Кости животных из ранних комплексов многослойного поселения Саган-Заба II (9 120-7 880 кал. л.н.) в Прибайкалье: планиграфия, хозяйственная деятельность и сезонность использования стоянки
 в Прибайкалье: планиграфия, хозяйственная деятельность и сезонность использования стоянки")
Автор: Номоконова Т.Ю., Горюнова О.И., Лозей Р.дЖ., Новиков А.Г., Вебер А.В.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Палеоэкология. Каменный век
Статья в выпуске: 3 т.44, 2016 года.
Бесплатный доступ
Статья является первой публикацией, посвященной анализу костей животных из ранних комплексов многослойного поселения Саган-Заба II (VII и VI слои), расположенного на западном побережье оз. Байкал. Рассматриваются вопросы видового состава фауны, радиоуглеродное датирование костей животных, планиграфия их нахождения в слоях, определение пола и возраста животных, их выбор и сезонность использования стоянки. По сравнению с предыдущими работами по датированию слоев памятника Саган-Заба II в данном исследовании учитывается ранее определенная возможная разница между некалиброванными датами, полученными по костным остаткам копытных и нерпы, в связи с проблемой «старого» углерода в оз. Байкал. Для VII слоя она в среднем составляет 682 года, а для VI - 509 лет. С учетом пересчета дат, полученных по костям нерпы, VII и VI слои теперь датируются в промежутке между 9 120 и 7 880 кал. л.н. Анализ фаунистических материалов из ранних комплексов поселения Саган-Заба II показал, что население занималось охотой на нерпу, копытных и других млекопитающих, а также на птиц и рыбной ловлей, что говорит о комплексном использовании природных ресурсов. Вероятно, исследуемые стоянки носили кратковременный, сезонный характер, что подтверждается особенностью размещения очагов и скоплений материалов, незначительной мощностью кострищ, отсутствием специализированных производственных площадок. О сезонном их использовании (преимущественно в весеннее и летнее время года) свидетельствуют результаты анализа слоев дентина на срезах клыков нерпы и наличие костей животных, присутствие которых в Прибайкалье ограничено теплыми месяцами года.
Голоцен, радиоуглеродное датирование
Короткий адрес: https://sciup.org/145145770
IDR: 145145770 | УДК: 904(571.53) | DOI: 10.17746/1563-0102.2016.44.3.037-046
Faunal remains from the early complexes of the Sagan-Zaba II multilayered habitation site (9120-7880 cal. bp) in the Cis-Baikal: planigraphy, subsistence patterns, and seasonality
This article is the first publication to analyze faunal remains from early complexes (layers VII and VI) at the multilayer settlement of Sagan-Zaba II, situated on the western shore of Lake Baikal. We discuss species composition of fauna from the site as well as associated radiocarbon dates, age and sex designations, spatial distribution, and their overall selection as it relates to site seasonality. We address the previously-defined potential offset between uncalibrated dates from remains of ungulates and seals at the site, relating to the problem of "old" carbon in Lake Baikal. For layer VII, this offset is 682 years on average, and for layer VI it is 509 years. Taking the offset from nerpa bones into account, layers VII and VI now appear to date to the period between 9120 and 7880 cal BP. An analysis of faunal materials from the early complexes of the Sagan-Zaba II settlement indicates that inhabitants of this site hunted nerpa seals, ungulates, and other mammals as well as birds and fish, demonstrating a complex use of natural resources. Most likely, sites featured short-term, seasonal occupations, as indicated by the distribution of hearths and other materials, the thickness of hearth features, and the absence of specialized production areas. Results of an analysis of dentine layers from cross-sections of nerpa teeth as well as the presence of faunal bones that are only available during the hotter months of the year in the Cis-Baikal region, are consistent with a model of spring and summer site use.
Текст научной статьи Кости животных из ранних комплексов многослойного поселения Саган-Заба II (9 120-7 880 кал. л.н.) в Прибайкалье: планиграфия, хозяйственная деятельность и сезонность использования стоянки
Многослойное поселение Саган-Заба II представлено несколькими комплексами, характеризующими древние культуры Прибайкалья в голоцене. Время существования стоянок определено на основании серийного радиоуглеродного датирования в интервале от ~9 000 до 900 кал. л.н. [Nomokonova et al., 2013]. Многообразные археологические и фаунистические материалы памятника предоставляют возможность проследить особенности хозяйственного освоения Приольхонья в разные хронологические срезы. Археологические комплексы ряда культурных слоев уже введены в научный оборот [Долганов и др., 2011, 2013; Горюнова и др., 2012; Новиков, Горюнова, Вебер, 2014; Nomokonova et al., 2010]. Однако многочисленные фаунистические остатки, полученные в ходе раскопок северо-восточной части бухты, и хозяйственные аспекты ранее не рассматривались. Предлагаемая статья является первой публикацией, посвященной анализу костей животных из ранних комплексов этой части поселения Саган-Заба II (VII и VI слои раскопов
2007–2008 гг.), включая видовой состав фауны, радиоуглеродное датирование костей животных, планиграфию их нахождения в слоях, выбор животных и сезонность использования данной стоянки.
Памятник находится в бухте Саган-Заба западного побережья оз. Байкал (рис. 1), в 154 км к северо-востоку от г. Иркутска и в 13,5 км к юго-востоку от пос. Еланцы (Ольхонский р-н Иркутской обл.). Он открыт отрядом Североазиатской экспедиции ИИФФ СО АН СССР, возглавляемым А.П. Окладниковым, в 1972 г. Раскопки были проведены тем же отрядом под руководством А.П. Окладникова и И.В. Асеева в 1974–1975 гг. Выделено пять культурных слоев [Окладников, 1975; Асеев, 2003, с. 51]. Раскопки, направленные на комплексное, междисциплинарное исследование стоянки, возобновились в 2006–2008 гг. Саган-Забинским отрядом Российско-Канадской археологической экспедиции (Иркутская лаборатория археологии и палеоэкологии ИАЭТ СО РАН – ИГУ и Отдел антропологии Университета Альберты, г. Эдмонтон, Канада) [Горюнова, Новиков, Воробьева и др., 2007; Горюнова, Новиков, Вебер и др., 2008].
Рис. 1. Карта-схема расположения стоянки Саган-Заба II на побережье оз. Байкал.
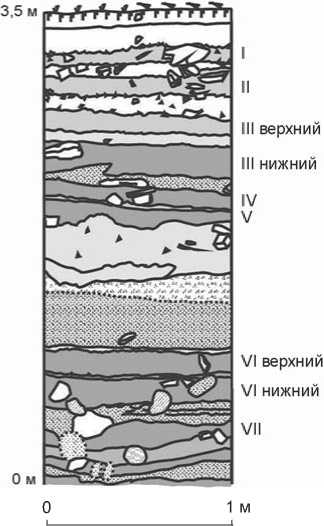
Рис. 2. Стратиграфический разрез северо-западной стенки раскопа 4с (кв. 4а).
В результате работ в северо-восточной части бухты (наиболее перспективной для раскопок) выделено 11 культурных слоев (с VII по I, с учетом подразделения некоторых из них на нижний и верхний).
Стратиграфический разрез в месте раскопов представляет собой серию гумусированных супесчаных почв, разделенных прослоями светлой щебенистой супеси и грубообломочных отложений пролювиальноделювиального генезиса (рис. 2) [Горюнова, Новиков, Вебер и др., 2008; Воробьева, 2010]. Комплексы VII, VI нижнего и верхнего культурных горизонтов стратиграфически привязаны к слоям гумусированной супеси серовато-бурого (до бурого) и темного серовато-бурого цвета мощностью 0,12–0,18 м. Их деление хорошо выражено на фронтальной стенке раскопа (в глубь абразионного уступа); у берегового обрыва толщина слоев уменьшается до 0,05 м (рис. 3).
Методика полевых исследований включала послойное вскрытие раскопов, тщательное просеивание отработанной почвы всех культурных отложений через сито с ячеей 3 мм, фиксирование материала электронным теодолитом по трехмерным показателям, детальное стратиграфическое изучение разрезов, отбор образцов для исследования естественно-научными методами.
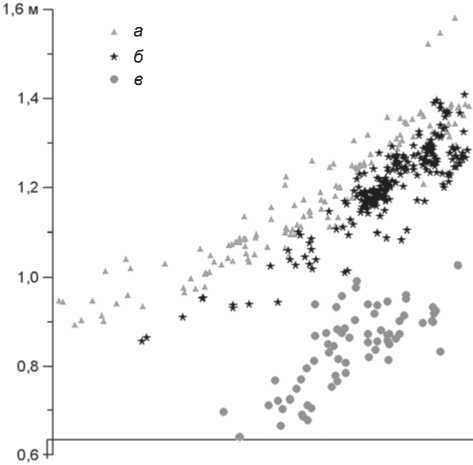
Рис. 3. Двухмерный вертикальный разрез раскопа 4с с плотностью археологического материала, найденного в VII и VI слоях.
а – VI верхний слой; б – VI нижний; в – VII слой.
Радиоуглеродное датирование костей животных
Хронология VII, VI нижнего и верхнего слоев стоянки Саган-Заба II определена с помощью 16 радиоуглеродных AMS-дат (табл. 1), калиброванных с ис- пользованием программы Calib 7.0.1 [Stuiver, Reimer, 1993] по базе данных IntCal13 [Reimer et al., 2013]. Даты получены по костям копытных и нерп, отобранных в одинаковом количестве из каждого анализируемого слоя, и представлены по калиброванной кри-
Таблица 1 . Результаты радиоуглеродного датирования костей животных
|
Номер образца |
Таксон |
14C-дата |
Дата с учетом «старого» углерода |
Калиброванные значения л.н. |
ETI (68 % вероятности) |
|
VII слой |
|||||
|
OxA22387 |
Cervus elaphus |
8 024 ± 40 |
– |
9 020–8 730 |
1–257 |
|
OxA22363 |
Capreolus pygargus |
8 010 ± 40 |
– |
9 010–8 730 |
|
|
OxA22364 |
» |
7 986 ± 37 |
– |
9 000–8 660 |
|
|
OxA22358 |
Artiodactyla |
7 971 ± 39 |
– |
9 000–8 650 |
|
|
OxA22411 |
Phoca sibirica |
8 745 ± 45 |
8 063 ± 45 |
9 120–8 770 |
1–244 |
|
OxA22419 |
» |
8 705 ± 40 |
8 023 ± 40 |
9 020–8 720 |
|
|
OxA22412 |
» |
8 680 ± 40 |
7 998 ± 40 |
9 010–8 660 |
|
|
OxA22421 |
» |
8 600 ± 40 |
7 918 ± 40 |
8 980–8 610 |
|
|
VI нижний и верхний слои |
|||||
|
OxA22357 |
Artiodactyla c.f. |
7 203 ± 37 |
– |
8 160–7 950 |
1–79 |
|
OxA22390 |
Artiodactyla |
7 188 ± 36 |
– |
8 150–7 940 |
|
|
OxA22356 |
» |
7 179 ± 36 |
– |
8 100–7 940 |
|
|
OxA22374 |
Cervidae c.f. |
7 147 ± 38 |
– |
8 020–7 880 |
|
|
OxA20578 |
Phoca sibirica |
7 924 ± 40 |
7 415 ± 40 |
8 340–8 170 |
1–222 |
|
OxA20580 |
» |
7 881 ± 37 |
7 372 ± 37 |
8 320–8 050 |
|
|
OxA22420 |
» |
7 865 ± 40 |
7 356 ± 40 |
8 310–8 040 |
|
|
OxA20579 |
» |
7 835 ± 37 |
7 326 ± 37 |
8 200–8 020 |
|
OxA22411_Phoca
OxA22387_Cervus
OxA22419_Phoca
OxA22363_Capreolus
OxA22412_Phoca
OxA22364_Capreolus
OxA22358_Artiodactyla
OxA22421_Phoca
OxA20578_Phoca
OxA20580_Phoca
OxA22420_Phoca
OxA20579_Phoca
OxA22357_Artiodactyla
OxA22390_Artiodactyla
OxA22356_Artiodactyla
OxA22374_Cervidae
9 500 9 000 8 500 8 000 кал. л.н.
вой на 95 % вероятности. В предыдущих публикациях мы использовали только результаты датирования образцов копытных, т.к. кости нерп содержат в себе «старый» углерод Байкала, в связи с чем радиоуглеродные даты, полученные по ним, удрев-нены на несколько столетий (подробно см.: [Nomokonova et al., 2013]). В данном исследовании учтена возможная разница между некалиброванными датами, полученными по ко стным остаткам копытных и нерп. Для VII слоя она составляет 682 года, для VI – 509 лет. С учетом пересчета и использования дат, полученных по костям нерпы, культурные остатки VII слоя датируются в интервале 9 120–8 610 кал. л.н., VI – 8 340–7 880 кал. л.н. (табл. 1; рис. 4).
В связи с тем, что для каждого слоя получено несколько радиоуглеродных дат, стало возможным использовать дополнительную функцию, доступную в программе BCAL [Buck, Christen, James, 1999],
Рис. 4. Радиоуглеродные даты, полученные по костям животных из VII и VI слоев.
под названием ETI (elapsed time intervals). Она позволяет в среднем прикинуть вероятность перекрывающихся временных интервалов по полученному набору дат, чтобы предположить, к какому промежутку времени относятся эти даты и когда могла использоваться стоянка. В табл. 1 приведены такие показатели, рассчитанные для дат, которые получены по костям копытных и нерп из каждого анализируемого слоя. Они демонстрируют, что, вероятно, промежуток времени, когда эти животные использовались на стоянке, более или менее одинаковый, за исключением копытных, остатки которых найдены в VI слое, где этот показатель меньше, до 80 лет.
Контекст нахождения костей животных
Фаунистические остатки в VII и VI слоях стоянки Саган-Заба II располагались преимущественно скоплениями в районах очагов, кострищ и каменных конструкций. Например, в VII слое зафиксированы одно кострище и шесть каменных выкладок, занимающих значительную часть вскрытой площади (рис. 5). Вероятно, все они служили очажными сооружениями. Конструкции овальной формы, кольцевые, как состоящие из разрозненных камней, так и сплошные. Размеры кладок 0,92÷0,34 × 0,71÷0,28 м. Мощность зольников незначительная. Археологический материал VII слоя располагался скоплениями в районе очагов. В основном это фаунистические остатки. Совместно с ними залегали каменные и костяные изделия: призматические пластины, резчик, скобель, резцы, провертка, вкладыши и обломки костяных обойм составных орудий, гарпун и фрагменты расщепленного клыка кабана (рис. 6, 3–7). Состав скоплений позволяет характеризовать их как хозяйственно-бытовые отбросы.
Планиграфическое размещение костей животных в VI нижнем и верхнем слоях в целом подтверждает контекст расположения находок в районах очагов и каменных конструкций. В отличие от предыдущего слоя, в этих комплексах зафиксированы фрагменты керамики (более чем от 17 сосудов). Археологический материал VI нижнего слоя был сконцентрирован тремя скоплениями, аналогичными по составу и типологическому набору инвентаря [Горюнова и др., 2012]. Все они группировались в районе очага (к востоку и западу от него). Очажная кладка полукольцевая, овальной формы, размерами 1,1 × 0,8 м. Мощность зольника небольшая. В составе скоплений преобладают фаунистические остатки, представлены фрагменты керамики от нескольких сосудов, призматические пластины, вкладыши, резцы, стерженек составного рыболовного крючка, обломки костяной иглы (рис. 6, 10 )
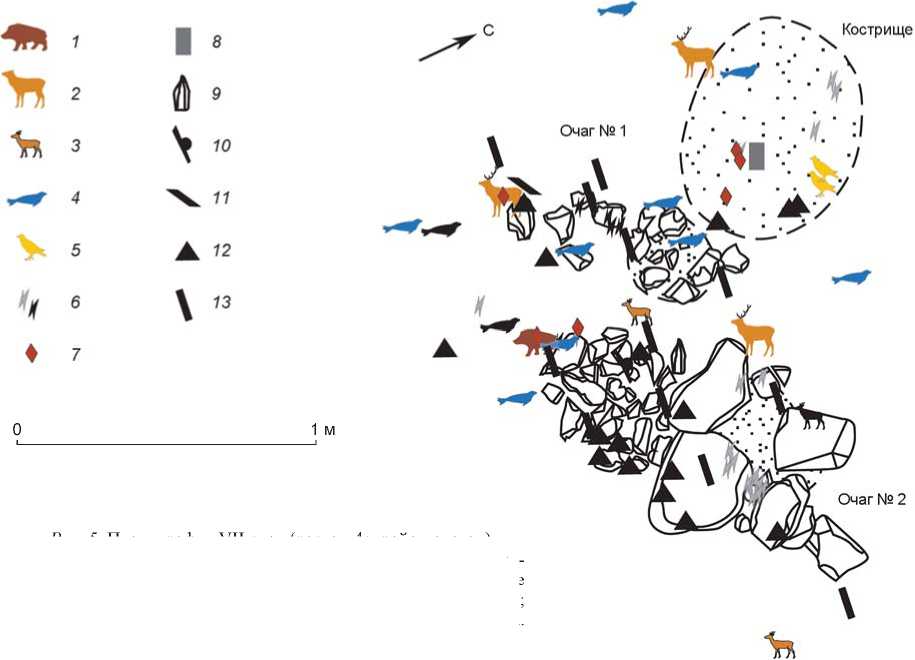
Рис. 5. Планиграфия VII слоя (раскоп 4с, район очагов).
1 – кости кабана; 2 – костные остатки крупных оленьих; 3 – кости мелких оленьих; 4 – костные остатки нерпы; 5 – кости птиц; 6 – жженые неопределимые кости; 7 – костяные изделия; 8 – абразив; 9 – нуклеус; 10 – скребок; 11 – резец; 12 – отщеп, скол; 13 – призматическая пластина.
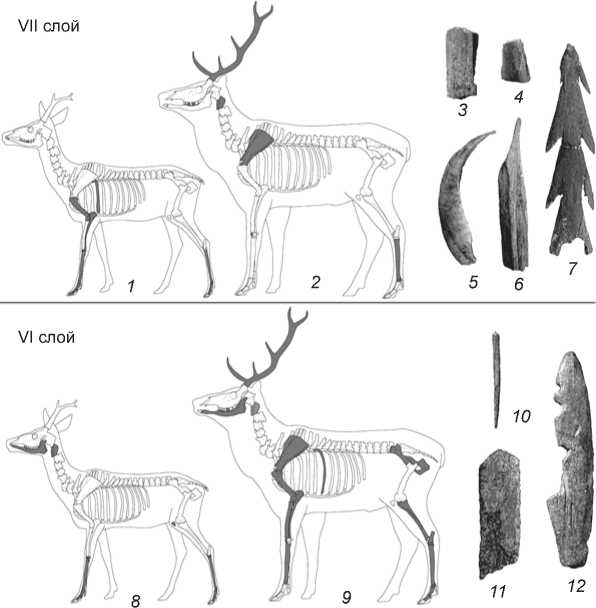
Рис. 6. Элементы скелета копытных (отмечены черным цветом) и орудия из кости.
и перламутровых бусинок. В месте одного скопления отмечена золистая почва.
В VI верхнем слое отмечено семь каменных конструкций, которые, возможно, служили очажными кладками [Там же]. Встречены полукольцевые и сплошные сооружения овальной формы, размерами 1,6÷0,8 × 1,2÷0,5 м. В некоторых из них зафиксированы размытые зольники. Возле очагов обнаружены фаунистические остатки, фрагменты керамики от разных сосудов, призматические пластины и вкладыши. Основная часть археологического материала VI верхнего слоя приурочена к скоплению, тяготеющему к береговой части раскопа. Почва в этом месте золистая. Скопление также составляли фаунистические остатки, фрагменты керамики от нескольких сосудов, каменные изделия (призматические пластины, вкладыши, резец, тесловидное орудие), гарпун и костяная обойма составного орудия (рис. 6, 11, 12 ). Набор и типология керамических и каменных изделий, найденных в VI нижнем и верхнем слоях, аналогичны, что дает основание говорить о формировании комплексов в рамках единой культурной традиции, в одном хронологическом срезе [Там же].
Количество очажных кладок, обнаруженных в VII и VI слоях, их близкое расположение между собой позволяют предположить, что система каменных кон- струкций отражает несколько по сле-довательных разносезонных ситуаций, а сравнительно незначительная мощность зольников свидетельствует о кратковременном использовании стоянок.
Судя по залеганию ко стей животных в очагах и кострищах, а также возле них, туши могли быть обработаны и частично употреблены непосредственно на стоянке. Это подтверждается следами на некоторых костях. В материалах VII слоя кости с порезами, жженые, с погрызами, а также костяные орудия составляют 299 экз., или 28 % от общего количества фаунистических остатков из этого слоя. Они принадлежат преимущественно млекопитающим. Преобладают жженые кости – 293 экз. (в т.ч. 15 – нерпы, несколько – парнокопытных, одна – птицы), что может ассоциироваться непосредственно с очагами и ко стрищами. Кости со следами обработки и в виде орудий представлены 10 экз. На одной трубчатой кости обнаружены погрызы хищников.
В комплексах VI слоя кости со следами обработки представлены 64 экз., что составляет 5 % от общего количества фаунистических остатков из это- го слоя. Среди них также преобладают жженые кости (53 экз., в т.ч. 15 – нерпы). Порезы обнаружены на большой берцовой кости нерпы, плечевой кости благородного оленя, ребре парнокопытного и неопределимом фрагменте ко сти млекопитающего. Кроме того, найдены пять костей с подработкой и в виде изделий, а также две с погрызами хищников. Наличие последних может указывать на то, что кости животных на стоянке поедались и хищниками, но, поскольку такие находки единичные, эти действия могут рассматриваться как минимальные.
Видовой состав животных
Фаунистические материалы, найденные в результате раскопок VII и VI слоев, представлены 2 351 экз. костей и зубов животных (табл. 2). Большинство из них принадлежат млекопитающим – 94 % от общего количества фаунистических остатков. Видовые определения в этой категории возможны по следующим таксономическим уровням (табл. 2): вид (косуля, благородный олень, лось, кабан и нерпа), семейство (оленьи с подразделением на крупных и средних представителей), отряд (парнокопытные и хищники) и класс (крупные и средние млекопитающие). Осталь-
Таблица 2 . Фаунистические материалы
|
Таксон |
Название |
VII слой |
VI слой |
Всего |
||
|
Кол-во костей |
Число особей min |
Кол-во костей |
Число особей min |
|||
|
Mammalia |
Млекопитающие |
711 |
886 |
1 597 |
||
|
Mammalia крупные |
31 |
28 |
59 |
|||
|
Mammalia средние |
30 |
15 |
45 |
|||
|
Artiodactyla |
Парнокопытные |
7 |
1 |
1 |
8 |
|
|
Artiodactyla крупные |
– |
– |
4 |
4 |
||
|
Artiodactyla средние |
5 |
5 |
10 |
|||
|
Cervidae |
Сем. оленьих |
– |
– |
4 |
2 |
4 |
|
Cervidae крупные |
5 |
1 |
11 |
2 |
16 |
|
|
Cervus elaphus |
Благородный олень |
19 |
1 |
13 |
2 |
32 |
|
Alces alces |
Лось |
– |
– |
1 |
1 |
1 |
|
Cervidae средние |
7 |
1 |
7 |
2 |
14 |
|
|
Capreolus pygargus |
Косуля |
9 |
2 |
10 |
2 |
19 |
|
Sus scrofa |
Кабан |
2 |
1 |
2 |
1 |
4 |
|
Carnivora средние |
Хищники |
1 |
1 |
1 |
1 |
2 |
|
Phoca sibirica |
Нерпа |
101 |
5 |
296 |
9 |
397 |
|
Aves |
Птицы |
3 |
– |
– |
3 |
|
|
Anatidae |
Сем. утиных |
2 |
1 |
– |
– |
2 |
|
Phalacrocax carbo |
Баклан |
9 |
2 |
– |
– |
9 |
|
Haliaeetus sp. |
Род орланов |
1 |
1 |
1 |
1 |
2 |
|
Corvus c.f. corax |
Ворон |
– |
– |
1 |
1 |
1 |
|
Pisces неопред. |
Рыбы |
107 |
1 |
1 |
108 |
|
|
Thymallus articus |
Хариус |
3 |
2 |
– |
– |
3 |
|
Acipenser baeri baic. |
Осетр |
– |
– |
1 |
1 |
1 |
|
Mollusca |
Раковины |
– |
– |
6 |
6 |
|
|
Неопределимые |
4 |
– |
– |
4 |
||
|
Всего |
1 057 |
1 294 |
2 351 |
|||
ная фауна представлена 17 костями птиц и 112 остатками рыб, а также несколькими фрагментами раковин (табл. 2).
Из костных остатков птиц найдены посткраниальные элементы скелетов минимум двух бакланов, коракоид ворона, плечевая кость и коракоид птицы рода орланов, фаланга и фрагмент вилочки представителя семейства уток и неопределимые выше уровня класса остатки позвонков. Несколько элементов черепа рыб были отнесены минимум к двум особям хариуса и осетру. Другие остатки рыб, представленные чешуей, ребрами и фрагментами черепа, неопределимы.
Большинство фаунистических остатков принадлежит байкальской нерпе – 17 % от общего количества костей или 75 %, если учитывать только фауну, определимую до уровня отряда. Остатки этого вида принадлежат минимум 14 особям. Представлены элемен- ты всех отделов скелета без какого-либо преобладания костей тех или иных частей туш животных. Вторую наиболее значимую группу составляют парнокопытные – 4,8 % от общего количе ства фаунистических остатков или 21 %, если учитывать только фауну, определимую до уровня отряда. Здесь преобладают кости косули и благородного оленя; также отмечены фрагмент лучевой кости лося и костные остатки кабана (фрагмент черепа, вторая фаланга и расщепленный клык).
Средние и крупные парнокопытные представлены элементами разных отделов скелета, но в основном костями конечностей и головы с фрагментами рогов (рис. 6, 1, 2, 8, 9). Насколько такая ситуация характерна для отбора определенных частей туш этих животных, остается неясным, т.к. среди костных остатков млекопитающих часто встречаются и элементы дру- гих отделов скелета, например, обломки ребер и позвонков, фрагментарность которых затрудняет их видовое определение.
Определение пола и возраста животных
В связи с тем, что кости и зубы нерпы, а также парнокопытных преобладают среди фаунистиче ских остатков на памятнике Саган-Заба II, были проведены дополнительные исследования для определения пола и возраста некоторых из этих животных. Возраст нерп определялся по двум методикам. Первая основана на отнесении индивидуальных элементов скелета к обобщенным возрастным категориям по степени прирастания проксимальных и дистальных эпифизов на костях (в основном конечностей) [Storå, 2000], вторая – на подсчете годовых структур в дентине на срезах клыков. Последняя была разработана и ранее применялась для анализа клыков нерп с ряда многослойных стоянок Прибайкалья [Weber, Goriunova, Konopatskii, 1993; Weber et al., 1998]. В отличие от предыдущих работ, заключительные подсчеты выполнялись с учетом минимального количества нерп для исключения повторного использования одних и тех же особей при интерпретации результатов.
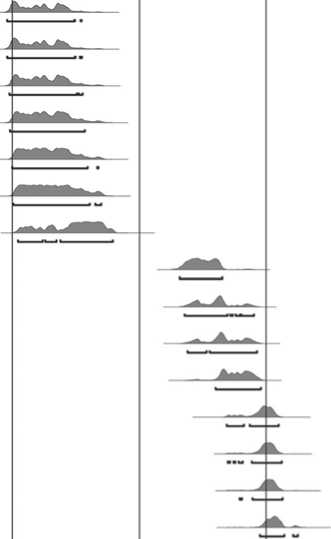
Судя по степени прирастания эпифизов, нерпы в VII слое представлены костными остатками минимум одной особи до одного года и двух половозрелых (молодой и немолодой); в VI слое – двух особей до одного года, одной неполовозрелой и трех немолодых взрослых. Анализ слоев дентина на срезах 10 клыков показал, что они принадлежали двум нерпам до одного года в VII слое; двум особям до одного года, пяти возрастом 1–2 года и одной пяти лет в VI слое. Обобщенные данные по возрастным показателям нерп представлены на рис. 7.
Относительный возраст парнокопытных определялся по состоянию зубов и степени прирастания эпифизов некоторых трубчатых костей. Такие данные полу- чены по четырем косулям [Tomé, Vigne, 2003]. В слое VII представлены остатки двух особей старше и младше 1,3 года, а в VI – животных до 12 месяцев и старше 1,3 года. В VI слое найдены кости благородного оленя, возраст которого между 2 и 4 годами. По наличию рогов представителей семейства оленьих можно предположить, что некоторые костные остатки принадлежали самцам. В VII слое найдены кабаньи клыки взрослого самца (определение пола по: [Hillson, 2005, p. 131]).
Сезонность использования стоянки
Реконструкция сезонов использования стоянки Саган-Заба II по ранним комплексам была осуществлена с помощью упомянутого выше анализа слоев дентина на срезах клыков нерпы, а также по наличию или отсутствию тех видов животных, присутствие которых в Прибайкалье ограничено теплыми месяцами года, как, например, перелетных птиц. Результаты анализа слоев дентина на срезах 10 клыков нерп в целом свидетельствуют о гибели этих животных в весеннее и летнее время (с марта по сентябрь): слой VII – две особи до одного года погибли в период с марта по июнь; слой VI – пятилетняя нерпа была добыта в марте–июне, три особи (1–2 года) – в мае–июне, остальные (до одного года и 1 год) – с июля по сентябрь (рис. 7). Таким образом, эти животные добывались в два сезона: весной, когда они выходят на лед и находятся возле лунок и отдушин, образуя скопления на тающем льду, и летом во время открытого Байкала, когда нерпы распространены по всему озеру и также выходят группами на берег [Пастухов, 1993].
Присутствие костей перелетных птиц тоже предполагает использование стоянки в теплое время года. Например, утки и бакланы, чьи о статки найдены в VII слое, и орланы, кости которых обнаружены в обоих исследуемых слоях, могли добываться в районе бухты Саган-Заба с конца марта до начала октября во время их прилета в Прибайкалье (рис. 7). Но необходимо
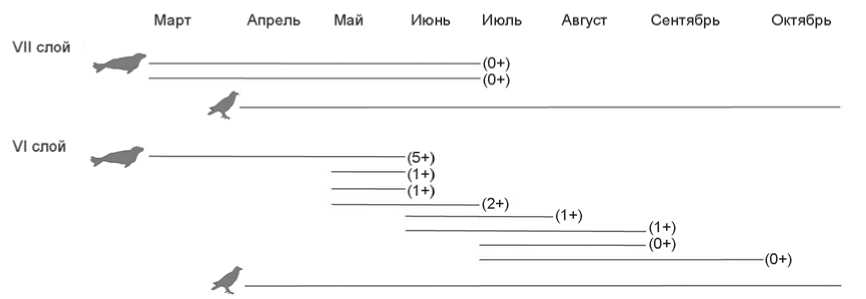
Рис. 7. Сезоны гибели нерп (в скобках указан возраст животного) и птиц на стоянке Саган-Заба II.
иметь в виду, что отдельные особи орланов иногда остаются на зимовку в этом регионе [Птицы Байкала, 2005]. Таким образом, имеющиеся данные по изученным комплексам позволяют предположить использование стоянки Саган-Заба II в основном весной и летом, что не исключает и другое время года.
Заключение
На основе анализа серии радиоуглеродных дат и вычисления показателей их объединенного усреднения стало возможным определить разницу между некалиброванными датами, полученными по костям копытных животных и нерп (для VII слоя – 682 года, для VI – 509 лет). В результате была уточнена датировка ранних комплексов поселения Саган-Заба II. С учетом коррекции дат, полученных по ко стям нерпы, VII слой датируется в интервале 9 120–8 610 кал. л.н., VI – 8 340–7 880 кал. л.н.
Планиграфический анализ показал, что фаунистические о статки в VII и VI слоях располагались преимущественно скоплениями совместно с каменными и ко стяными изделиями, а в VI слое и с керамикой в районах очагов и кострищ. Вероятно, они представляют собой недифференцированные хозяйственнобытовые отбросы. Самостоятельных скоплений фаунистических остатков в слоях не зафиксировано. Концентрация археологических материалов в районе очагов и кострищ характерна для всех комплексов раннего – среднего голоцена не только на побережье Байкала, но и в Прибайкалье в целом.
Вероятно, исследуемые стоянки носили кратковременный, сезонный характер, что подтверждается особенностью размещения очагов и скоплений материалов, незначительной мощностью кострищ, отсутствием специализированных производственных площадок, сравнительно небольшим общим количеством фаунистических остатков. О сезонном их использовании свидетельствуют результаты анализа слоев дентина на срезах клыков нерпы и наличие костей животных, присутствие которых в Прибайкалье ограничено теплыми месяцами года (например, перелетных птиц). Таким образом, можно предположить, что бухта Саган-Заба была обитаемой в период между 9 120 и 7 880 кал. л.н. преимущественно в весеннее и летнее время года.
Основу хозяйственной деятельности в указанный период, судя по видовому составу животных, составляли промысел нерпы и охота на копытных животных. Определенное значение имела и добыча птиц, а также рыболовство. Присутствие в комплексах большого количества битой кости («кухонные отбросы» и отходы производства костяных орудий), ее расположение в основном возле кострищ и очагов, нали- чие костей со следами разделки и жженых позволяют предположить, что добыча, разделка и употребление некоторых животных производились в окрестностях или непосредственно в самой бухте. Следует отметить, что рога и кости парнокопытных использовались для изготовления орудий. Результаты анализа фаунистических материалов из слоев VII и VI многослойного поселения Саган-Заба II свидетельствуют о комплексном использовании природных ресурсов. Необходимо заметить, что на протяжении рассматриваемого промежутка времени (с 9 120 до 7 880 кал. л.н.) в исследованных комплексах не наблюдается какого-либо изменения в добывании животных того или иного вида.
Список литературы Кости животных из ранних комплексов многослойного поселения Саган-Заба II (9 120-7 880 кал. л.н.) в Прибайкалье: планиграфия, хозяйственная деятельность и сезонность использования стоянки
- Асеев И. В. Юго-Восточная Сибирь в эпоху камня и металла. - Новосибирск: Изд-во ИАЭТ СО РАН, 2003. - 208 с.
- Воробьева Г. А. Почва как летопись природных событий Прибайкалья: проблемы эволюции и классификации почв. -Иркутск: Изд-во Иркут. гос. ун-та, 2010. - 205 с.
- Горюнова О.И., Долганов В.А., Новиков А.Г., Вебер А.В. Ранний неолит Приольхонья: по материалам VI культурных слоев геоархеологического объекта Саган-Заба II // Феномен геоархеологической многослойности Байкальской Сибири: 100 лет Байкальской научной археологии. - Иркутск: Изд-во Иркут, гос. ун-та, 2012. - С. 86-93. -(Евразия в кайнозое: Стратиграфия, палеоэкология, культуры; вып. 1).
- Горюнова О.И., Новиков А.Г., Вебер А.В., Воробьева Г. А., Орлова Л.А. Завершение раскопок Российско-Канадской экспедиции в бухте Саган-Заба на Байкале // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. - Новосибирск: Изд-во ИАЭТ СО РАН, 2008. - Т. 14. - С. 32-35.
- Горюнова О.И., Новиков А.Г., Воробьева Г.А., Вебер А.В., Лозей Р.Дж., Номоконова Т.Ю., Орлова Л.А. Продолжение работ Российско-Канадской экспедиции в бухте Саган-Заба на Байкале // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. - Новосибирск: Изд-во ИАЭТ СО РАН, 2007. - Т. 13. -С. 212-215.
