респираторной природы возникал под воздействием в течение 4 ч температуры 38 оС при относительной влажности воздуха 40 %.
Для оценки адаптивных реакций опыты повторяли ежесуточно и неоднократно. Во всех сериях экспериментов анализировали сопряженность нарушений кислотно-щелочного баланса с биоэнергетическими процессами. Контролем служили исходные данные тех же животных.
Кислотно-щелочной баланс крови оценивали по истинной (рН ист. ) и метаболической (рН мет. ) концентрации ионов водорода, парциальному давлению углекислого газа (рСО 2 ), концентрации буферных оснований (БО), изменению концентрации буферных оснований ( ∆ БО), стандартной (СБ) и истинной (ИБ) концентрации бикарбонатов, общей концентрации углекислого газа (тСО2). Параметры определяли микрометодом Аструпа в модификации Зиггаард-Андерсена (6).
Затраты энергии рассчитывали по показателям теплопродукции, которые определяли масочным методом по газообмену. Для выявления потребности в обменной энергии использовали формулу ОЭ = ТП + ЭП, где ОЭ — обменная энергия, ТП — теплопродукция, ЭП — затраты энергии на прирост живой массы (МДж/сут) (7).
Дополнительную потребность в энергии на прирост живой массы у растущих животных рассчитывали на основе математической модели энергетического питания крупного рогатого скота с учетом живой массы и планируемого суточного прироста последней (8). Теплоприращение принимали за часть обменной энергии, выделяющейся в виде тепла и расходуемой сверх энергии поддержания физиологических функций. Эффективность производства продукции рассматривали как отношение потребности в энергии на прирост живой массы к теплопродукции, коэффициент продукции — как отношение затрат энергии на прирост живой массы к содержанию обменной энергии (9).
Результаты. В I опыте у бычков усиливалась двигательная активность и появлялась холодовая дрожь. Потребление кислорода в 1-е сут охлаждения уже через 1 ч возросло в среднем на 30,6 % (Р < 0,001) и сохранялось высоким в течение всего периода эксперимента.
Усиление метаболизма при охлаждении способствовало повышению концентрации СО 2 . При недостаточной легочной вентиляции это создавало угрозу сдвига кислотно-щелочного равновесия. Однократное охлаждение животных при температуре - 3... - 5 оС привело к небольшому повышению рСО 2 и накоплению в крови кислых продуктов обмена, о чем можно было судить по ∆ БО (P < 0,01) (табл. 1). Уменьшилась концентрация буферных оснований (БО) и бикарбонатов (СБ), что связано с расходованием этих соединений на нейтрализацию нелетучих кислот; снизились концентрация в крови СО2 и рН; возник ацидоз, который характеризовался как метаболический. При этом значения рНист. находились в нижних пределах нормы, что свидетельствует об ацидозе компенсированной формы. Через 2 ч после воздействия пониженной температуры признаки ацидоза сохранялись. Кислотно-щелочной баланс восстанавливался только после завершения опыта.
1. Показатели кислотно-щелочного баланса крови бычков красной степной породы под воздействием температурных стрессов
|
Показатель |
Метаболический ацидоз |
Респираторный алкалоз |
||||
|
Исходный показатель |
I опыт (-3…-5 оС) |
Исходный показатель |
II опыт (38 оС) |
|||
|
1 ч |
2 ч |
1 ч |
2 ч |
|||
|
рНист., ед. |
7,383 ± 0,004 |
7,345 ± 0,013* |
7,350 ± 0,013 |
7,377 ± 0,004 |
7,394 ± 0,004* |
7,379 ± 0,004 |
|
рНмет., ед. |
7,404 ± 0,010 |
7,340 ± 0,009*** |
7,361 ± 0,014* |
7,388 ± 0,004 |
7,375 ± 0,003* |
7,373 ± 0,009 |
|
Парциальное давление |
||||||
|
СО 2 , кПа |
5,66 ± 0,17 |
5,85 ± 0,20 |
5,46 ± 0,20 |
5,53 ± 0,09 |
4,98 ± 0,14** |
5,70 ± 0,01* |
|
Концентрация, ммоль/л: |
||||||
|
буферных оснований (БО) |
48,3 ± 0,7 |
44,9 ± 0,2*** |
45,3 ± 1,0* |
47,2 ± 0,2 |
46,3 ± 0,3* |
46,1 ± 0,6 |
|
∆ БО |
0,0 ± 0,8 |
–3,4 ± 0,2** |
–2,8 ± 0,9* |
–0,9 ± 0,2 |
–1,7 ± 0,3* |
–1,9 ± 0,6 |
|
бикарбонатов: |
||||||
|
стандартная |
23,0 ± 0,7 |
20,2 ± 0,2** |
20,6 ± 0,7* |
22,4 ± 0,2 |
21,8 ± 0,3 |
21,7 ± 0,5 |
|
истинная |
24,9 ± 0,8 |
22,1 ± 0,3** |
22,2 ± 1,0 |
24,1 ± 0,3 |
22,3 ± 0,4* |
22,5 ± 0,8 |
|
тСО 2 26,2 ± 0,8 * Р < 0,05; ** Р < 0,01; *** Р < 0,001. |
23,7 ± 0,4* |
23,4 ± 1,0 |
25,5 ± 0,4 |
23,5 ± 0,5** |
23,5 ± 0,7* |
|
П р и м е ч а н и е. Описание групп и условий эксперимента см. в разделе «Методика».
При планируемом среднесуточном приросте живой массы бычков 600 г потребность в обменной энергии составляла в среднем от 17,4 до 18,8 МДж/сут, ожидаемые затраты энергии на прирост живой массы — около 5,4 МДж/сут.
В I опыте зарегистрированы максимальные затраты энергии (табл. 2). Через 1 ч теплопродукция повысилась на 31,7 %. При этом теплоприращение было весьма значительным, а ожидаемые затраты энергии на прирост живой массы снизились на 74,1 %. В результате эффективность производства продукции уменьшилась в среднем в 5 раз, коэффициент продукции — в 4 раза. Постепенно показатели приближались к исходным значениям, но не достигали их даже через 2 ч после согрева- ния.
-
2. Затраты энергии у бычков красной степной породы при нарушениях кислотнощелочного баланса под воздействием температурных стрессов
Показатель
Исходный показатель
I опыт (-3…-5 оС)
Исходный показатель
II опыт (38 оС)
1 ч 1
2 ч
1 ч 1
2 ч
Теплопродукция, МДж/сут
12,6 ± 0,4
16,6 ± 0,8**
16,2 ± 0,7**
13,7 ± 0,4
17,2 ± 0,6*
16,3 ± 0,3*
Энергия прироста живой массы (ЭП), МДж/сут
5,4 ± 0,2
1,4 ± 0,2***
1,8 ± 0,3***
5,4 ± 0,1
1,9 ± 0,3*
2,8 ± 0,1*
∆ ∆ ЭП, %
–
–74,1 ± 8,3
–66,7 ± 7,7
–
–64,8 ± 2,8
–48,1 ± 3,0
Теплоприращение: МДж/сут
2,4 ± 0,2
6,4 ± 0,4***
6,0 ± 0,3***
3,5 ± 0,3
7,2 ± 0,4*
5,1 ± 0,3*
% ОЭ
13,3 ± 0,8
35,6 ± 2,0***
33,3 ± 1,2***
18,3 ± 0,7
36,6 ± 1,2*
31,9 ± 2,5*
Эффективность производства продукции, %
42,9 ± 3,1
8,4 ± ,6***
11,1 ± 1,8***
39,4 ± 2,6
11,0 ± 0,9*
17,2 ± 1,1*
Коэффициент продукции, %
30,0 ± 2,6
7,8 ± 0,8***
10,0 ± 1,1***
28,3 ± 1,9
9,9 ± 0,9*
14,7 ± 1,3**
* Р < 0,05; ** Р < 0,01; *** Р < 0,001.
П р и м е ч а н и е. То же, что в таблице 1.
Выдерживание телят на холоде в течение 2 ч сопровождалось появлением признаков ацидоза: ∆ БО = - 2,3 ммоль/л (Р < 0,05), показатель рН ист. понижался по сравнению с исходным. При согревании бычков кислотно-щелочной баланс восстанавливался.
ммоль/л 2т
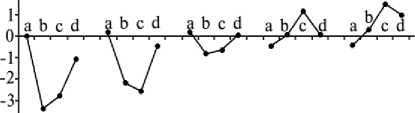
1 11 III IV V
Повторность опыта
Рис. 1. Концентрация буферных оснований в крови телят красной степной породы при адаптации к температуре воздуха –3... –5 оС: a, b, c и d — соответственно исходные данные, через 1, 2 ч после охлаждения и через 1 ч после согревания; I-V — 1-5-кратная повторность воздействия.
Ацидотический сдвиг кислотно-щелочного баланса исчезал к концу 3-кратной повторности охлаждения, и гомеостаз восстанавливался. Последующее выдерживание бычков на холоде уже не вызывало сдвигов кислотнощелочного баланса крови в сторону ацидоза; показатель рНист. крови удерживался на уровне исходных значений, что было обусловлено накоплением буферных оснований крови при повторении воздействия стрессового фактора (рис. 1). Концентрация ИБ крови после четвертой повторности воздействия холода возрастала (Р < 0,05), что приводило к увеличению концентрации СО2 (Р < 0,001).
При повторном воздействии низкой температуры наибольшие изменения происходили через 1 ч, а начиная с 3-кратной повторности — уже через 2 ч пребыва- ния животных в климатической камере.
При повторении экспериментов нарушения кислотно-щелочного баланса крови и затраты энергии на приспособление организма к низкой температуре уменьшались по сравнению с первыми сутками однократного воздействия. Так, теплопродукция через 1 ч 2-4-кратной повторности достоверно увеличилась в среднем на 1,81,5 МДж/сут. Теплоприращение, то есть часть энергии, выделяемая в виде тепла, при 2-, 3- и 4-кратной повторности составляло соответственно 64,5; 44,1 и 34,3 % (от исходных значений). При той же потребности в обменной энергии ожидаемые затраты энергии на прирост живой массы уменьшились до 2,7-4,0 МДж/сут. Эффективность производства продукции при 2-, 3- и 4-кратной повторности воздействия стресса составляла соответственно 23,8; 25,8 и 28,2 %.
При ацидотических сдвигах метаболического характера наиболее эффективной была дыхательная компенсация, но ее возможности при холодовом воздействии ограничены. Несмотря на снижение объема легочной вентиляции при 2-часовом охлаждении, выделение СО 2 увеличилось в среднем на 20,4 %. Это стало возможным благодаря повышению содержания СО 2 в выдыхаемом воздухе с 2,7 до 3,8 %.
В условиях ограничения легочной вентиляции возросла роль почечной компенсации. В процессе последующих повторностей воздействия низкой температуры моча у телят приобретала более кислую реакцию, то есть почки, выводя кислые соли, восстанавливали кислотно-щелочной баланс крови.
При 5-кратной повторности воздействия стрессового фактора отмечены наименьшие изменения показателей обмена энергии: теплопродукция возросла в среднем на 1,2 МДж (Р < 0,05), теплоприращение составляло 28,5 % от исходного значения (Р < 0,01), ожидаемые затраты энергии на прирост живой массы уменьшились на 22,2 % (Р < 0,01).
Во II опыте (t = 38 оС) уже через 40 мин в крови у телят возникал компенсиро- ванный респираторный алкалоз, характеризующийся достоверным повышением рНист. крови, уменьшением рНмет. и рСО2, отрицательными показателями ДБО, уменьшением концентрации БО, и особенно ИБ (см. табл. 1). Причиной этому послужила гипервентиляция, обусловленная учащением дыхания. У телят чернопестрой и красной степной пород частота дыхания составляла по сравнению с исходной соответственно 242 и 299 % (рис. 2). Наличие одышки при сохраняющейся гипокапнии в условиях высокой температуры воздуха подтвердило преимущественную роль для организма терморегуляторных реакций. Восстановление нарушенного гомеостаза происходило к 4-му ч воздействия стресса прежде всего за счет постепенного увеличения щелочности мочи.
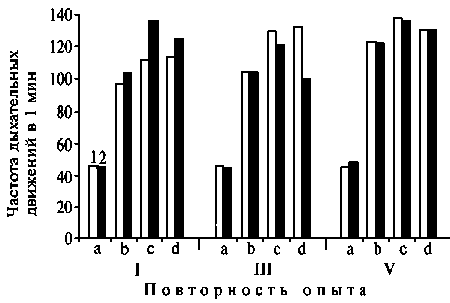
Рис. 2. Частота дыхания у телят красной степной (1) и черно-пестрой пород (2) при адаптации к температуре воздуха 38 оС : a, b, c и d — соответственно исходные данные, через 1, 2 и 4 ч после воздействия высокой температуры; I, II и III — соответственно 1-, 2- и 3-кратная повторность воздействия.
Обмен энергии у бычков при респираторном алкалозе существенно изменялся и характеризовался прежде всего достоверным увеличением теплопродукции на 10,6 %, несмотря на повышение температуры тела. Причиной этому, по-видимому, служили одышка и гипервентиляция, что способствовало увеличению потребления кислорода.
Теплоприращение возросло уже через 1 ч и достигло максимума через 2 ч (1,8 МДж). Затраты энергии на прирост живой массы при той же потребности в обменной энергии уменьшились в среднем на 33,3 %, в результате чего снизились эффективность и коэффициент производства продукции. Через 4 ч после теплового стресса все показатели обмена энергии вернулись к исходным значениям.
При ежесуточном повторении 4-часового воздействия теплового стресса степень нарушений уменьшалась и компенсация последних происходила быстрее. Несмотря на это, при 2- и 3-кратной повторности воздействия наблюдались характерные признаки респираторного алкалоза компенсированной стадии, хотя выражен- ность последнего уменьшилась.
При 3-кратной повторности наибольшие нарушения обмена энергии возникали раньше и были максимальными. Уже через 1 ч прирост теплопродукции у бычков составлял 25,5 %, теплоприращение по сравнению с исходным значением увеличивалось в 2 раза (3,5 МДж, или 36,6 % ОЭ), затраты энергии на прирост живой массы уменьшались на 65,0 %, а эффективность производства продукции снижалась с 39,4 до 11,0 %.
Во время 5-кратной повторности воздействия высокой температуры, несмотря на полипное, достоверных изменений рНист . и рСО2 не отмечено, но существенно изменилась реакция мочи — со слабокислой (у телят в возрасте 1 мес) на щелочную (увеличение составило в среднем 0,7 ед., Р < 0,001). Титрационная щелочность мочи через 40 мин и 2 ч возросла соответственно в 1,7 и 3,0 раза, что, по-видимому, позволило сохранить баланс кислых и щелочных соединений в крови. Следовательно, ме-28
ханизм выведения избытка щелочных солей почками в процессе адаптации постепенно совершенствовался.
При 5-кратной повторности по сравнению с 3-кратной затраты энергии телят уменьшались, теплопродукция максимально возросла на 18 %, теплоприращение по отношению к обменной энергии составляло 13 %, а затраты энергии на прирост живой массы и эффективность производства продукции снизились соответственно на 50 и 58 %.
Таким образом, при нарушениях кислотно-щелочного баланса у бычков красной степной породы под воздействием стрессоров значительно повышаются затраты энергии. При метаболическом ацидозе, вызванном воздействием низкой температуры среды, затраты энергии наибольшие; максимальные показатели теплопродукции и теплоприращения отмечены при однократном воздействии этого фактора. Многократное повторение стресса приводит к компенсации ацидоза, затраты энергии при этом постепенно снижаются. Респираторный алкалоз, вызванный воздействием высокой температуры среды, сопровождается меньшими затратами энергии. Повторение стресса приводит вначале к увеличению теплопродукции и теплоприращения (максимальные показатели отмечены после 3-кратного воздействия). Компенсация алкалоза сопровождается снижением затрат энергии. При увеличении затрат энергии на приспособление к действующему фактору доля последней на прирост живой массы снижается. Ожидаемые затраты энергии на прирост живой массы уменьшаются, и для достижения планируемого показателя потребность в обменной энергии значительно возрастает. Это приводит к непроизводительному расходованию кормов. Полученные данные необходимо учитывать при создании условий содержания и нормировании энергетического питания молодняка крупного рогатого скота.
Л И Т Е Р А Т У Р А
-
1. К а л ю ж н ы й И.И. Ацидоз рубца. Ветеринария, 1998, 7: 42-47.
-
2. Г р и щ е н к о В.А., Л ю б е ц ь к а Т.В., М е л ь н и ч у к Д.О. Компенсация змiн кислотно-лужного
балансу в iнтактних новонарождених телят i за умов експериментального метаболiчного ацидозу та алкалозу. Укр. биохим. журн., 1999, 71, 6: 71-75.
-
3. S a u v a n t D., M e s c h y F., M e r t e n s D. Les composantes de l,acidose ruminale et les effects acido-genes des rations. Prod. Anim. INRA, 1999, 12. 1: 49-60.
-
4. М е е р с о н Ф.З., М а л ы ш е в И.Ю. Феномен адаптационной стабилизации структур и защита сердца. М., 1993.
-
5. О з е р н ю к Н.Д. Механизмы адаптации. М., 1992.
-
6. А г а п о в Ю.А. Кислотно-щелочной баланс. М., 1968.
-
7. А г а ф о н о в В.И., Р е ш е т о в В.Б., З а б о л о т н о в Л.А. и др. Физиологическое обоснование
нормирования энергетического питания крупного рогатого скота. В сб.: Биологические основы высокой продуктивности сельскохозяйственных животных. Боровск, 1990, 1: 3-4.
-
8. З а б о л о т н о в Л.А. Физиологические аспекты в нормировании энергетического питания молодняка крупного рогатого скота. В сб.: Энергетическое питание сельскохозяйственных животных. Боровск, 1987, 34: 60-68.
-
9. К а л а ш н и к о в А.П., Щ е г л о в В.В., Г р у з д е в Н.В. и др. Новая концепция о балансе энергии в организме животного. Зоотехния, 1997, 12: 10-14.
Белгородский государственный университет,
