Kinship analysis of human remains from the Sargat mounds, Baraba forest-steppe, Western Siberia

Author: Pilipenko A.S., Cherdantsev S.V., Trapezov R.O., Molodin V.I., Kobeleva L.S., Pozdnyakov D.V., Polosmak N.V.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 4 т.45, 2017.
Free access
Short address: https://sciup.org/145145336
IDR: 145145336 | DOI: 10.17746/1563-0110.2017.45.4.132-142
Text of the article Kinship analysis of human remains from the Sargat mounds, Baraba forest-steppe, Western Siberia
The Sargat culture of the Early Iron Age has existed for about 1000 years (from the middle of the I millennium BC to the middle of the I millennium AD) and occupied the vast areas of the forest-steppe zone of Eurasia—from the Trans-Urals in the West to the Baraba forest-steppe (interfluve of Ob and Irtysh rivers, western Siberia) in
the East. The people of this culture lived in between the Savromatian-Sarmatian nomads and the eastern nomads of southern Siberia and Central Asia.
A typical feature of the Sargat burial tradition was the construction of several burials beneath the same kurgan mound. Such burial complexes are found throughout the area of this culture, they comprise more than one third of all complexes in the Baraba forest-steppe (Polosmak,
1987: 16). There are two types of burial arrangement in such mounds: 1) one central burial and several others around, surrounding it and lying at the periphery of the mound; 2) several burials in a row. Another feature of the Sargat burial complexes is construction of burials at different depths—both in the subsoil and in the mound. It has been hypothesized that individuals buried under the same mound were usually inhumed at different times, and the burials might have been divided by substantial time spans (Berseneva, 2011: 129–133).
Such “multiburial” mounds are most often interpreted as family (or kin) cemeteries (Koryakova, 1988: 156; Matveeva, 1993: 149; Berseneva, 2011: 129–133). This implies a biological kinship of some degree between at least some of the deceased. But typically, neither archaeological nor physical anthropological data can be used for a reliable determination of kinship. For the Sargat culture, there are only a few cases when the kinship between skeletal individuals was convincingly confirmed by osteological methods (Kurto, Razhev, 1997). The studies analyzing the Sargat burial traditions have concluded the difficulty of the assessment of kinship structure, and the lack of possibilities for resolving this question without applying some new approaches. Paleogenetics appears to be the most powerful solution for evaluation of the degree of genetic relatedness between skeletal individuals.
In this study, the remains of individuals of the Sargat culture from two “multiburial” mounds from the Baraba forest-steppe (mound 8 of Pogorelka-2 burial site, and mound 1 of Vengerovo-6 burial site) have been analyzed by the methods of molecular genetics. The results of the study are discussed in terms of the possible relatedness of the individuals and, more broadly, in the light of the burial traditions of the Sargat population.
Material and methods
Skeletal remains and their archaeological context. The remains of individuals of the Sargat culture from two burial grounds from the Baraba forest-steppe, Pogorelka-2 (mound 8) and Vengerovo-6 (mound 1), were studied. The two sites are different in terms of their arrangement of burials under the mound.
Pogorelka-2 is situated in the Chanovsky District of the Novosibirsk Region, 2.5 km south of the village of Pogorelka, along the country road leading to the village of Osintsevo. At this burial site, more than 40 mounds were detected visually. Mound 8 was excavated in 2009 by an expedition led by V.I. Molodin. Six burials were found: five adult and one subadult (burial 1). The burials are placed such that they form a chain in plan (Fig. 1). Judging from the items found in the burials, the latter can be unequivocally assigned to the Sargat culture
(Fig. 2) (Molodin et al., 2009). Burial 1 contained fragments of the skull vault of an infant. Owing to the very poor preservation of its bones, this burial could not be sampled for the genetic study. Burials 2 to 6 yielded the remains of five adult individuals, in various states of preservation. Samples from all 5 individuals (long bone fragments) were taken (Table 1, Pg1–Pg5).
Vengerovo-6 is situated in the second fluvial terrace of the right bank of Tartas River, 2.5 km south from the village of Vengerovo, in the Vengerovsky District, Novosibirsk Region. The site comprises two mounds. Mound 1 was studied in 2011 by Molodin (Molodin et al., 2011). Five burials of adult individuals were excavated (Fig. 3, 4). Burial 1 was situated in the center of the mound, while other burials were at its periphery, in the western, southwestern, and southern parts of the burial ground. This is one of the two most typical patterns of burials: one central burial is surrounded by several peripheral ones (see above). All burials had been looted. Only the skeleton of the individual from burial 3 was found in the correct anatomical order, while in all other burials bones were commingled. Burial 5 contained only poorly preserved fragments of bones, not suitable for sampling. The individuals from burials 1–4 were sampled for the genetic study (Table 1, Sg1– Sg4). The bones from burials 1–3 were well preserved, while the remains from burial 4 were poorly preserved. According to the gravegoods found in the burials, they can be assigned to the Sargat culture (Fig. 5, 6).
Preliminary treatment of the skeletal samples and DNA extraction. The protocol of this study is described in our previous publications (Pilipenko et al., 2015, 2017). In order to eliminate modern DNA contamination, the external surface of the samples was treated by 5 % sodium hypochlorite and then irradiated with UV. The external bone layer (~1–2 mm thick) was mechanically removed. Fine bone powder was then drilled out from the cortical layer. Then, the powder was incubated in a 5M guanidine thiocyanate buffer at 65 ºC, while being constantly mixed during incubation. DNA extraction was performed using a phenol/chloroform protocol with subsequent sedimentation with isopropanol.
Analysis of genetic markers. This study employed four systems of genetic markers: mtDNA (HVR I region was employed in this study as a matrilineal kinship marker); polymorphic fragment of the amelogenin gene (sex marker); nine autosomal STR-loci (universal markers of kinship); 17 STR-loci of the Y-chromosome (employed in this study as patrilineal kinship markers) (see (Pilipenko, Trapezov, Polosmak, 2015)). The genotyping protocols for all marker systems are described below.
Amplification of the mtDNA HVR I region was carried out using two different protocols: four short overlapping fragments in one round of PCR (Haak et al., 2005), and one long fragment by nested PCR (consisted
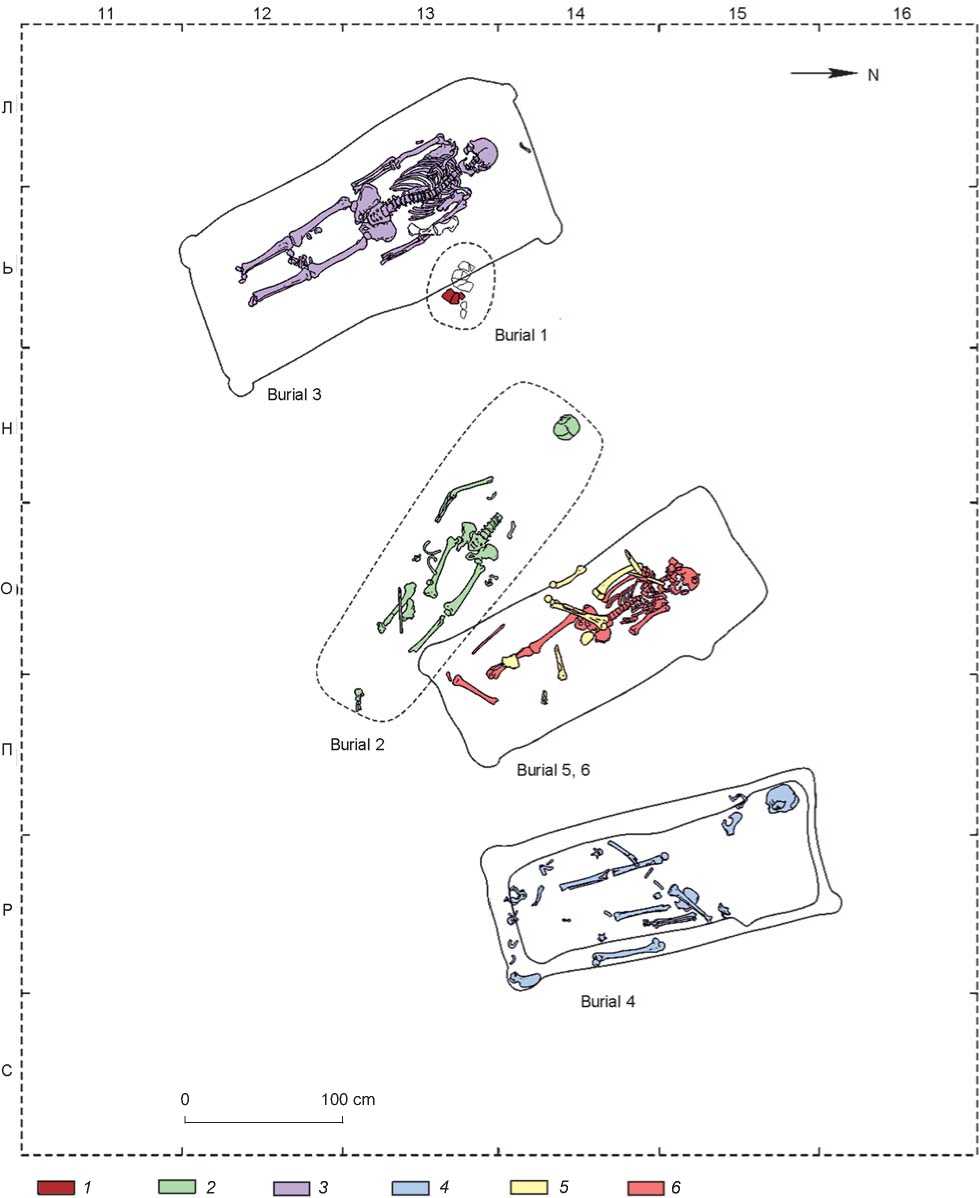
Fig. 1. Layout of burials in the central part of mound 8 of the site of Pogorelka-2.
1 – human remains from burial 1; 2 – human remains from burial 2; 3 – human remains from burial 3; 4 – human remains from burial 4;
5 – human remains from burial 5; 6 – human remains from burial 6.
0 1 cm
0 1 cm
0 1 cm
0 1 cm 5
1 cm
0 1 cm
0 1 cm
1 cm
0 1 cm
«Ss,
0 1 cm
12 0 1 cm 13
16 17 18
Fig. 2. Grave-goods from mound 8 of Pogorelka-2.
1–5 – ceramic vessels; 6 – ceramic spindle whorl; 7 – horn cheek-piece; 8 – horn buckle; 9–11 – plaques-stripes of yellow metal;
12 , 13 – iron items; 14–19 – bone arrowheads.
Table 1. Skeletal individuals studied and the results of the mtDNA structure analysis
|
Code of individual |
Burial complex |
Haplotype of the mtDNA HVR I |
Haplogroup of mtDNA |
|
Pg1 |
Pogorelka-2, mound 8, burial 2 |
16147A-16172C-16189C-16223T-16248T-16320T |
N1a1a1a |
|
Pg2 |
Ditto, burial 3 |
16192T-16256T-17270T-16399G |
U5a1 |
|
Pg3 |
Ditto, burial 5 |
16366T |
H |
|
Pg4 |
Ditto, burial 4 |
16256T-16270T-16399G |
U5a1 |
|
Pg5 |
Ditto, burial 6 |
16223T-16239T-16298C-16327T-16357C |
C4a2c1 |
|
Sg1 |
Vengerovo-6, mound 1, burial 1 |
16256T-16270T-16399G |
U5a1 |
|
Sg2 |
Ditto, burial 2 |
16288C-16362C |
H8 |
|
Sg3 |
Ditto, burial 3 |
16288C-16362C |
H8 |
|
Sg4 |
Ditto, burial 4 |
No data |
No data |
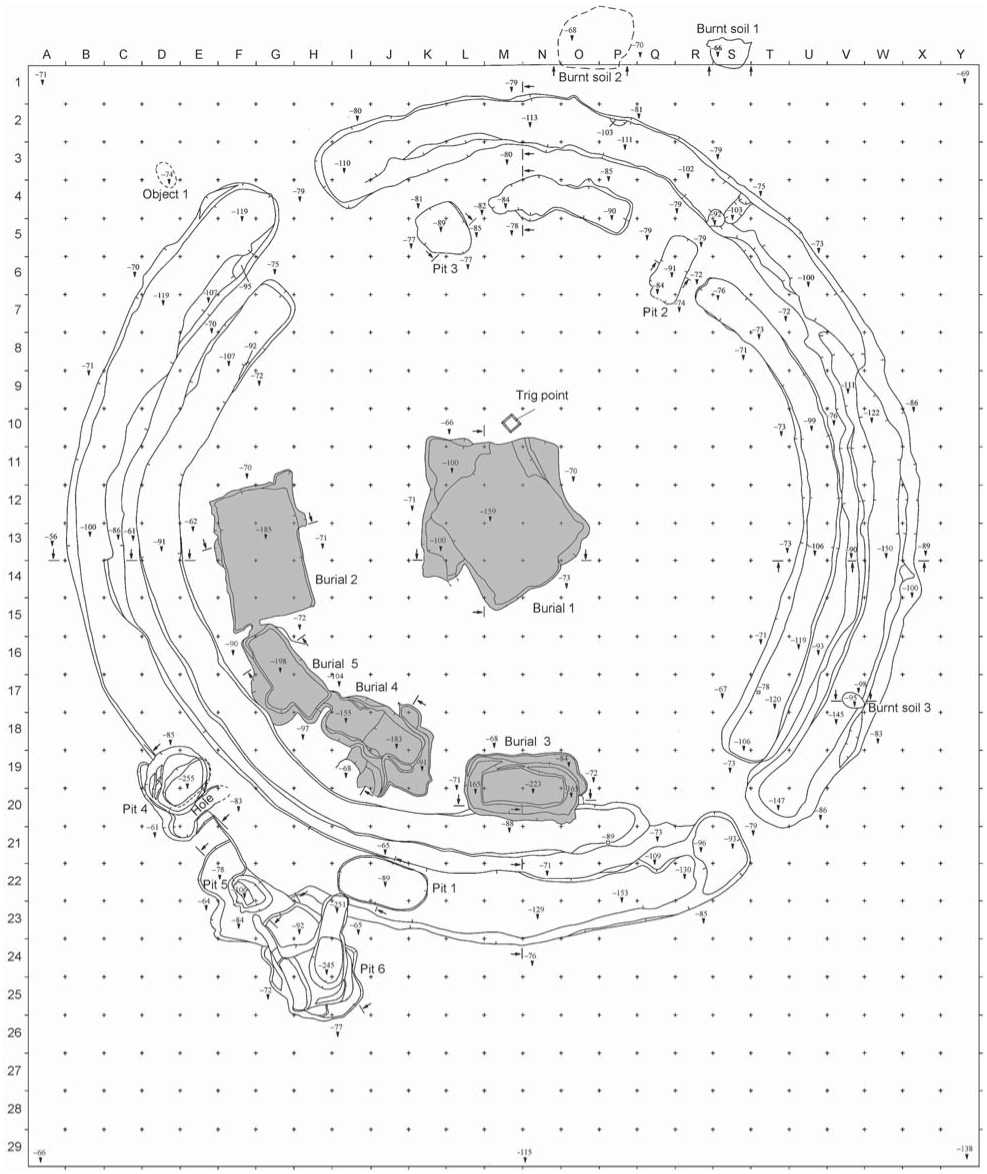
0 1 m
Fig. 3. Layout of mound 1 of the site of Vengerovo-6.
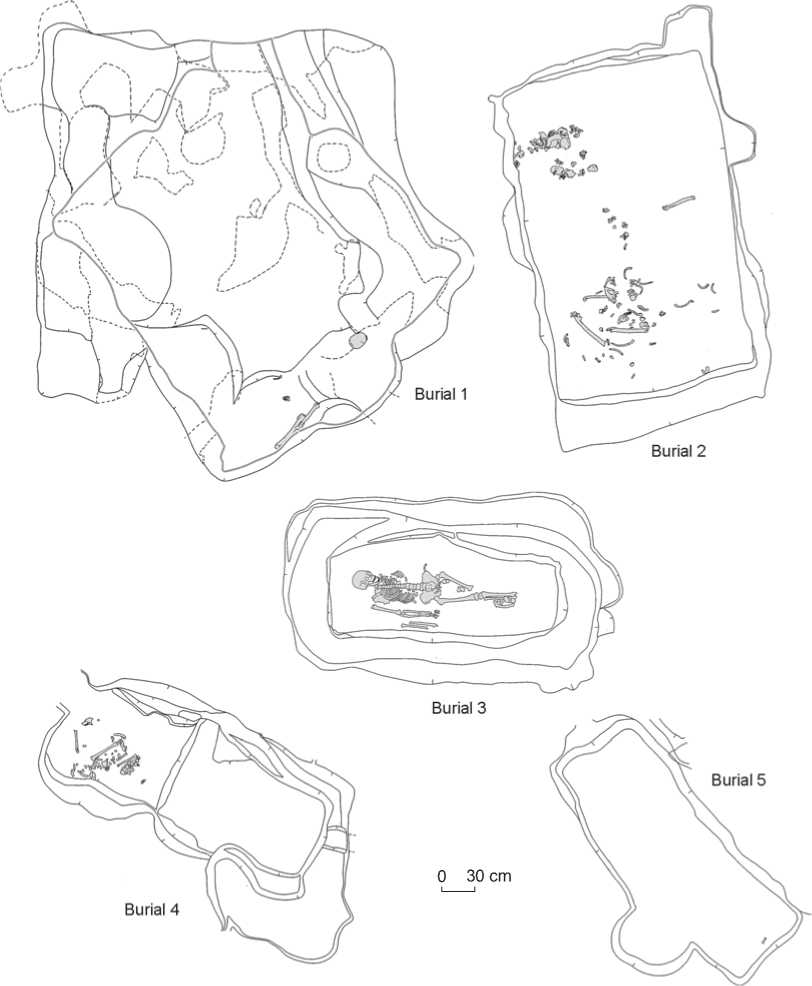
Fig. 4. Layout of burials of mound 1 of Vengerovo-6.
Fig. 5. Ceramic vessels from the burials of mound 1 of the cemetery of Vengerovo-6.

Fig. 6. Finds from the burials ( 1–8 ) and filling of the ditch ( 9 ) of mound 1 of Vengerovo-6.
of two reaction rounds) (Pilipenko et al., 2008). DNA sequencing was carried out with an ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). Sequencing extracts were analyzed with an ABI Prism 3100XL Genetic Analyzer automatic capillary sequencer (Applied Biosystems, USA) at the SB RAS Genomics Core Facility .
Profiling of nine autosomal STR-loci and analysis of the amelogenin gene region polymorphism was performed using commercial AmpFlSTR® Profiler® Plus PCR Amplification Kit (Applied Biosystems, USA), following the manufacturer’s protocol. Profiles of 17 STR-markers of the Y-chromosome were determined using commercial AmpFlSTR® Y-filer® PCR Amplification Kit (Applied Biosystems, USA), following the manufacturer’s protocol. Haplogroups of the STR haplotypes of the Y-chromosome were determined using two programs: Whit Athey’s Haplogroup predictor com/hapest5/) and Vadim Yurasin’s YPredictor 1.5.0 .
Measures against contamination and verifi cation of the results. All procedures with the skeletal specimens were carried out in a specially-equipped Laboratory for Molecular Paleogenetics (Institute of Cytology and Genetics of the SB RAS and Institute of Archaeology and Ethnography of the SB RAS, Novosibirsk, Russia). A description of the anti-contamination measures and verification of the results can be found in our previous publication (Pilipenko, Trapezov, Polosmak, 2015). Employment of those measures and consistency of the obtained results ensure reliability of our experimental data.
Results and discussion
DNA preservation. The effectiveness of molecular genetics methods in the determination of kinship of individuals in collective burials strongly depends on the degree of preservation of the DNA in the remains. For assessment of kinship between individuals, it is necessary to have data on several systems of genetic markers, including mtDNA, autosomal, and Y-chromosome STR-loci and sex-markers. Full, or at least partial, data on the markers mentioned above can be only obtained by PCR-based techniques if the preservation of the DNA in the remains is good enough to extract long (100–300 base pairs) fragments of mitochondrial, as well as nuclear, DNA.
Our previous research on heterochronous Bronze Age burial sites has shown that skeletal remains from the Baraba forest-steppe are suitable, in general, for a molecular genetic analysis, at least at the level of mtDNA. But despite climatic conditions in this region being favorable for DNA preservation, the latter varies substantially not only among different sites but also among burials at the same site, and even between individuals from the same communal burial (Molodin et al., 2012; Molodin et al., 2013). Apparently the degree of DNA preservation in skeletal remains from the Baraba forest-steppe depends on a number of factors, including the depth of a burial, local variations in soil composition and humidity, and, finally, burial traditions (the degree of the body’s decomposition before inhumation, the effect of high temperatures, etc.).
The nine samples analyzed in this study also demonstrated different degrees of DNA preservation.
Table 2. Results of the analysis of autosomal STR-loci and the sex-specific region of the amelogenin gene
|
Code of individual |
D3S1358 |
vWA |
FGA |
D8S1179 |
D21S11 |
D18S51 |
D5S818 |
D13S317 |
D7S820 |
Amelogenin (sex) |
|
Pg1 |
16/18 |
16/16 |
21/22 |
13/15 |
31/33.2 |
15/18 |
11/12 |
10/12 |
10/12 |
XX (female) |
|
Pg2 |
17/17 |
15/18 |
23/25 |
14/14 |
30/31.2 |
14/18 |
7/11 |
10/12 |
9/9 * |
XY (male) |
|
Pg3 |
16/16 |
17/18 |
22/23 |
14/16 |
30/32.2 |
No data |
11/12 |
10/13 |
12/12 * |
XY (male) |
|
Pg4 |
16/17 |
14/18 |
20/21 |
13/13 |
30/32.2 |
17/18 |
12/12 |
8/11 |
8/11 |
XY (male) |
|
Pg5 |
15/15 |
14/19 |
22/24 |
13/13 |
28/31.2 |
17/17 * |
9/11 |
10/10 * |
13/13 * |
XY (male) |
|
Sg1 |
14/17 |
18/18 |
23/24 |
14/16 |
30/31.2 |
14/14 * |
10/11 |
8/8 * |
10/11 |
XY (male) |
|
Sg2 |
17/18 |
16/16 |
20/23 |
12/13 |
31/33.2 |
12/17 |
11/11 |
11/13 |
8/9 |
XY (male) |
|
Sg3 |
14/19 |
16/18 |
21/23 |
10/13 |
30/31 |
13/13 * |
11/12 |
8/11 |
No data |
XY (male) |
* There is a possibility of the absence of a signal from the second allele, which was not amplified owing to poor DNA preservation.
In burial 4 of mound 1 in Vengerovo-6 (Table 1, Sg4) it was not possible to obtain reliable results for any of the genetic markers because of poor DNA preservation. This is concordant with the visually observed poor preservation of bone-tissue in this individual. But other skeletons yielded either full or partial molecular genetic data. In all of these individuals, the mtDNA HVR I region was determined, which made possible the evaluation of their phylogenetic status (Table 1). We also obtained full or almost full profiles of autosomal STR-loci (eight out of nine), and assessed the status of the sex-specific region of amelogenin gene (Table 2). A full profile of the 17 STR-loci of the Y-chromosome was determined for 3 out of 7 males employed in this study, an almost full profile (16 out of 17 loci) was determined for one of the individuals, while for the rest of the sample from 11 to 13 loci were determined (Table 3). In all cases, the Y-chromosome variants were unequivocally assigned to haplogroups, using the predictor software. Thus, we were able to obtain the bulk of the possible molecular data for eight out of the nine individuals who were initially sampled: five individuals from Pogorelka-2, mound 8, and three from Vengerovo-6, mound 1.
Notably, the samples from the latter (excluding the poorly preserved Sg4 specimen) exhibit a better DNA preservation than the specimens Pogorelka-2, mound 8. This is particularly evident from the results for the Y-chromosome STR-loci (Table 3). Importantly, there is a correlation between the success of analysis of a locus and the length of DNA fragment that is necessary for performing the analysis. As it could be expected for degraded ancient DNA, amplification of short segments of nuclear DNA has always been more effective than amplification of longer segments. This is an additional argument in favor of the authenticity of the studied DNA. Our results unambiguously support the high value of skeletal samples from the Baraba forest-steppe for a wide range of molecular genetics research on both mtDNA and nuclear DNA.
Sex determination. In seven out of the eight successfully sampled individuals, two variants of the amelogenin gene (typical of the X- and Y-chromosomes) were determined; thus the individuals were males. In one of the individuals, just one of the variants, typical of the X-chromosome, was found (female; see Tables 1, 2, Pg1). These molecular-genetics results are fully consistent with the preliminary sex determinations made by physical anthropologists. Interestingly, the prevalence of males is typical of the Sargat burial mounds, including multiple burials (Razhev, 2009: 74–75; Berseneva, 2011: 83).
Autosomal STR-makers and an assessment of possible direct “parent-child” kinship. It was hypothesized that individuals buried under the same mound might have been inhumed at different times, and the burials might have been divided by different time spans. One of the hypotheses suggests that the descendants could have been buried in the same mound as their ancestor (or ancestors). In the light of such views, it is important to address specifically the issue of possible direct kinship between the individuals from both mounds: five skeletons from Pogorelka-2, mound 8, and three from Vengerovo-6, mound 1. For all eight individuals, full (nine loci) or almost full (eight loci) allelic profiles of the autosomal STR-loci were obtained (see Table 2)*.
The direct kinship of “parent-child” type is imprinted in the structure of the allelic profile: direct relatives possess one common allele for each of the studied loci. Following this criterion, there are no “parent-child” pairs among the individuals employed in this study. Thus, the
Table 3. Results of genotyping of the Y-chromosome STR-loci
|
dnoj6o|dei| p910lp9Jd |
05 05 5 5 z z z z z |
|
svvsaq |
05 05 ■§ 6 □> ■§ □> □> □> ah'- о z z |
|
sevsaq |
05 05 го о го о -Q CD -q CD CD о О o Q z: z |
|
zevsaq |
05 -o'^'^'^'^'^'^ o z |
|
VH V1V9 A |
05 co CXJ CXJ -Q CXJ CXJ CXJ z |
|
36ESAQ |
05 05 ГО o ^ ro ^ ^ ^ О Q О '~ z z |
|
S£9SAa |
^ co co co co co co (M (M (M (M (M (M (M |
|
6EVSAQ |
05 T- § T- T- T- T- T- о z |
|
^6£SAa |
^ CD ^ ^ ^ ^ ^ |
|
E6ESAQ |
co co ^ ^ ^ ^ ^ |
|
q/e S8ESAQ |
05 05 5 ra 2 ro 2 2 2 О z z |
|
6tSAa |
CD CD ^ ^ ^ ^ ^ |
|
esvsAa |
05 05 § - § 2 - - о о z z |
|
II 68£SAa |
T- CN T- T- о c> c> CO CO CO CO CO CN CN |
|
06£SAa |
СОЮчГчГчГчГчГ (M (M (M (M (M (M (M |
|
168£SAa |
4j- 4j- LO LO 4j- CO CO |
|
gsvsAa |
Ю CD ^ ^ ^ ^ ^ |
|
o| 8 1 |
0к| CO ^- 1Л T- OkI CO CD CD CD CL CL CL CL (Л 0) 0) |
results of this study cannot provide empirical support for the hypothesis as to the existence of a tradition in the Sargat culture of burying descendants in the mounds of their direct ancestors.
Structure of mtDNA and the allelic profi les of STR of the Y-chromosome, matrilineal and patrilineal kinship. The uniparental markers—mtDNA (matrilineal) and the Y-chromosome (patrilineal)—are phylogenetically and phylogeographically informative for reconstruction of the genetic history of human populations. In this study we do not address these aspects, as the structure of both male and female gene-pools of the Sargat population of Baraba will be assessed by molecular data for a more representative sample in a special publication (now in preparation). Here, we only present the results of an analysis of diversity of the Y-chromosome and mtDNA lineages in the sample from the point of view of kinship between the individuals.
The sample used in this study exhibits a great diversity of mtDNA lineages. The same, according to author’s data, applies to the Sargat population in general. All five individuals from Pogorelka-2, mound 8 display different structures of mtDNA and, consequently, belong to different maternal lineages. For instance, we were not able to detect a genetic relationship of the single woman in this mound with any of the four male individuals buried there. Among three individuals from mound 1 of Vengerovo-6, two variants of mtDNA were detected. The two adult males from peripheral burials 2 and 3 possess the same variant (at least at the level of the mtDNA HVR I) that belongs to Western Eurasian haplogroup H8 (see Table 1, Sg2 and Sg3). Thus, they might be matrilineal relatives.
The analysis of allelic profiles of the Y-chromosome STR-loci (full, including 17 loci, or partial—11–16 loci) using the predictor software has assigned the seven Y-chromosome specimens (from all male individuals) to two haplogroups—N and R1a (see Table 3). In two out of four individuals from mound 8 of Pogorelka-2, variants of the haplogroups were detected as well. Allelic profiles of the individuals possessing variants of haplogroup R1a were substantially different: the difference was detected in six out of nine Y-chromosome STR-loci successfully genotyped in both individuals (see Table 3, Pg2 and Pg3). Thus, these people could not have been patrilineal relatives. In contrast, the two carriers of haplogroup N display identical allelic profiles of all 10 loci (see Table 3, Pg4 and Pg5). Therefore, a patrilineal kinship between these two individuals is plausible.
The Y-chromosomes of all three skeletons from mound 1 of Vengerovo-6 belong to haplogroup N. Also, the two individuals for whom full allelic profiles of the 17 STR-loci were determined possessed identical variants of the Y-chromosome (see Table 3, Sg2 and Sg3). Taking into account the fact that their mtDNA variants were also identical, we might reasonably suggest that these two individuals were close relatives on both maternal and paternal sides. Such a pattern of genetic similarity can be observed in siblings. Another possibility might be that they had one common parent of either sex, while the second parents were siblings. More distant kinship cannot be excluded, but is less probable.
The structure of the Y-chromosome variant of the third individual from the same mound (16 STR-loci) is quite close to the two others, but differs by 3 loci out of 16. Thus, this individual from the central burial cannot be a close patrilineal relative of the individuals from the peripheral burials. The similarity of the structure of the allelic haplotype could point to a more distant relatedness, i.e. at the level of kin or several paternally related kin. The similarity of allelic variants of haplogroup N, observed in some specimens from both sites, is a solid argument for the presence in the Sargat population of Baraba individuals who were not direct patrilineal relatives but shared a common patrilineal origin (e.g. kin or clans) in the relatively recent past. So, the individuals for whom full data on the Y-chromosome STR-loci allelic profile were obtained (one from mound 8 of Pogorelka-2 and two from mound 1 of Vengerovo-6; see Table 3, Pg4, Sg2, and Sg3), shared a common allelic motif, which unites 15 out of 17 STR-loci. Such similarity suggests a remote paternal kinship between these individuals.
Summing up, our molecular genetic data enable us to make some preliminary inferences regarding the kin relationships of individuals buried in the same mound. To the moment, we have not found any empirical evidence for burying adult close relatives (“parentchild”) under the same mound. Some cases of close patri- and matrilineal kinship were detected, which suggests that such kinship might be a reason for burying people in the same mound. At the moment, patrilineal kinship appears to be a more important factor: the only case of possible matrilineal kinship was observed in the pair of individuals who possessed identical variants of the Y-chromosome as well (see Tables 1–3, Sg2 and Sg3). Meanwhile, not all people buried in the same mound were close relatives. For instance, the presence of individuals not connected even by a remote kinship in the same mound was detected: the best example is the carriers of haplogroups R1a and N from mound 8 of Pogorelka-2. The diversity of mtDNA variants is even higher. Thus, other reasons for burying in the same mound, besides kinship, existed in the Sargat society. One of the reasons might have been membership of a particular social group, i.e. a military elite (Razhev, 2009: 74–75). Then, the burial of individuals not connected by close blood kinship in the same mound can be inferred. Such inference corresponds well with the high prevalence of males, as compared to females and children, in burial mounds. Meanwhile, only one female skeleton from a mound with several burials was studied, which prevents conclusions regarding kin relationships between females and males from such mounds. It can only be pointed out that the absence of direct blood kinship of “mother-son” type between the female and the males buried in this mound might point towards another type of relationship of the female with one of the males, e.g. marital.
The presence of individuals possessing not identical but very similar variants of the Y-chromosome in mounds from different sites separated by dozens of kilometers could provide evidence for a particular role of some groups of males of shared patrilineal ancestry in the Sargat society, maybe “male” kin or clans. We realize that this inference, like other conclusions of this study, is preliminary and requires additional confirmation. There is also a demand for a much more detailed molecular genetic analysis of numerous additional samples from skeletons of the Sargat culture from different regions of the Baraba forest-steppe, and also from other areas occupied by this culture in the Trans-Urals forest-steppe and western Siberia.
Acknowledgements
This study was supported by the Russian Science Foundation (Project No. 14-18-03124). The use of paleogenetic facilities of the Institute of Cytology and Genetics of the SB RAS was covered from the funds of the public contract for Project No. 0324-2016-0002.
