Эпикутикулярный воск и его роль в защите растений семейства Brassicaceae от биотических и абиотических стрессов (обзор)
")
Бесплатный доступ
На основании анализа зарубежных источников научной литературы освещена роль эпикутикулярного воска с точки зрения его морфологии и физиологии как одного из механизмов устойчивости растений семейства Brassicaceae к различным биотическим и абиотическим стрессам. Приводится детальное описание строения кутикулы и расположенных на ее поверхности эпикутикулярных восков, которые могут быть представлены в виде достаточно разнообразных композитных микроструктур. Проанализирована их важная роль в качестве эффективного и стабильного транспирационного барьера при ускорении потери воды, и установлено, что это является универсальной защитой растений при дефиците влаги. Восковые композиции различаются по своей реакции на температуру. Количество воска и морфологическая структура кристаллоидов изменяется под воздействием высоких и низких температур. Выработка воска при пониженных температурах (15 °С и ниже), как правило, увеличивается, а при температурах выше 15 °С уменьшается. Отмечено, что при повышении УФ-излучения возрастало количество воска в кутикуле растений. Воск, формирующийся на листьях растений семейства Brassicaceae, является одним из факторов, влияющих на характер повреждения насекомыми-вредителями (такими, как блошки крестоцветные, клопы и др.). Он может влиять на питание насекомых как с помощью физических механизмов, так и путем воздействия на пищеварительную систему. Процент снижения повреждения в зависимости от плотности воскового налета на поверхности растения равен 64. Эпикутикулярный воск на листовой поверхности является одним из механизмов защиты растений семейства Brassicaceae от грибных и бактериальных патогенов (Sclerotinia sclerotiorum, Alternaria brassicae и др.). Морфология, расположение и плотность кристаллоидов, а также химическая характеристика определяют, насколько эффективно растение способно бороться с различного рода инфекциями.
Эпикутикулярный воск, семейство brassicaceae, биотические и абиотические стрессоры, грибная инфекция, насекомые-вредители
Короткий адрес: https://sciup.org/142236134
IDR: 142236134 | УДК: 632.911.3 | DOI: 10.25230/2412-608X-2022-4-192-88-95
Epicuticular wax and its role in plant protection of the Brassicaceae family against biotic and abiotic stresses (review)
The article describes the role of epicu-ticular wax in terms of its morphology and physiology as one of the resistance mechanisms of plants of the Brassicaceae family to various biotic and abiotic stresses based on the analysis of foreign scientific literature. The review provides a detailed description of the cuticle structure and epicuticular waxes on its surface, which can be represented as very diverse composite microstructures. We analyzed their important role in the efficiency and stability of the transpiration barrier in accelerating water loss, and we found it to be universal protection for plants under moisture deficit. The wax compositions vary in their response to temperature. The amount of wax and the morphological structure of the crystalloids changes when exposed to high and low temperatures. Wax production tends to increase at lower temperatures (15 °C and below) and to decrease at temperatures above 15 °C. It has been observed that the amount of wax in the plant cuticle increased with increasing UV radiation. Wax formed on the leaves of plants of the Brassicaceae family is one of the factors affecting the damage pattern of insect pests (such as cruciferous fleas, bedbugs, etc.). It can affect insect feeding both through physical mechanisms and by affecting the digestive system. The percentage of damage decrease is 64 % depending on the wax thickness on the surface of the plant. Epicuticular wax on the leaf surface is one of the mechanisms of plant protection of the Brassicaceae family against fungal and bacterial pathogens (Sclerotinia sclerotiorum, Alternaria bras-sicae, etc.). The morphology, placement, and thickness of the crystalloids, as well as the chemical characteristics, determine how effectively the plant can control various kinds of infections.
Текст научной статьи Эпикутикулярный воск и его роль в защите растений семейства Brassicaceae от биотических и абиотических стрессов (обзор)
Растения подвергаются широкому спектру биотических и абиотических стрессов, включая засуху, холод, ультрафиолетовое излучение (УФ) и атаку патогенов во время их роста и развития [1]. Эти факторы первоначально воздействуют на поверхность растения, и поэтому им можно эффективно противостоять с помощью защитных механизмов, расположенных во внешней оболочке, называемой кутикулой [2].
Кутикула состоит из двух различных биохимических соединений, включающих воски и липофильную полимерную матрицу кутина [1]. Растительные воски, внедренные в кутикулу, называются «внутрикутикулярными восками», тогда как воски, находящиеся на кутикуле, называются «эпикутикулярными восками».
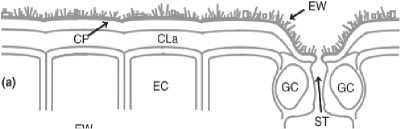
Рисунок 1 – Поперечный разрез листа растения, показывающий эпикутикулярный воск (EW); собственно кутикулу (CP), состоящую из эпикутикулярного воска, внутрикутикулярного воска и кутина; кутикулярный слой (CLa), состоящий из внутрикутикулярных восков, кутина и полисахаридов; эпидермальные клетки (EC); защитные клетки (GC) и устьица (ST) [21]
Эпикутикулярный воск чаще всего представлен в виде белой или голубоватой окраски на некоторых овощах: томат, огурец; фруктах: виноград, слива, яблоко; на листьях растений различных видов семейства Brassicaceae: капуста, рапс, горчица абиссинская. Некоторые исследователи с помощью сканирующей электронной микроскопии (СЭМ) показали, что большинство эпикутикулярных восков образуют трехмерные структуры с большими вариациями их морфологии [3].
W. Barthlott et al. и C.E. Jeffree представили широкий обзор морфологии и терминологии типов эпикутикулярного воска 13000 видов растений, основанный на анализе СЭМ с высоким разрешением. Они описали в общей сложности 23 типа воска, тонкие восковые пленки, а также более заметные восковые выступы [4; 5]. Считается, что эти выступы в основном имеют кристаллическую природу и поэтому называются восковыми кристаллоидами [6].
Восковые кристаллоиды – отчетливые восковые выступы, обычно с характерной формой, размером и ориентацией к поверхности. Они подразделяются на гранулы, тромбоциты и тарелки [4].
Примером рода с очень сложной морфологией кристаллов воска является род Brassica , в котором присутствуют несколько доминирующих восковых соединений [7] (рис. 2).
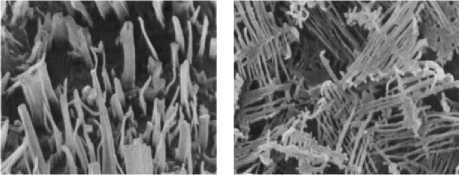
Рисунок 2 – Микроморфологическая структура восковых кристаллоидных форм Brassica oleracea L. (Brassicaceae) – полиморфизм [4]
Существует мнение о том, что микроморфология восковых кристаллоидов зависит главным образом от их химического состава. Обычно считается, что до- минирующие восковые компоненты ответственны за образование кристаллоидов. Для различных видов Brassica D.M. Hall et al. и E.A. Baker отводили главную роль кетонам на пути к образованию поперечно-ребристых лепешек [8; 9]. C.E. Jeffree также считал, что большое количество кетонов несёт ответственность за формирование разнообразных, дендридных пластинок [5]. Например, у разновидностей вида Brassica oleracea эти особые пластинки являются полиморфными кристаллоидными формами поперечно-ребристых стержней, образующихся в результате воздействия окружающей среды на ультраструктуру [4].
Растительные кутикулярные воски играют одну из значимых ролей в качестве защитного барьера от стрессов окружающей среды. Например, одной из наиболее важных функций эпикутикулярного воска является защита растений от чрезмерного ультрафиолетового излучения (УФИ). Многие исследователи показали, что повышенное УФИ влияет на образование воска в кутикуле растений. Его содержание увеличивается на 20–28 % у огурца ( Cucumis sativus L.), фасоли ( Phaseolus vulgaris L.) и ячменя ( Hordeum vulgare L.) [10].
Помимо различного рода ультрафиолетового излучения растения подвергаются широкому спектру других абиотических стрессов: дефицит или избыток влаги, повышенные или пониженные температуры воздуха и др. [10]. Некоторые исследователи утверждают, что дефицит воды и высокие температуры способствуют утолщению кутикулы у ряда видов растений, что является одной из форм адаптации к водному стрессу. Изменения в качестве, а также количестве воска также связаны с устойчивостью к засушливым условиям. Повышенная доля спиртов, кетонов и альдегидов в воске, как правило, снижает проводимость кутикулы, в то время как повышенная доля жирных кислот связана с более высокой проводимостью [11].
Также было показано, что стресс от дефицита воды может вызвать увеличение количества воска, осаждаемого на единицу площади поверхности листа у многих растений, включая обычную и твердую пшеницу, канатник, тимьян, хлопок, розу, горох, арахис и древесный табак. Увеличение количества кутикулярных восков связано с засухоустойчивостью овса, риса, сорго, люцерны и пырея хохлатого [12]. В результате изучения влияния водного стресса на растения сорго ( Sorghum bicolor L. Moench) был сделан вывод о том, что увеличение производства эпикутикулярного воска является адаптацией к засухе, которая уменьшает кутикулярную транспирацию и увеличивает отражение УФИ [13; 14]. Однако было отмечено, что количество кутикулярного воска не всегда обратно пропорционально коррелирует со скоростью транспирации или положительно – с устойчивостью к засухе [12].
В различных климатических зонах дефицит влаги часто сопровождается высокими температурами. В течение дня относительная влажность воздуха может достигать низких значений по мере повышения температуры, что приводит к соответствующему увеличению дефицита давления водного испарения. Следовательно, влияние температуры на водопроницаемость кутикулы является решающим параметром, определяющим физиологическую и экологическую роль кутикулы, особенно в условиях высокой потребности в испарении и повышенной температуры. Проницаемость кутикулы всех изученных к настоящему моменту растений, незначительно повышалась при температурах от 15 до 35 ºС, в то время как температуры выше 35 ºС увеличивали её проницаемость, что приводило к резкому увеличению водного испарения [15].
Исследования учёных на разных культурах показали, что выработка воска при более низкой температуре (15 ºС) была выше, чем при более высокой температуре (25 ºС) [16; 17]. Y. Ni et al. обнаружи- ли, что количество воска значительно увеличивается при холодовом стрессе (4 ºС). Это в основном объясняется значительным увеличением содержания алканов и вторичных спиртов [19]. Также в условиях более низких температур растения Brassica oleracea, Brassica oleracea kale и Brassica napus содержали больше продуктов декарбонилирования (алканов, вторичных спиртов и кетонов) и меньше продуктов восстановления (свободных жирных кислот, альдегидов, первичных спиртов и сложных эфиров), чем при более высокой температуре. Все эти исследования показывают, что восковые композиции различаются по своей реакции на температуру.
Температура, интенсивность света и влажность влияют на морфологию воска, и эти параметры, как правило, действуют совместно, поэтому трудно различить их соответствующие эффекты. В целом более высокие температуры благоприятствуют структурам, параллельным поверхности кутикулы, таким как пластинки и чешуйки, в то время как более низкие температуры благоприятствуют вертикальным структурам, таким как стержни и трубки. При более высоких температурах (> 35 оС) могут образовываться сложные структуры, например, дендридные решётки, наблюдаемые в восках Brassica (рис. 3).
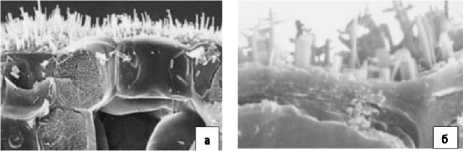
Рисунок 3 – Сканирующие электронные микрофотографии, показывающие поперечные срезы листьев Brassica oleracea (а) и Brassica napus (б) [20]
Трубчатые кристаллические формы термодинамически нестабильны из-за их высокого отношения площади поверхности к объему, и при повышении температуры, они превращаются в более плоские и термодинамически стабильные формы. Это объясняет наблюдение, где трубчатый воск на обычной брюссельской капусте превращался в дендриды в течение 48 ч после повышения температуры с 15 до 35 ºС с небольшим изменением состава [20].
Кроме неблагоприятных условий среды на растения оказывают негативное влияние и биотические стрессы. Эпику-тикулярный воск является жизненно важным компонентом комплексной устойчивости растений к насекомым-вредителям. R.H. Painter классифицировал устойчивость растений к вредителям как «непривлекательность» (переименованное Kogan и Ortman в 1978 г. в «антиксеноз»), «антибиоз» и «толерантность». Одним из примеров устойчивости растений – «антиксеноз», является снижение выживаемости вредителей на видах растений, имеющих восковой налёт [21; 22]. Среди наименее употребляемых в пищу были формы Brassica oleraceae с восковыми листьями, краснокочанная капуста, белокочанная капуста, брюссельская капуста и брокколи [23]. В 2008 г. в Словении D. Žnidarčič et al. [24] изучали химический состав эпикутикулярного воска поверхности листьев различных сортов капусты, как механизм защиты от повреждения насекомыми-вредителями, питающимися флоэмой (блошки ( Phyllot-reta spp.), крестоцветные клопы ( Euryde-ma ventral ). Насекомые-вредители в полевых испытаниях продемонстрировали различные в предпочтения в питании на разных сортах капусты. Ранние краснокочанные сорта капусты Red Dinasty и Erfurtsko rdeče имели значительно меньшее количество повреждений листовой поверхности, вызванных Phyllotreta spp. Напротив ранние белокочанные сорта Green rich, Tucana и Delphi в большей степени повреждались этим вредителем [24].
Известно, что воск на поверхности листьев может влиять на питание насеко- мых как с помощью физических механизмов, так и путем химического воздействия на пищеварительную систему. R.P. Bodnaryk [23] изучал скорость и характер питания блошки крестоцветной (Phyllotreta cruciferae) в зависимости от количества воска на поверхности листьев у некоторых видов Brassicaceae дикого типа с высоким содержанием воска и мутантов с низким уровнем вещества. Нарушения целостности листовой пластинки у изучаемых видов семейства Brassicaceae дикого типа и мутантов дают убедительные доказательства того, что эпикутикулярный воск определяет скорость и характер повреждения, нанесенного блошкой крестоцветной. Сорта масличного рапса и листовой капусты дикого типа имели высокий уровень эпику-тикулярного воска и показали рисунок повреждения только края листа. В то время как у мутантов количество эпику-тикулярного воска было снижено, и отмечалось повреждение как краев, так и всей листовой поверхности. Регрессия нормы повреждения в зависимости от количества эпикутикулярного воска составила R = 0,64. Отсюда следует, что эпикутикуляр-ный воск является одним из основных антиксенотических факторов, определяющих скорость и характер питания блошки крестоцветной [23].
Различие в количестве эпикутикуляр-ного воска на листьях и связанная с ним разница в пищевом поведении Phyllotreta cruciferae на растениях различных видов семейства Brassicaceae имели генетическую основу. Это исследование выявило, что СС-геном Brassica oleraceae является источником воскообразного характера листьев, снижающих скорость и характер питания блошки крестоцветной. Диге-номный амфидиплоид Brassica napus (геном ААСС), полученный из моногенных диплоидов Brassica oleraceae (геном СС) и Brassica rapa (геном АА), и дигеном-ный амфидиплоид Brassica carinata (геном ВВСС), полученный из моногенных диплоидов Brassica oleraceae и Brassica nigra (геном ВВ), имели воскообразные листья с плотностью воскового налёта >1000 мг/кг. С другой стороны, моноге-номные диплоиды Brassica rapa (геном АА), Brassica nigra (геном ВВ) и диге-номный амфидиплоид Brassica juncea (геном ААВВ) имели листья с низкой плотностью воскового налёта <240 мг/кг, что приводило к более высокому повреждению листовой поверхности блошкой крестоцветной. Другие факторы, не связанные с воском, такие как содержание глюкозиналатов и эфирных масел, а также продуктов их распада несомненно вносят свой вклад в повышение устойчивости различных видов семейства Brassicaceae к насекомым-вредителям [23].
Эпикутикулярные воски на листовой поверхности играют важную роль в защите растений от бактериальных и грибных патогенов. Морфология, расположение и плотность кристаллов, а также гидрофобность компонентов воска влияют на жизненно важные функции растения. Компоненты воска и химические характеристики играют важную роль в определении того, насколько эффективно растение способно бороться с инфекцией.
Y. Ni et al. изучали влияние Sclerotinia sclerotiorum на морфологию, состав и кристаллическую структуру эпикутику-лярного воска Brassica napus у устойчивого Zhongshuang 9 (ZS9) и восприимчивого Yuyou 19 (YY19) сортов. Эпикути-кулярный воск листьев Brassica napus включал алканы, спирты, длиноцепочные жирные кислоты, кетоны, альдегиды, сложные эфиры. У сортов ZS9 и YY19 содержание алканов составило 60 и 52 % соответственно от общего количества воска. У изучаемых сортов Brassica napus эпикутикулярный воск состоял из трех слоев. Между сортами ZS9 и YY19 не наблюдалось существенной разницы в морфологической структуре. Эпикутику-лярный слой состоял из пластинок, коротких стержней, ветвящихся стержней и дендридных кристаллов. Инокуляция Sclerotinia sclerotiorum привела к увели- чению количества пластинок и коротких стержней, особенно у устойчивого ZS9, и уменьшила количество ветвящихся стержней и дендридных кристаллов. Это указывало на то, что оба сорта адаптировались к грибной инфекции, изменяя структуру кристаллоидов эпикутикуляр-ного воска. Увеличение количества пластинок позволяло увеличить покрытие листа воском, уменьшая прямой контакт между спорами и поверхностью листа, что снижало процент заражения Sclerotinia sclerotiorum [18]. Эти результаты подразумевают, что эпикутикуляр-ный воск важен как для генетически устойчивых к болезням сортов, так и для восприимчивых к грибной инфекции.
Природа эпикутикулярного воска в рапсе и его роль в придании физического типа устойчивости к болезни черной пятнистости, вызываемой Alternaria brassicae (Berk.) Sacc., была изучена W.P. Skopard и J.P. Tewari [25] в 1976 г. Они показали роль эпикутикулярного воска в придании устойчивости к Alternaria brassicae на сортах сурепицы: Torch (Brassica cam-pestris L.); рапса Midas (Brassica napus L.), Tower (Brassica napus L.) и горчицы Lethbridge 22A (Brassica juncea (L.) Czern.). Для этого с листьев при помощи ватного тампона удаляли восковой налет и наносили суспензию конидий Alternaria brassicae (приблизительно 50000 кони-дий/мл), в качестве контроля оставались растения с неповрежденным восковым налетом. У листьев устойчивых сортов рапса Midas и Tower, которые характеризовались высоким содержанием эпикути-кулярного воска, после его удаления значительно повышалась восприимчивость к черной пятнистости. В то время как у восприимчивых сортов сурепицы Torch и горчицы Lethbridge содержание эпикутикулярного воска было минимальным, поэтому не выявило существенного изменения в восприимчивости к болезни при удалении воска ватным тампоном. Отсюда следует, что эпикутикулярный воск на растениях рапса и горчицы при- дает физический тип устойчивости к Al-ternaria brassicae в полевых условиях и этот признак может быть использован для выведения сортов, устойчивых к черной пятнистости [25].
Морфологические особенности, химический состав, защитные функции эпику-тикулярного воска представляют интерес в качестве одного из компонентов комплексной устойчивости к биотическим и абиотическим стрессам. Исследования ученых показали, что воски представляют собой внешнее покрытие в виде очень разнообразных композитных микроструктур. Они обладают рядом свойств и играют важную роль в эффективности расхода воды, устойчивости к высоким и низким температурам, являются фактором, определяющим повреждение насекомыми-вредителями и степень поражения болезнями.
В дальнейшем планируется изучение микроморфологии эпикутикулярных восков и плотности воскового налета на листьях сортов и гибридов рапса озимого селекции ВНИИМК.
Список литературы Эпикутикулярный воск и его роль в защите растений семейства Brassicaceae от биотических и абиотических стрессов (обзор)
- Laila R., Robin A.H.K., Yang K., Park J-I., Suh M.C., Kim J., Nou I.-S. Developmental and genotypic variation in leaf wax content and composition and in expression of wax biosynthetic genes in Brassica oleracea var. capitata // Frontiers in Plant Science. -2017. - No 7. - P. 1-14.
- Buschhaus C., Jetter R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? // Journal of Experimental Botany. - 2011. -No 62. - P. 841-853.
- Koch K. and Ensikat H.J. The hydrophobic coatings of plant surfaces: epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly // Micron. - 2008. - No 39. - P. 759772.
- Barthlott W., Neinhuis C., Cutler D. Classification and terminology of plant epicuticular waxes // Botanical Journal of the Linnean Society. - 1998. -No 126. - P. 237-260.
- Jefree C.E. The cuticle, epicuticular waxes and trichomes of plants, with reference to their structure, functions and evolution // In: Insects and the plant surface. - 1986. - P. 23-63.
- Meusel I., Neinhuis C., Markstadter C., Barth-lott W. Ultrastructure, chemical composition, and re-crystallization of epicuticular waxes: transversely ridged rodlets // Canadian Journal of Botany. -1999. - Vol. 77 (5). - P. 706-720.
- Koch K., Barthlott W. Plant epicuticular waxes: chemistry, form, self-assembly and function // Natural Product Communications. - 2006. - No 1 (11). - P. 1067-1072.
- Hall D.M., Matus A.I., Lamberton J.A.Y., Barber H.N. Infra-specific variation in wax on leaf surfaces. // Australian Journal of Biologica1 Sciences. -1965. - No 18. - P. 323-332.
- Baker E.A. Chemistry and morphology of plant epicuticular waxes. - Academic Press, 1982. - P. 139-166.
- Xue D., Zhang X., Lu X., Chen G., Chen Z.-H. Molecular and evolutionary mechanisms of cuticular wax for plant drought tolerance // Frontiers in Plant Science. - 2017. - No 8. - P. 1-12.
- Jordan W.R., Shouse P.J., Blum A. Environmental physiology of sorghum. II. Epicuticular wax load and cuticular transpiration // Crop Science. -1984. - No 2. - P. 1168-1173.
- Kim K.S. [et al.]. Changes in leaf cuticular waxes of sesame (Sesamum indicum L.) plants exposed to water deficit // Journal of Plant Physiology. -2007. - No 164. - P. 1134-1143.
- Jordan W.R., Monk R.L., Miller F.R. [et al.]. Environmental physiology of sorgum. I. Environmental and genetic control of epicuticular wax load // Crop Science. - 1983. - No 23. - P. 552-558.
- Jefferson P.G., Johnson D.A., Rumbaugh M.D. Water stress and genotypic effects on epicuticular wax production of alfalfa and crested wheatgrass in relation to yield and excised leaf water loss rate // Canadian Journal of Plant Science. - 1989. - No 69 (2). - P. 481-490.
- Bueno A., Alfarhan A., Arand K. Effects of temperature on the cuticular transpiration barrier of two desert plants with water-spender and water-saver strategies // Journal of Experimental Botany. - 2019. -No 70. - P. 1613-1625.
- Baker E.A. Influence of environment on leaf wax development in Brassica oleracea var. gemmifera // New Phytol. - 1974. - No 73. - P. 955-966.
- Wolff K. and M. Morgan-Richards. PCR markers distinguish Plantago major subspecies. // Theor. Appl. Genet. - 1998. - No 96. - P. 282-286.
- Ni Y., Guo Y.-J., Wang J. [et al.]. Responses of physiological indexes and leaf epicuticular waxes of Brassica napus to Sclerotinia sclerotiorum infection // Plant Pathology. - 2014. - No 63 (1). - P. 174-184.
- Guo Y., Guo N., He Y. Cuticular waxes in alpine meadow plants: climate effect inferred from latitude gradient in Qinghai-Tibetan Plateau // Ecology and Evolution. - 2015. - No 5 (18). - P. 3954-3968.
- Shepherd T., Griffiths W. The effects of stress on plant cuticular waxes // New Phytologist. - 2006. -No 171. - P. 469-499.
- Painter R.H. Insect resistance in crop plants. -The Macmillan company, New York, 1951. - 544 p.
- Kogan M. and Ortman E.E. Antixenosis: A new term proposed to replace Painter's non-preference modality of resistance // Eastern Southern Africa Bulletine. - 1978. - No 24. - P. 175-176.
- Bodnaryk R.P. Leaf epicuticular wax, an antixenotic factor in Brassicaceae that affects the rate: and pattern of feeding of flea beetles, Phyllotreta cru-ciferae (Goeze) // Canadian Journal of Plant Science. - 1992. - No 72. - P. 1295-1303.
- Znidarcic. D., Valic N., Trdan S. Epicuticular wax content in the leaves of cabbage (Brassica oleracea L. var. capitata) as a mechanical barrier against three insect pests // Acta Agriculturae Slo-venica. - 2008. - No 91 (2). - P. 361-370.
- Skoropad W.P., Tewari J.P. Field evaluation of the role of epicuticular waxin rapeseed and mustard in resistance to alternaria blackspot // Canadian Journal of Plant Science. - 1976. - No 57 (3). -P. 1001-1003.
