Эволюция морских палеоэкосистем и стратиграфическая корреляция

Автор: Цыганко В.С., Безносова Т.М., Лукин В.Ю., Соболев Д.Б.
Журнал: Вестник геонаук @vestnik-geo
Статья в выпуске: 11 (191), 2010 года.
Бесплатный доступ
На основе анализа эволюционных трендов и динамики разнообразия ряда широко распространенных групп животных организмов палеозоя на территории севера Евразии выявлена важная, а в ряде случаев определяющая роль событийных явлений в этих процессах. Причиной кризисов в эволюции ископаемой биоты региона, приводивших в большинстве случаев к перестройкам структуры сообществ и вымиранию, являлись прежде всего существенные изменения условий обитания или полная ликвидация биотопов в результате изменений климата или резких колебаний уровня Мирового океана.
Палеозой, сообщества, события, эволюция, эвстатика
Короткий адрес: https://sciup.org/149129003
IDR: 149129003
Evolution of marine palaeoecosystems and stratigraphical correlation
Based on the analysis of evolution trends and diversity dynamics of the number of widespread Paleozoic animals in the Northern Eurasia the important, and often crucial, role of event-driven phenomena has been determined for these processes. The reason for crises in the evolution of the regional fossil biota, causing the rebuilding of community structure and extinctions, was, first of all, considerable change of habitat conditions or complete abandoning of biotops resulted from climate changes or sharp fluctuations of global ocean level.
Текст научной статьи Эволюция морских палеоэкосистем и стратиграфическая корреляция
Наиболее важным доказательством реальности эволюции органического мира и источником представлений об однонаправленном геологическом времени является смена био-фоссилий в последовательных слоях осадочной толщи земной коры, а к основным формам эволюции биоты земли относятся процессы возникновения новых таксонов и смена их сообществ. В настоящее время существует много различных толкований понятия сообщество организмов, основывающихся прежде всего на материале, которым оперировал тот или иной исследователь. В экологии в основе этих определений лежит понятие биоценоз [1]. Он интерпретируется как группа организмов, живущих в тесной связи друг с другом и образу ющих тесное экологическое общество, или как ассоциация организмов, населяющих участок среды обитания с более или менее однородными условиями существования. В палеонтологии и палеоэкологии наиболее близким по смыслу является предложенный Л. Ш. Давиташвили [2] термин ориктоценоз, т. е. комплекс (сообщество) ископаемых организмов данного местонахождения. Выявление всех элементов ископаемого сообщества и их всесторонняя классификация представляют собой сложную задачу, требующую привлечения специалистов по многим группам (типам) палеонтологических остатков. Возможности для подобных исследований имеются далеко не всегда не только в палеоэкологии, но и в экологии живых организмов. В связи с этим чаще проводятся анализ и классификация части биоценоза или ориктоценоза, представленной одной группой организмов или выборкой нескольких организмов, обычно наиболее важных с точки зрения поставленной задачи. В этом случае используется термин свободного пользования — сообщество [3]. Реже практикуются исследования ароморфозов — степени продвинуто-сти таксонов организмов и их сообществ.
Отправной точкой в изучении ароморфозов исследуемых групп организмов стала оригинальная трактовка А. В. Каныгиным [4—6] биосферы как высшего иерархического уровня организации живых систем. В качестве модели эволюционного взры-

Основные событийные
Батиметрическая кривая
Региональные стратиграфические подразделения
Лудфордсжив
Горстииский
Падимейтывисский
Войвывекий
Маршрутнинский
Филиппъельский
Аэронский
Лолашорский
Рудданский
Яренейский
Кырьинский
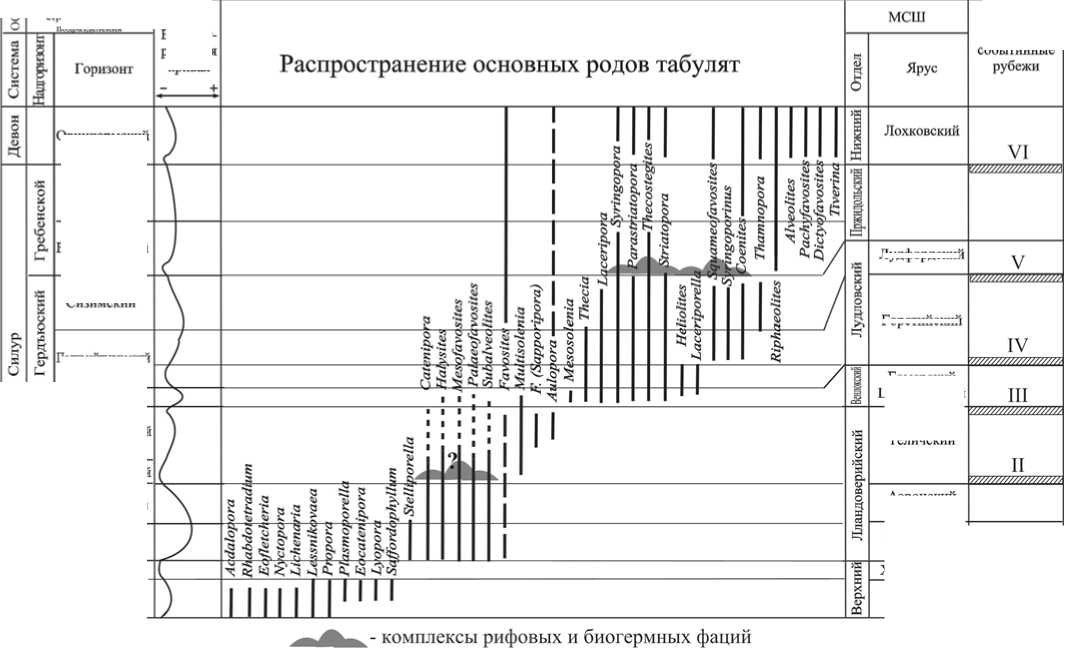
Рис. 1. Изменение родового разнообразия табулят в позднем ордовике, силуре и раннем девоне и событийные рубежи в развитии Тимано-Североуральской биоты
Хирнантский Катийский
--I—
.Bzzzzzzzzzzzzzzzzza
Овинпармскии
Карпове кий
Белушьинский
Силимский
Гомерскии Шейнвудский
Теличский
ва он детально рассмотрел ордовикский феномен взрывной радиации органического мира Земли, характеризующийся уникальным сочетанием крупномасштабных биотических и геологических событий. Появление в среднем ордовике среди морских бентосных организмов большой группы фильтраторов (строматопорат, табулят, ругоз, морских лилий и мшанок) и таких трофических универсалов, как гастроподы и остракоды, способствовало началу кардинальной перестройки морских экосистем, выразившейся в образовании новых сообществ и активном освоении ими всех экологических ниш, а также в формировании пелагиали как стабильной зоны жизни в течение всей последующей геологической истории Земли. Была установлена тесная связь между основными трендами эволюции биосферы и периодичностью, этапностью экосистемных перестроек, связанных с возникновением геоароморфозов.
Начало позднеордовикской эпохи, последовавшей за средним ордовиком — эпохой основного ордовикского эволюционного взрыва, на западном склоне Приполярного Урала характеризовалось широким распространением строматопорат, табулят, ругоз, мшанок, брахиопод и кринои-дей [7—9, 12, 18]. Доминантами воз- никшего сообщества были табуляты рода Catenipora. Регрессия морского бассейна в середине ашгильского века (малотавротинское время) и формирование сублагунных условий на западном склоне Урала привели практически к ликвидации раннеашгиль-ского бентосного сообщества организмов.
Реставрация сообщества ровного дна на новой основе была связана с трансгрессивным пульсом развития «североуральского» моря в раннекы-рьинское время [7,10,11,13]. Формирование экосистемы осуществлялось преимущественно за счет иммигрантов. Доминантами сообщества (экосистемы) стали брахиоподы Proconchi-dium muensteri (St. Joseph) и Holorhyn-chus giganteus Kiaer. Видовое название последнего отражает возникшую среди брахиопод и некоторых представителей прочих групп фауны тенденцию к гигантизму. Другие экологические ниши сообщества ровного дна данной экосистемы были освоены многочисленными табулятами, ругозами, гелиолитоидеями, губками, мшанками, гастроподами, различными водорослями.
Резкое снижение биоразнообразия и продуктивности биоты на Приполярном Урале в позднекырьинское время, вплоть до полного исчезнове ния всех бентосных форм животных, было связано с существенным снижением температуры морских вод и резким падением их уровня. Причиной массовой гибели биоты стало мощное оледенение «южных» частей материков Лаврентия и Балтия [14, 15]. В конце хирнанта — начале руддана (граница ордовик/силур) началось освоение освободившихся во время кризиса экологических ниш немногочисленными сохранившимися и зарождающимися таксонами бентосной фауны (рис. 1). Посткризисная адаптивная радиация и формирование новых сообществ организмов и экосистем являются наглядным примером экологической сукцессии бентосных организмов в начале лландоверийско-го периода раннего силура.
Менее значимое событийное явление произошло на уровне границы между силурийской и девонской системами (рис. 2). Тем не менее на Приполярном Урале этот уровень хорошо выражен как литологически, так и палеонтологически [15]. Он фиксирует начало раннедевонской трансгрессии в овинпармское время на фоне общей регрессивной стадии развития Тимано-Североуральского бассейна в позднем силуре и раннем девоне. Очень показательна динамика численности организмов сообществ ров-

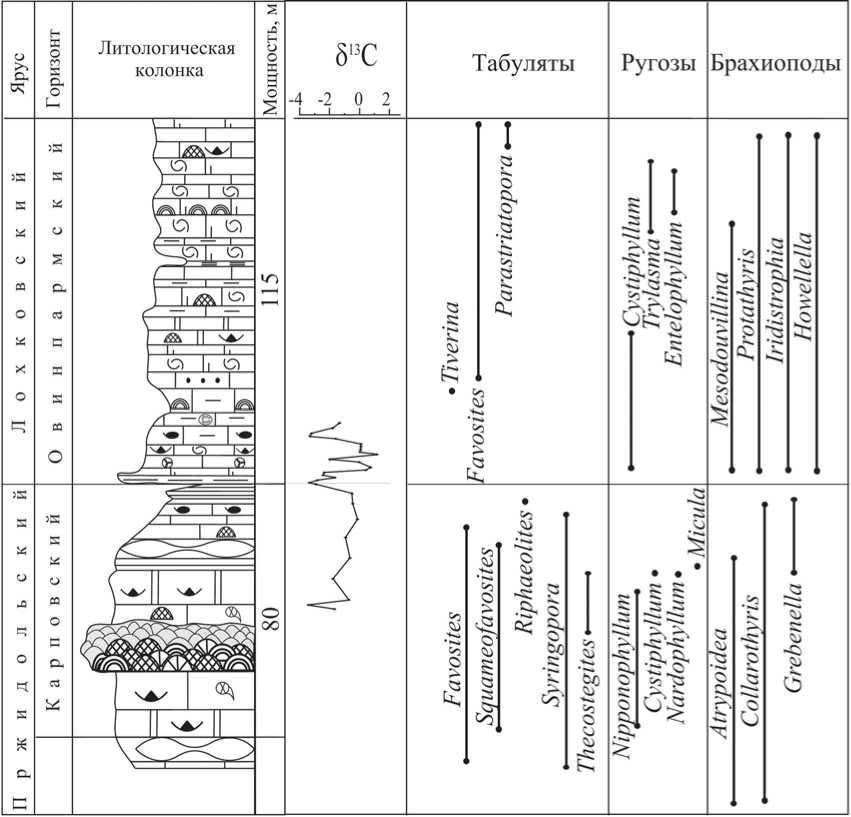
Рис. 2. Биогеологические и геохимические изменения в Тимано-Североуральском морском бассейне на рубеже силура и девона
Условные
ного дна на этом рубеже в бассейне р. Кожым. Рубеж S/D отмечен сменой регрессивного этапа развития «североуральского» морского бассейна на трансгрессивный. Преодолеть его смогли лишь два рода остракод — Hogmochilina ( H. subformosa ) и Her-rmannina . Принадлежность отложений к верхнему силуру или нижнему девону у самой границы S/D достаточно четко определяется по смене позднесилурийского сообщества брахи-опод Grebenella parvula раннедевонским — Protathyrispraecursor [7]. На этой же границе отмечается практически полная смена сообществ табулят и ругоз [10, 16, 17].
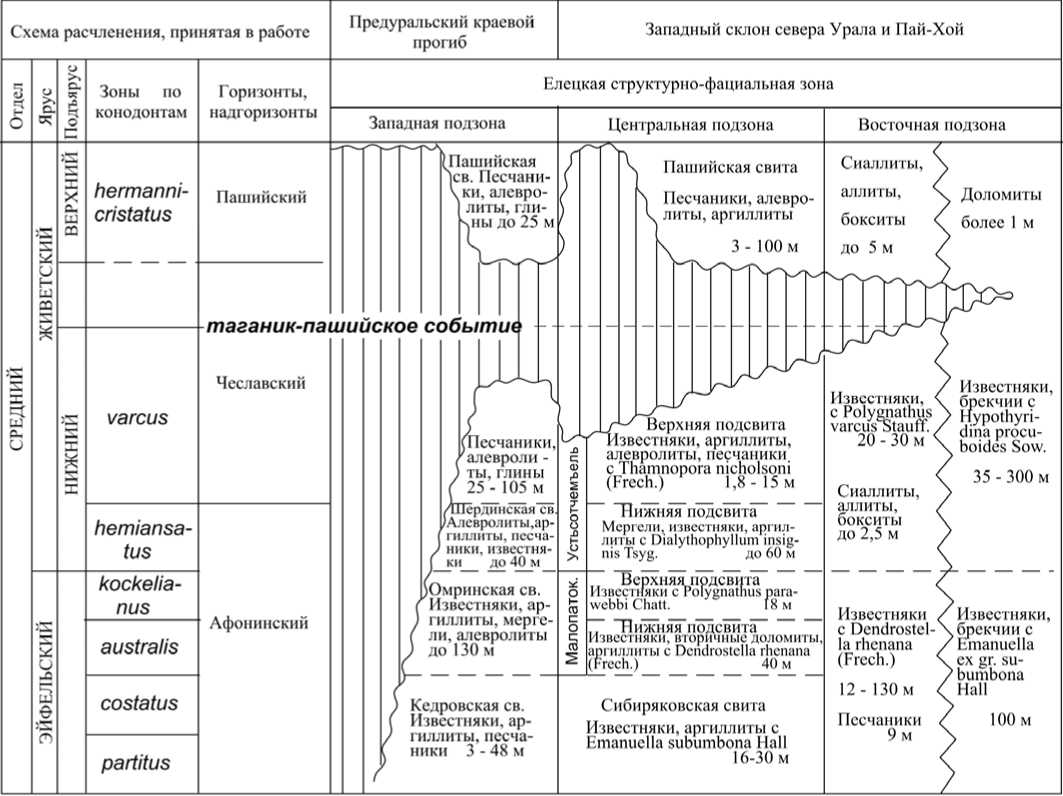
Важный рубеж в эволюции сообществ организмов приурочен к концу среднедевонской эпохи. Пусковым механизмом начавшейся в это время перестройки раннеживетских сообществ организмов в акватории прибрежных морских бассейнов континента Лавруссия, существовавшего с позднего силура, стало регрессивнотрансгрессивное таганик-пашийское событие (рис. 3) [18—20]. Оно суще ственным образом коснулось и окраин Европейского материка, входившего в состав данного континента. Событие зафиксировано практически во всех разрезах западного склона Урала и востока Русской плиты перерывом в осадконакоплении, предшествовавшим образованию пашийской свиты.
Трансгрессивная серия поздне-живетских осадков на Приполярном Урале залегает на неровной поверхности брекчированных и закарстован-ных известняков нижнеживетского подъяруса. Ее основание представлено пашийской свитой. Завершает серию толща переслаивания песчаников, глин и песчанисто-глинистых известняков кыновской свиты. Несмотря на то, что в кыновское время восстановились близкие к нормально-морским условия обитания, населявшим эти бассейны сообществам организмов была свойственна высокая степень провинциализма. Последний был нарушен только в результате эв-статического события фран (Frasnes Event) [21], приведшего к ликвидации большинства изолированных эколо гических ниш и к широкому распространению космополитных и полире-гиональных таксонов. Уровень события фран практически совпадает с границей между средним и верхним отделами девона [22].
Характерные особенности «критических» рубежей в интервале поздний фамен — ранний карбон обстоятельно изучены на примере сообществ остракод. На основе данных о соотношении появляющихся и проходящих видов остракод в раннекаменноугольном Североуральском бассейне выявлено несколько рубежей, на которых происходили резкие изменения в составе остракодовой фауны (рис. 4).
Вблизи границы фамен/турне хангенбергское регрессивно-трансгрессивное событие в Кожымской глубоководной шельфовой впадине стало причиной массового вымирания представителей глубоководного комплекса фауны остракод. Однако в более мелководных отложениях бортов впадин проявление этого события не носило катастрофического характера.
|
Предуральский краевой прогиб |
Западный склон севера Урала и Пай-Хой |
|
|
Елецкая структурно-фациальная зона |
||
|
Западная подзона |
Центральная подзона |
Восточная подзона |
Сиаллиты,
100 м до 5 м
Схема расчленения, принятая в работе
Пашийская свита
Доломиты аллиты,
Пашийский бокситы более 1 м таганик-пашийское событие
Чеславский varcus
35 - 300 м kockelia- nus
Афонинский australis
12- 130 м costatus
100 м partitus
Песчаники, алевроли -
Пашийская кв. Песчани-\ ки, алевролиты, гли-\ны до 25 м
Известняки . с Dcndrostcl-la rhenana (Freeh.)
Песчаники, алевролиты, аргиллиты
Горизонты, надгоризонты hermanni-cristatus
Песчаники 9 м
Кедровская св. Известняки, аргиллиты, песчаники 3 - 48 м
Сиаллиты, аллиты, бокситы до 2,5 м
Юмринская св. Известняки, аргиллиты, мергели, алевролиты до 130 м
Сибиряковская свита Известняки, аргиллиты с Emanuella subumbona Hall 16-30 м
Зоны по конодонтам hemiansa-tus
Известняки, брекчии с Emanuella ex gr. subumbona Hall
Мергели, известняки, аргиллиты с Dialythophyllum insig-nis Tsyg. до 60 м
Известняки, с Polygnathus varcus stauff.
20 - 30 м '
А^верхняя подсвита Известняки, аргиллиты, алевролиты, песчаники с Thamnopora nicholsoni (Freeh.) 1,8-15 м
"ПийГняя'подсвйта-
£ —
Известняки, брекчии с Hypothyri-dina procu-boides Sow.
Верхняя .прдсвита Известняки c Polygnathus para-webbi Chart. (8 м
77 7Пия7няя’подсвйта— Известняки. вторичные доломиты, аргиллиты с Dcndrostclla rhenana (Freeh.) 40 м ты, глины ( 25 - 105 м
/Шёрдйнская'св. /Алевролиты.ар-гиллиты, песчаники, известняки до 40 м
Рис. 3. Событийный уровень в среднем девоне западного склона севера Урала
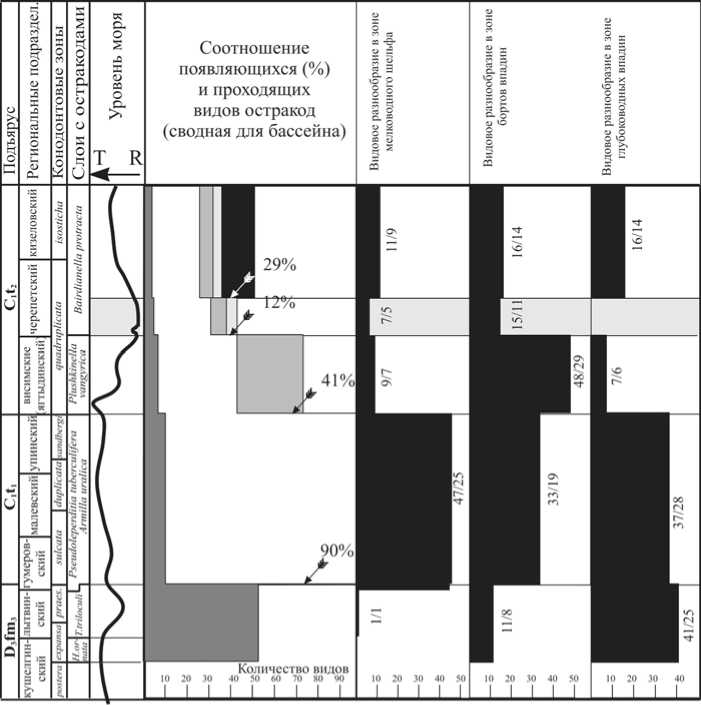
В результате изучения сообществ остракод на рубежах упинского и ви-симского (ягтыдинского), висимско-го (ягтыдинского) и черепетского, а также кизеловского и косьвинского горизонтов было установлено, что критические явления в развитии раннекаменноугольных остракод, связанные с эвстатическимии колебаниями уровня моря, по-разному отражались в различных зонах шельфа. Наиболее резко они проявлялись в мелководных и глубоководных зонах. На мелководье существенную роль играли регрессии, в результате которых происходили размывы и перерывы в осадконакоплении. В глубоководных шельфовых впадинах «критическую» роль в развитии фауны играли трансгрессии, зачастую приводившие к возникновению бескислородных условий и массовым вымираниям биоты. Более спокойно на изменение глубин реагировали комплексы остракод, селивших-
Рис. 4. Видовое разнообразие остракод Североуральского палеобассейна в позднефаменско-турнейское время в различных зонах шельфа (цифры обозначают количество видов/родов)
ся на склонах впадин, где всегда имелась возможность к миграции на небольшие расстояния без прерывания родственных связей.
Таким образом, на основе анализа эволюционных трендов и динамики разнообразия самых распространенных групп животных организмов в палеозое на территории севера Евразии была выявлена важная, а в ряде случаев и определяющая роль событийных явлений в этих процессах. Причинами кризисов в эволюции ископаемой биоты региона, приводившими в большинстве случаев к перестройкам структуры сообществ и вымиранию, являлись прежде всего существенные ухудшения условий обитания либо полная ликвидация биотопов в результате изменений климата или резких колебаний уровня Мирового океана.
Работа выполнена при поддержке Программы Президиума РАН № 15 «Происхождение биосферы и эволюция геобиологических систем» (Проекты № 09-П-5-1010, № 09-П-5-1012).
Список литературы Эволюция морских палеоэкосистем и стратиграфическая корреляция
- Mobius K. Die Austern und die Austerwirthschaft. Berlin, 1887. S. 683-751.
- Давиташвили Л. Ш. Ценозы живых организмов и органических остатков // Сообщения АН Грузинской ССР, 1945. Т. 6. № 7. С. 530-534.
- Янин Б. Т. Терминологический словарь по палеонтологии. М.: МГУ, 1990. 134 с.
- Каныгин А. В. Ордовикский этап развития биосферы: кардинальная перестройка морских экосистем // Геодинамика и эволюция Земли: Материалы к науч. конф. РФФИ. Новосибирск, 1996. С. 170-173.
- Каныгин А. В. Ордовикский феномен взрывной радиации органического мира: экологическая революция в морских экосистемах // Биоразнообразие в истории Земли: Тез. докл. 47-й сессии ВПО. СПб., 2001. С. 37-40.
