Комплексная оценка молочной продуктивности коров в разрезе генотипов

Автор: Ильина А.В., Абрамова М.В., Зырянова С.В., Евдокимов Е.Г., Муштукова Ю.В., Приданникова Н.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Зоотехния и ветеринария
Статья в выпуске: 10, 2025 года.
Бесплатный доступ
Цель исследования – изучение влияния полиморфизма генов на молочную продуктивность и производственную типичность крупного рогатого скота ярославской породы. Задачи: изучить распределение генотипов и генетическое разнообразие по генам молочной продуктивности в популяции крупного рогатого скота ярославской породы; оценить молочную продуктивность коров в зависимости от генотипа по генам: синтаза жирных кислот (FASN), тиреоглобулин 5 (TG5), гипофизарный фактор транскрипции 1 (Pit 1), диацилглицерол Оацилтрансфераза 1 (DGAT 1). Научнопроизводственный опыт проводился в племенных стадах Ярославской области в период 2024–2025 гг. Объект исследования – чистопородные коровыпервотелки ярославской породы (229 голов). Анализировали показатели молочной продуктивности (удой за 305 дней, содержание жира и белка, количество молочного жира и белка), промеры экстерьера (2–4й месяц первой лактации), биологический материал (цельная кровь). Ген DGAT 1 мономорфен, это связано с направленным искусственным отбором. Ген FASN показал умеренное генетическое разнообразие, эффективное число аллелей 1,68, информационный индекс Шеннона 0,59, наблюдаемая гетерозиготность 0,48, ожидаемая гетерозиготность 0,40, индекс фиксации 0,18. Ген PIT1 имеет два аллеля, но более низкое эффективное число аллелей – 1,61, информационный индекс Шеннона 0,57. При этом отмечали существенное отклонение наблюдаемой гетерозиготности (0,28) от ожидаемой (0,38), что подтверждалось положительным значением индекса фиксации (0,26). По гену TG5 выявлено наибольшее генетическое разнообразие: эффективное число аллелей 1,91, информационный индекс Шеннона 0,67, наблюдаемая гетерозиготность 0,62, ожидаемая гетерозиготность 0,48, индекс фиксации –0,29. Исследование молочной продуктивности показало, что генотип гена FASNAG оказывает наибольшее влияние на удой. Коровы с этим вариантом давали на 639 кг молока больше, чем коровы с вариантом FASNGG (p ≤ 0,001). Коровы с генотипомTG5CT на 377 кг превосходили по удою коров с вариантом TG5CC. Коровы с вариантом FASNAG имели более высокое содержание жира в молоке (4,56 %, р ≤ 0,05) по сравнению с коровами с FASNAA (+0,29 %). Коровы с генотипомTG5CT имели более высокое содержание белка (3,32 %, р ≤ 0,001) по сравнению с коровами с вариантом TG5СC (+0,12 %). Все исследованные животные относились к молочному типу, наибольший показатель коэффициента производственной типичности выявлен у коров с генотипами FASNAG (4,3, p ≤ 0,01), TG5TT (4,6) и Pit1AA (4,5). Коров с комбинацией генетических вариантов FASNAG, TG5TT и Pit1AA на основании коэффициента производственной типичности можно отнести к высокомолочному типу. Полученные данные необходимы для оценки генетической структуры породы и могут быть использованы при планировании селекционной работы.
Крупный рогатый скот, гены молочной продуктивности, генетическое разнообразие, молочная продуктивность
Короткий адрес: https://sciup.org/140312642
IDR: 140312642 | УДК: 636.2.034:636.082 | DOI: 10.36718/1819-4036-2025-10-109-121
Comprehensive assessment of cows dairy productivity in the genotypes context
The aim of the study is to investigate the influence of gene polymorphism on milk productivity and pro-duction typicality of Yaroslavl cattle. Objectives: to study the distribution of genotypes and genetic diversity for milk productivity genes in the Yaroslavl cattle population; to evaluate milk productivity of cows depen-ding on the genotype for the following genes: fatty acid synthase (FASN), thyroglobulin 5 (TG5), pituitary transcription factor 1 (Pit 1), diacylglycerol O-acyltransferase 1 (DGAT 1). Research and production expe-riment was conducted in breeding herds of the Yaroslavl Region in the period 2024–2025. The object of the study was purebred first-calf heifers of the Yaroslavl breed (229 heads). The following parameters were analyzed: milk productivity (305-day milk yield, fat and protein content, milk fat and protein content), conformation measurements (2nd–4th month of the first lactation), and biological material (whole blood). The DGAT 1 gene is monomorphic, which is due to directional artificial selection. The FASN gene showed moderate genetic diversity, an effective allele count of 1.68, a Shannon information index of 0.59, an ob-served heterozygosity of 0.48, an expected heterozygosity of 0.40, and a fixation index of -0.18. The PIT1 gene has two alleles, but a lower effective allele count of 1.61 and a Shannon information index of 0.57. A significant deviation of the observed heterozygosity (0.28) from the expected one (0.38) was noted, which was confirmed by a positive fixation index value (0.26). The TG5 gene showed the greatest genetic diversity: effective number of alleles 1.91, Shannon information index 0.67, observed heterozygosity 0.62, expected heterozygosity 0.48, fixation index –0.29. A study of dairy productivity showed that the FASNAG gene genotype has the greatest impact on milk yield. Cows with this variant produced 639 kg more milk than cows with the FASNGG variant (p ≤ 0.001). Cows with the TG5CT genotype yielded 377 kg more milk than cows with the TG5CC variant. Cows with the FASNAG variant had a higher fat content in milk (4.56 %, p ≤ 0.05) compared to cows with FASNAA (+0.29 %). Cows with the TG5CT genotype had a higher protein content (3.32 %, p ≤ 0.001) compared to cows with the TG5CC variant (+0.12 %). All animals studied were dairy cows, with the highest production typicality coefficients found in cows with the FASNAG (4.3, p ≤ 0.01), TG5TT (4.6), and Pit1AA (4.5) genotypes. Cows with a combination of the FASNAG, TG5TT, and Pit1AA genetic variants can be classified as high-yielding dairy cows based on their production typicality coefficients. The data obtained are necessary for assessing the genetic structure of the breed and can be used in planning breeding efforts.
Текст научной статьи Комплексная оценка молочной продуктивности коров в разрезе генотипов
Введение. Глобальная тенденция к индустриализации сельского хозяйства сопряжена с рядом рисков. Одним из таких рисков является уменьшение национальных генетических ресурсов, или генофондов животных [1].
Анализ полиморфных вариантов генов, отвечающих за молочную продуктивность, предоставляет возможность оценить уровень вариации аллельных форм в стадах крупного рогатого скота и определить степень гетерозиготности. Сохранение этих генов и их низкая степень эволюции у крупного рогатого скота по сравнению с другими плацентарными видами предоставляет возможность использовать их в качестве ДНК-маркеров в селекционном процессе [2–4]. Это особенно актуально при отборе быков-производителей и маточного поголовья по показателям продуктивности. В дальнейшем это позволит сформировать микропопуляции, способные производить продукцию с необходимыми технологическими свойствами [5, 6]. Изучение генетического полиморфизма открывает перспективы для вычисления генетических расстояний и составления генетических карт. Анализ аллельных вариантов генов, ответственных за белковую молочность, также способствует решению задачи сохранения чистоты отечественных молочных пород, особенно на фоне активного процесса голштинизации скота, наблюдаемого в последние десятилетия.
Улучшение отечественных пород сельскохозяйственных животных с целью увеличения объемов производства и удовлетворения потребностей населения зависит от наличия гене- тического разнообразия популяций, а также от применения современных методов селекции. Использование метода ДНК-маркирования способствует выявлению генетических характеристик и ускорению селекционного процесса за счет отбора животных с предпочтительными генотипами [7–9].
Рост объемов валового производства молока достигается благодаря увеличению молочной продуктивности коров, что представляет собой главную тенденцию развития отрасли молочного скотоводства в Российской Федерации [10]. Сокращение поголовья пород отечественной селекции вызывает опасения, так как ценные генетические ресурсы этих животных заключают в себе высокую адаптивность к разным климатическим условиям, равно как и связь с историческими и культурными традициями российских регионов [11–13].
Цель исследования – изучение влияния полиморфизма генов на молочную продуктивность и производственную типичность крупного рогатого скота ярославской породы.
Задачи : изучить распределение генотипов и генетическое разнообразие по генам молочной продуктивности в популяции крупного рогатого скота ярославской породы; оценить молочную продуктивность коров в зависимости от генотипа по генам: синтаза жирных кислот (FASN), тиреоглобулин 5 (TG5), гипофизарный фактор транскрипции 1 (Pit 1), диацилглицерол О-ацил-трансфераза 1 (DGAT 1).
Объекты и методы. Научно-производственный опыт проводился в племенных стадах
Ярославской области в период 2024–2025 гг. Объект исследования – чистопородные коровы-первотелки ярославской породы в количестве 229 голов. Материалом для исследования послужили показатели молочной продуктивности (удой за 305 дней, содержание жира и белка, количество молочного жира и белка), промеры экстерьера (2–4-й месяц первой лактации), биологический материал (цельная кровь).
Исследования по изучению полиморфизма генов проводились в лаборатории генетики и биотехнологии Ярославского НИИЖК – филиала ФНЦ «ВИК им. В.Р. Вильямса».
Для анализа генетического разнообразия крупного рогатого скота изучен полиморфизм генов молочной продуктивности: синтаза жир- ных кислот (FASN), тиреоглобулин 5 (TG5), гипофизарный фактор транскрипции 1 (Pit 1), диа-цилглицерол О-ацилтрансфераза 1 (DGAT 1).
Геномную ДНК выделяли из цельной крови, используя наборы «ДНК-Экстран-1» (ООО «Син-тол», Россия) согласно инструкциям производителя. Полимеразно-цепную реакцию (ПЦР) проводили на термоциклере C1000 Touch (ООО «Био-Рад Лаборатории», Россия). При проведении ПЦР использовали готовую инкубационную смесь 5X ScreenMix-HS (ЗАО «Евроген») с добавлением 0,2 мкМ каждого праймера (ООО «Синтол», Россия).
Праймерные пары для проведения амплификации указаны в таблице 1.
Таблица 1
|
Наименование гена |
Последовательность праймера |
|
Синтаза жирных кислот (FASN) |
F: 5′– AGAGCTGACGGACTCCACAC – 3′ R: 5′– GCCGATGCACTCGATGTAG – 3′ |
|
Тиреоглобулин (TG5) |
F: 5′ – GGGGATGACTACGAGTATGACTG – 3′ R: 5′ – GTGAAAATCTTGTGGAGGCTGTA – 3′. |
|
Гипофизарный фактор транскрипции, Pit1 / HinfI |
F: 5′ – CAATGAGAAAGTTGGTGC – 3′ R: 5′ – TCTGCATTCGAGATGCTC – 3′ |
|
АцилСоАдиацилглицеролацилтрансфераза 1 (DGAT1) |
F: 5' – TGCTGGCCCTGATGGTCTACAC – 3' R: 5' – GAAGGAAGCAAGCGGACAGT – 3' |
Праймеры для амплификации участков генов Primers for amplification of gene regions
Температура отжига праймеров, концентрация ДНК подбирались эмпирически на основе теоретических расчетов. Для проверки и поиска оптимальных температур и времени этапов амплификации проводилась множественная градиентная ПЦР. Диапазон температур для подбора температуры на этапе отжига праймеров составил от 58 до 68 °C, разведения ДНК были кратны 0,5; 1; 2.
Полученные продукты амплификации обрабатывали при необходимости эндонуклеазами рестрикции (ООО «СибЭнзайм», Россия). Срок инкубации составлял не менее 16 ч при температуре 37 ºС в сушильном шкафу Binder ED 115 (BINDER GmbH, Германия). Разделение рестрикционных фрагментов проводили с помощью электрофореза в 2 %-м агарозном геле (ООО «Компания Хеликон», Россия) окрашенным бромистым этидием (ООО «Компания Хеликон», Россия) в электрофорезной горизонтальной ка- мере SubCell GT (ООО «Био-Рад Лаборатории», Россия) при напряжении 120 В с помощью источника питания «Эльф-4» («ООО ДНК-Технология», Россия) в течение 80 мин.
Результаты электрофореза анализировали с помощью гель-документирующей системы Mole-cularImagerGelDocXR (ООО «Био-Рад Лаборатории», Россия).
Для оценки генетического разнообразия рассчитаны показатели на основе формул, представленных в работах Л.А. Животовского (1991) [14] и В.М. Кузнецова (2014) [15].
Количество аллелей – N a .
Количество эффективных аллелей:
Ne = Σ , (1)
где p i – частота i-й аллели.
Информационный индекс Шеннона
КМ = Ж У М ∙ 100 ,
I = -1∙ ∑Pi ∙ ln Pi ,(2)
где p i – частота i-й аллели.
Наблюдаемая гетерозиготность
Ho = гет ,(3)
где N – общее количество особей; N гет – количество особей-гетерозигот.
Ожидаемая гетерозиготность
He =1- ∑Pi2 ,(4)
где pi– частота i-й аллели. Индекс фиксации и (He-Ho)
Fis = ,
где H o – наблюдаемая гетерозиготность; H e – ожидаемая гетерозиготность.
Число степеней свободы
DF =( r -1)∙( c -1) , (6)
Критерий Хи-квадрат
^2 = ∑ r ∑ c ( Oij Eij )
= ∑ ∑ ,
где i – номер строки; j – номер столбца; r – общее количество строк; c – общее количество столбцов; O ij – фактическое количество наблюдений в ячейке ij; E ji – ожидаемое число наблюдений в ячейке ij.
Для оценки статистической значимости различий между эмпирическими и теоретическими частотами встречаемости генотипов применяли критерий Хи-квадрат (χ2) Пирсона [16]. Теоретические частоты встречаемости рассчитывали на основе закона Харди – Вайнберга [14]. Для выявления частных различий эмпирического и теоретического распределений применялся z-тест. Критический уровень значимости принимали равным 0,05 (α = 0,05). Для обработки и статистического анализа данных использовали программное обеспечение: Python 3.9.7, MS Excel, GenAlEx 6.51 [16].
Количество молока, полученного на 100 кг живой массы подопытных коров, – коэффициент молочности (КМ) – определяли по формуле где У – удой коровы за 305 дней первой лактации, кг; ЖМ – живая масса по первой лактации, кг.
Коэффициент производственной типичности (КПТ) определяли по формуле, предложенной Ничиком Б. (1987) [17]:
КПТ = У В ∙ ∙ И И Д С , (9)
где У – удой за 305 дней первой лактации, кг; ИД – индекс длинноногости, %; В – живая масса по первой лактации, кг; ИС – индекс сбитости, %.
Производственную типичность определяли по значению коэффициента следующим образом: КПТ больше 3 – молочное направление; КПТ от 2 до 3 – молочно-мясное направление; КПТ от 1 до 2 – мясо-молочное направление.
Промеры экстерьера определялись по стандартным методикам, биометрическая обработка данных проводилась с использованием «пакета анализа», встроенного в MS Excel, и программы Statistica 10 по методикам Н.А. Плохинского [18].
Результаты и их обсуждение. В результате исследований установлено, что во всех генах наблюдается аллельный полиморфизм, кроме диацилглицерол О-ацилтрансферазы 1, для которого выявлен только один аллель и гомозиготный генотип DGATKK. Аналогичные результаты по отсутствию полиморфизма в гене (DGAT1) установили на коровах белорусской черно-пестрой породы А.Н. Михалюк и Л.А. Танана (2023) [19].
Для гена синтазы жирных кислот наблюдается значительное отклонение от ожидаемых значений: гомозиготный генотип FASNAA встречается реже (4,21 %), чем ожидаемо в равновесном распределении (7,93 %), в то время как гетерозиготный генотип FASNAG и гомозиготный генотип FASNGG встречаются чаще ожидаемого (47,89 % к 40,46 % и 47,89 % к 51,61 % соответственно).
Для гена гипофизарного фактора транскрипции отмечается преобладание гомозиготного генотипа PIT 1/Hinf 1AA (60,56 % к 55,72 %), при этом гетерозиготный генотип PIT 1/Hinf 1AB встречается реже ожидаемого (28,17 % к 37,85 %). Гомозиготный генотип PIT 1/Hinf 1BB также демонстрирует значительное отклонение, встречаясь чаще ожидаемого (11,27 % к 6,43 %).
В случае гена тиреоглобулина наблюдается преобладание гетерозиготного генотипа TG5CT относительно ожидаемого распределения (61,57 % против 47,62 %). Гомозиготный генотип TG5CC встречается реже ожидаемого (30,13 % против 37,11 %), а гомозиготный генотип TG5TT также демонстрирует значительное отклонение, встречаясь реже (8,30 % против 15,27 %).
По-видимому, такое распределение частот генотипов характерно для ярославской породы, поскольку, изучая распределение полиморфных генотипов в популяции татарстанского типа холмогорской породы, Ю.Р. Юльметьевой, Ш.К. Шакировым (2020) было установлено преобладание гомозиготного генотипа TG5CС, частота которого составила 63,5 %. Количество гетерозигот составило 32,1 %, а гомозиготный генотип TG5ТT имел наименьшую частоту встречаемости – 4,4 % [20].
Анализ данных показывает, что наиболее выраженные отклонения от ожидаемых частот наблюдаются для генотипов TG5, PIT 1/Hinf 1 и FASN, это может свидетельствовать о наличии селективного давления на данные аллели в исследуемой популяции. Полученные данные подтверждаются наличием статистически значимых различий между теоретически ожидаемым по закону Харди – Вайнберга и наблюдаемым распределением частот генотипов (табл. 2).
Ген диацилглицерол О-ацилтрансферазы 1 (DAGAT1) характеризуется полным отсутствием генетического разнообразия, что подтверждается значениями числа аллелей ( N a = 1,00), эффективного числа аллелей ( N e = 1,00), информационным индексом Шеннона ( I = 0,00) и наблюдаемой гетерозиготностью ( H o = 0,00), указывающими на наличие только одной аллели и мономорфность гена в исследуемой популяции (табл. 3).
Таблица 2
Анализ соответствия распределения частот генотипов теоретически ожидаемому равновесному распределению Харди – Вайнберга
Analysis of the correspondence of the genotype frequency distribution to the theoretically expected Hardy-Weinberg equilibrium distribution
|
Ген |
Число степеней свободы DF |
Значение критерия Хи-квадрат χ2 |
Достигнутый уровень значимости p-value |
|
Ген диацилглицерол О-ацилтрансферазы 1 |
– |
– |
– |
|
Синтаза жирных кислот |
1 |
6,419 |
0,011 |
|
Гипофизарный фактор транскрипции |
1 |
9,289 |
0,002 |
|
Тиреоглобулин |
1 |
19,671 |
0,000 |
Ген синтазы жирных кислот (FASN) имеет умеренное генетическое разнообразие с двумя аллелями, эффективным числом аллелей 1,68 и информационным индексом Шеннона, равным 0,59. Наблюдаемая гетерозиготность ( H o = 0,48) незначительно превышает ожидаемую ( H e = 0,40), что подтверждается отрицательным значением индекса фиксации ( Fis = –0,18) (табл. 3).
Гипофизарный фактор транскрипции (PIT1) также имеет два аллеля, но более низкие эффективное число аллелей (Ne = 1,61) и информационный индекс Шеннона (I = 0,57). При этом наблюдается существенное отклонение наблюдаемой гетерозиготности (Ho = 0,28) от ожидае- мой (He = 0,38), что подтверждается положительным значением индекса фиксации (Fis = 0,26) и говорит о дефиците гетерозигот по этому гену (см. табл. 3).
Наиболее высоким генетическим разнообразием характеризуется ген тиреоглобулина (TG5), имеющий два аллеля, эффективное число аллелей 1,91 и информационный индекс Шеннона I = 0,67. Наблюдаемая гетерозиготность ( H o = 0,62) значительно превышает ожидаемую ( H e = 0,48), что подтверждается отрицательным значением индекса фиксации ( F is = –0,29) (см. табл. 3).
Таблица 3
Показатели генетического разнообразия популяции крупного рогатого скота ярославской породы
Indicators of the genetic diversity of the Yaroslavl cattle population
|
Показатель |
Ген диацилглицерол О-ацилтрансферазы 1 |
Синтаза жирных кислот |
Гипофизарный фактор транскрипции |
Тиреоглобулин |
|
Объем выборки N |
70 |
190 |
142 |
229 |
|
Число аллелей N a |
1,00 |
2,00 |
2,00 |
2,00 |
|
Эффективное число аллелей N e |
1,00 |
1,68 |
1,61 |
1,91 |
|
Информационный индекс Шеннона I |
0,00 |
0,59 |
0,57 |
0,67 |
|
Наблюдаемая гетерозиготность H o |
0,00 |
0,48 |
0,28 |
0,62 |
|
Ожидаемая гетерозиготность H e |
0,00 |
0,40 |
0,38 |
0,48 |
|
Индекс фиксации Fis |
– |
-0,18 |
0,26 |
-0,29 |
Оценка взаимосвязи генотипов по изученным генам с молочной продуктивностью коров показала: животные с генотипом FASNAG достоверно превосходили гомозиготных животных с генотипом FASNGG по надою за 305 дней первой лак- тации на 639 кг (p ≤ 0,001) молока (табл. 4). По содержанию молочного жира достоверное превосходство также выявлено у первотелок с генотипом FASNAG над животными с генотипом FASNAA 0,29 % (p ≤ 0,05): оно составило 4,54 %.
Таблица 4
Молочная продуктивность коров по первой лактации в зависимости от генотипа (M±m) Milk productivity of cows after the first lactation, depending on the genotype (M±m)
|
Признак |
FASN |
TG5 |
Pit 1/Hinf |
||||||
|
AA |
AG |
GG |
TT |
CT |
CC |
AA |
AB |
BB |
|
|
Удой за 305 дней, кг |
5700± |
5699± |
5060± |
5758± |
5549± |
5172± |
5941± |
5816± |
5437± |
|
517,3 |
144,9*** |
115,0 |
332,7 |
114,7* |
103,5 |
157,9 |
269,7 |
343,9 |
|
|
МДЖ, % |
4,25± |
4,54± |
4,48± |
4,36± |
4,46± |
4,37± |
4,41± |
4,34± |
4,48± |
|
0,14 |
0,06* |
0,06 |
0,10 |
0,04 |
0,07 |
0,04 |
0,06 |
0,08 |
|
|
Количество |
243,4± |
257,7± |
226,2± |
250,5± |
247,1± |
226,3± |
262,0± |
255,5± |
243,2± |
|
молочного жира, кг |
24,7 |
6,7*** |
24,7 |
14,6 |
5,4* |
6,0 |
7,4 |
12,7 |
15,5 |
|
МДБ, % |
3,40± |
3,27± |
3,27± |
3,31± |
3,32± |
3,20± |
3,26± |
3,33± |
3,32± |
|
0,06* |
0,02 |
0,02 |
0,06 |
0,02*** |
0,02 |
0,02 |
0,04 |
0,05 |
|
|
Количество |
194,3± |
187,4± |
166,4± |
191,1± |
184,5± |
165,7± |
194,3± |
197,9± |
181,6± |
|
молочного белка, кг |
18,2 |
5,4** |
4,3 |
11,7* |
4,2*** |
3,6 |
5,7 |
10,9 |
12,7 |
|
Живая масса, кг |
519± |
499± |
476± |
477± |
494± |
474± |
501± |
509± |
487± |
|
21,5 |
5,4*** |
3,4 |
8,7 |
4,1*** |
4,3 |
5,3 |
10,7 |
15,7 |
|
|
Коэффициент |
1094,0± |
1219± |
1104,0± |
1376,0± |
1169,0± |
1175,0± |
1268± |
1170,0± |
1123,0± |
|
молочности, кг |
92,1 |
32,7** |
28,2 |
133,7 |
25,0 |
32,6 |
39,2* |
54,7 |
58,4 |
По комплексному выходу молочного жира и белка на 100 кг живой массы также особи с генотипом FASNAG превосходили показатели гомо- зигот с генотипом FASNGG на 7,5 кг (p ≤ 0,01) (см. табл. 4).
Особи с генотипом FASNAG произвели больше молока на 100 кг живой массы по сравнению АА
FASNGG на составил 1
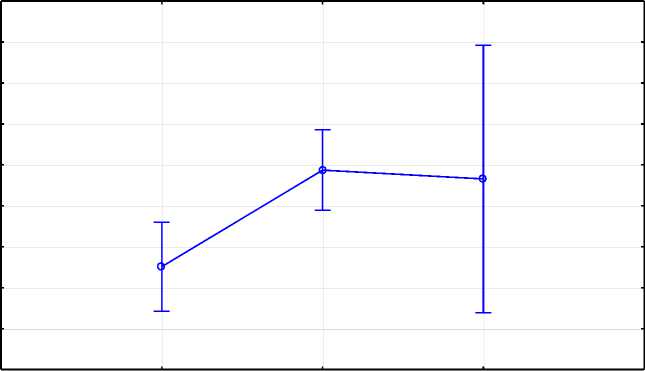
По производственной типичности коровы всех генотипов FASN относились к молочному типу м FASNAG p ≤ 0,01). SNAG 4,3, FASNaa - 4,3; FASNGG - 3,9 (рис. 1).
5,2
5,0
4,8
4,6
4,4
4,2
4,0
3,8
3,6
3,4
GG
AG
AA
FASN
Рис. 1. Расчет различий средних методом LSMeans по гену FASN Calculation of differences in averages using the LS Means method for the FASN gene
Установлена статистически значимая разница между показателями коэффициента производственной типичности у особей с генотипами FASNAG и FASNGG (p = 0,0018) (рис. 1).
При оценке продуктивных показателей коров, генотипированных по гену TG5, была выявлена тенденция превосходства особей с гетерозиготным генотипом TG5СТ по качественным показателям молока (см. табл. 4). Так, по содержанию жира показатель гетерозигот равнялся 4,46 %, превосходство над гомозиготами TG5ТТ и TG5СС составило 0,10 и 0,09 абс. % соответственно. По содержанию массовой доли белка животные с генотипом TG5СТ превосходили показатели гомозигот TG5СС на 0,12 абс. % (p ≤ 0,01).
Изучая взаимосвязь продуктивных признаков с полиморфизмом гена TG5, Е.О. Крупин и Ш.К. Шакиров (2019) выявили существенные различия по содержанию массовой доли жира в молоке у коров с генотипами CC и TC (на 0,29 %) [21].
По удою за 305 дней первой лактации, количеству молока на 100 кг живой массы гомозиготы TG5ТТ превосходили особей с генотипами
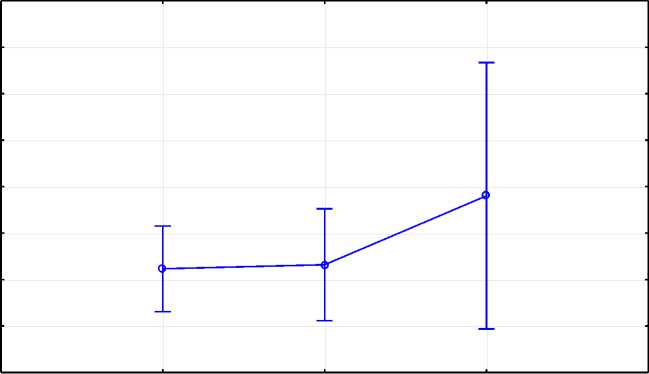
TG5СТ и TG5СС. Это подтверждается и значением коэффициента производственной типичности, первотелки с генотипом TG5ТТ имели более молочный тип (КПТ = 4,6), однако разница не была статистически значима (р = 0,3472) (рис. 2).
У животных, генотипированных по гену Pit 1/Hinf, достоверных различий по показателям молочной продуктивности установить не удалось. Однако проведенный анализ показал, что особи с генотипом Pit 1АА превосходили показатели животных с генотипами Pit 1АВ и Pit 1ВВ по удою за первую лактацию на 125 и 504 кг молока соответственно, по общему выходу молочного жира и белка – на 2,9 и 31,6 кг соответственно, по общему выходу молочного жира и белка на 100 кг живой массы – на 3,4 и 8,5 кг соответственно. Также от коров с гомозиготным генотипом Pit 1АА получено больше молока на 100 кг живой массы (КМ = 1268,0 кг) по сравнению с особями с генотипом Pit 1АВ (КМ = 1170,0 кг) и Pit 1ВВ (КМ = 1123,0 кг), т. е. они имели лучшую конверсию корма.
5,4
5,2
5,0
4,8
4,6
4,4
4,2
4,0
3,8
СТ
CC
TT
TG5
Рис. 2. Расчет различий средних методом LSMeans по гену TG5 Calculation of differences in averages by the LS Means method for the TG5 gene
Аналогичную зависимость молочной продуктивности от генотипа Pit 1АА установили М.В. Позовникова и Г.Н. Сердюк (2017) у помесных коров черно-пестрой и голштинской пород Лен
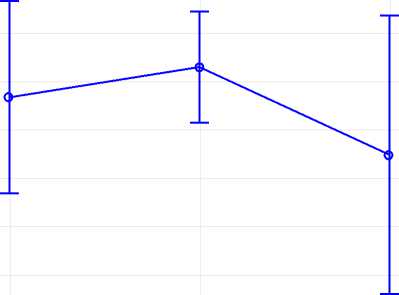
Также гомозиготы Pit Iм имели более мо-лоч ниц
(КПТ = 4,1) (р АА–АВ = 0,1534; р АА–ВВ = 0,1231) (рис. 3).
Т.Ф. Лефлер, И.В. Крашенинникова (2022) сообщают о влиянии генотипа коров на молоч-ос-,6– рта т о соответствии коров молочному типу [23].
5,0
>s о и я о Я сп S о
4,8
4,6
4,4
s
и о
S я
S
о и S’ S к
S
4,2
4,0
3,8
3,6
о
3,4
3,2
AB
AA
BB
Pit 1/Hinf
Рис. 3. Расчет различий средних методом LSMeans по гену Pit 1 Calculation of differences in averages by the LS Means method for the Pit 1 gene
Заключение. Исследование генетического разнообразия крупного рогатого скота ярославской породы выявило существенные различия между анализируемыми генами. Наиболее важным результатом является полное отсутствие полиморфизма в гене DAGAT1, что может свидетельствовать о его высокой консервативности или действии направленного стабилизирующего селекционного отбора. Ген тиреоглобулина проявляет максимальное генетическое разнообразие ( Fis = –0,29), что указывает на преобладание гетерозигот и аутбредный способ подбора родительских пар. Умеренное разнообразие FASN ( Fis = –0,18) указывает на его значимость в метаболических процессах, а особенности распределения PIT1 ( Fis = 0,26) могут быть связаны с его функцией в регуляции транскрипции. Полученные данные имеют важное значение для понимания генетической структуры породы и могут быть использованы при планировании селекционной работы.
На основании оценки молочной продуктивности коров по первой лактации установлено, что
Вестник КрасГАУ. 2025. № 10 (223) наибольшее статистически значимое влияние на удой молока оказывает аллельный полиморфизм гена FASNAG. От них получено на 639 кг молока больше (p ≤ 0,001) относительно гомозигот FASNGG. Коровы с генотипом TG5СТ имели больший удой по сравнению со сверстницами с генотипом TG5СС на 377 кг молока (p ≤ 0,05).
Качественные показатели молока в исследуемых стадах были в наибольшей степени обусловлены полиморфизмом генов FASN и TG5: первотелки с генотипом FASNAG превосходили по массовой доле жира животных с генотипом FASNAA на 0,29 % (p ≤ 0,05); животные с генотипом TG5СТ превосходили показатели гомозигот TG5СС по массовой доле белка на 0,12 % (p ≤ 0,001).
Наибольший коэффициент молочности установлен у особей с генотипом FASNAG и Pit 1АА – 1219 и 1268 кг молока на 100 кг живой массыЪ соответственно. Согласно значению коэффициента производственной типичности коров с генотипами FASNAGTG5ТТ и Pit 1АА можно отнести к высокомолочному типу.
