Морфологические особенности почек кур в постнатальном онтогенезе

Автор: Гречко В.В., Овчинников Д.К.
Журнал: Молочнохозяйственный вестник @vestnik-molochnoe
Рубрика: Сельскохозяйственные и ветеринарные науки
Статья в выпуске: 4 (52), 2023 года.
Бесплатный доступ
Статья посвящена исследованию морфологических особенностей мочевыделительной системы у кур кросса «Сибиряк-2» в постнатальном онтогенезе с суточного до девяностосуточного возраста. В ходе исследования были использованы классические гистологические методики. Полученные результаты позволяют уточнить и дополнить информацию по особенностям морфологического строения, диагностике при вирусных заболеваниях, нарушениях содержания и кормления птицы, изменениях органов мочевыделительной системы.
Птицы, курообразные, почки, морфология, гистология, постнатальный онтогенез
Короткий адрес: https://sciup.org/149144593
IDR: 149144593 | УДК: 619:611.61:636.5 | DOI: 10.52231/2225-4269_2023_4_23
Morphological traits of chicken kidneys in postnatal ontogenesis
This scientific article presents a study of the morphological traits of the urinary system in chickens of the cross «Sibiryak-2» in postnatal ontogenesis at the age from one day to ninety days. In the course of the study, classical histological techniques have been used. The results obtained make it possible to describe the morphological structure more accurately and fully, as well as to use it in the diagnosis of diseases, violations of poultry maintenance and feeding, changes in the organs of the urinary system.
Текст научной статьи Морфологические особенности почек кур в постнатальном онтогенезе
Птицеводческая отрасль является одной из ведущих производств сельского хозяйства, так как обеспечивает высококачественными натуральными продуктами питания и сырьем [4–7].
Имеющиеся работы отечественных и зарубежных морфологов посвящены изучению строения отдельных органов и систем птиц, а исследования по морфологии органов выделительной системы у птиц в постнатальном онтогенезе, по мнению многих авторов учебных руководств и оригинальных исследований, по своему содержанию имеют фрагментарный характер [1, 2, 3, 8, 9, 10].
Цель исследования: изучение особенностей строения почек у кур в постнатальном онтогенезе.
Материалы и методы
Объектами для проведения комплексного микроморфологического исследования служили птицы кросса «Сибиряк-2» в постнатальном онтогенезе 1, 15, 30, 60 и 90 дней.
Для гистологических исследований брали кусочки почек размером 10×10 мм, фиксировали в 4% растворе нейтрального формальдегида. Уплотнение материала проводили путем заливки в парафин. С помощью ротационного микротома МБС-2 получали срезы толщиной 5–7 мкм. Срезы окрашивали гематоксилин-эозином, по Ван-Гизону, по Маллори [11–14]. Полученные гистологические препараты изучали с помощью светового микроскопа МИКМЕД-5, окуляр × 10, 15; объектив × 10, × 20, × 40, × 90. Работа выполнена на кафедре диагностики, внутренних незаразных болезней, фармакологии, хирургии и акушерства факультета ветеринарной медицины.
Результаты исследования
Капсула почки покрыта соединительнотканной фиброзной капсулой, покрывающий почку снаружи. В суточном возрасте капсула плотно покрывает паренхиму почки, в ней четко различимы базофильные клетки овальной формы – фиброциты. Фиброциты в капсуле почки суточных цыплят располагаются, как правило, в несколько рядов по 4-5 клеток. В местах, где капсула становится тоньше, данные ряды представлены 2-3 клетками. Ядра фибробластов тесно связаны с окружающей соединительной тканью, что не позволяет различить их цитоплазму. Из-за большого количества фиброцитов капсула имеет более темное окрашивание. Соединительнотканные волокна капсулы почки имеют не только прямое, но и волнообразное направление. Если соединительные волокна направлены волнообразно, то и фиброциты повторяют их форму, и данная структура состоит из 2-3 соединительнотканных тяжей. Наружная капсула практически не сдержит фиброцитов (р ис. 1).
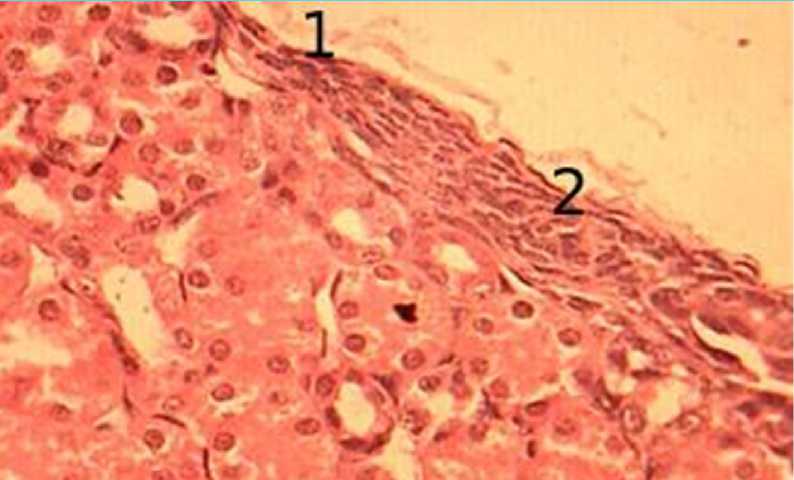
Рисунок 1 - Фиброзная капсула почки суточного цыпленка, окраска гематоксилин эозин, увеличение ×400: 1 – фиброзная капсула, 2 – фиброциты
В 15-суточном возрасте по сравнению с суточным возрастом соединительнотканная фиброзная капсула имеет более светлую окраску и скудное содержание фиброцитов. Фиброциты слабо дифференцируются и представлены в виде тонких базофильных «полосок». Волокна идут волнообразно, и связь с паренхимой почки слабее, чем в суточном возрасте. При окраске гистологических препаратовпоВан-Гизонуколлагеновыеволокнакапсулыокрашиваются в яркий рубиново-красный цвет, а волокна субфиброзной оболочки окрашиваются в более бледный желтый цвет. Данная субфиброзная оболочка дифференцируется на препаратах окрашенных только по Ван-Гизону. Также выявляются волокнистые структуры фиброзной капсулы окраской по Маллори (рис. 2).
В 30-суточном возрасте капсула имеет аналогичное строение, светло розовую окраску, волнообразный ход соединительнотканных волокон и скудное содержание фиброцитов, как и в 15-суточном возрасте.
В процессе онтогенеза к 60-суточному возрасту капсула становится тоньше, а связь с паренхимой органа усиливается. Капсула плотно прилегает, фиброциты имеют как овальную форму с интенсивной базофильной краской, так и округлую.
В 90-суточном возрасте фиброзная капсула слабо дифференцируется, практически не встречаются ядра фиброцитов и они единичные. По сравнению с суточным возрастом, где они располагались в несколько рядов, капсула представлена всего одним слоем соединительнотканных волокон. Коллагеновые волокна тонкие, преобладает волокнистая соединительная ткань, окрашенная по Маллори в темно синий цвет.
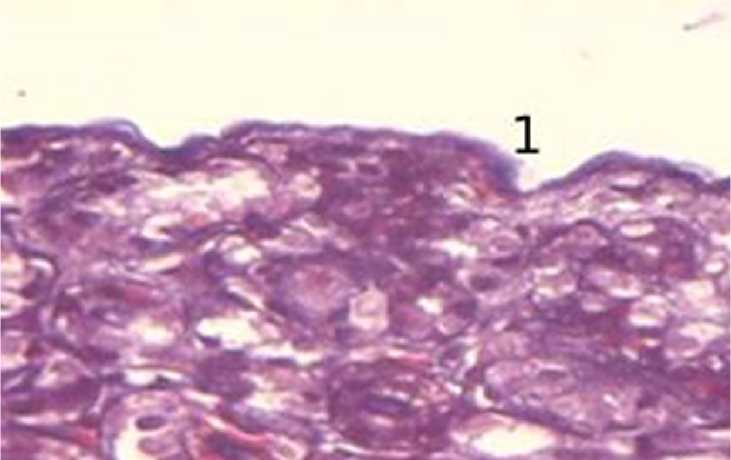
Рисунок 2 - Фиброзная капсула почки в 15-суточном возрасте, окраска по Маллори, увеличение ×400: 1 – рыхлая соединительная ткань
Обобщая все вышесказанное, мы можем сделать вывод: онтогенез соединительнотканной капсулы происходит следующим образом. В суточном возрасте капсула толстая, насыщенная окраска преимущественно базофильная, состоит из нескольких рядов фиброцитов, соединительные волокна в основном коллагеновые. По мере взросления капсула почки истончается, имеет светло-розовую окраску, фиброциты единичные, а волокна представлены преимущественно рыхлой соединительной тканью, коллагеновых волокон становится меньше.
В первые сутки у кур ядра клеток кубического эпителия извитых почечных канальцев имеют округлую форму с небольшими включениями нуклеопротеида. Проксимальные почечные канальцы покрыты плотно лежащими тонкими цитоплазматическими выростами. В их строении прослеживается нечеткая граница между клетками эпителия. Базальная мембрана слабо окрашена. Между проксимальными почечными канальцами наблюдаются дистальные почечные канальцы с меньшим диаметром, но с тем же просветом. Дистальные почечные канальцы выстланы плоским эпителием, а их ядра сжаты в тангенциальном направлении.
В возрасте 15 суток структура почечных канальцев не подвержена значительным изменениям (рис. 3).
По достижении возраста 30 суток происходит увеличение диаметра проксимальных извитых почечных канальцев. В нефроцитах не отмечается наличие выраженных микроворсинок на базальной мембране. Эпителиальные клетки представлены в форме высокопризматического эпителия со смещенными ядрами к нижнему полюсу клетки. В проксимальных и дистальных канальцах ядра имеют правильную форму.
В возрасте 60 суток отмечается более плотное расположение клеток друг к другу. Диаметр центральной части проксимальных извитых почечных канальцев увеличивается.
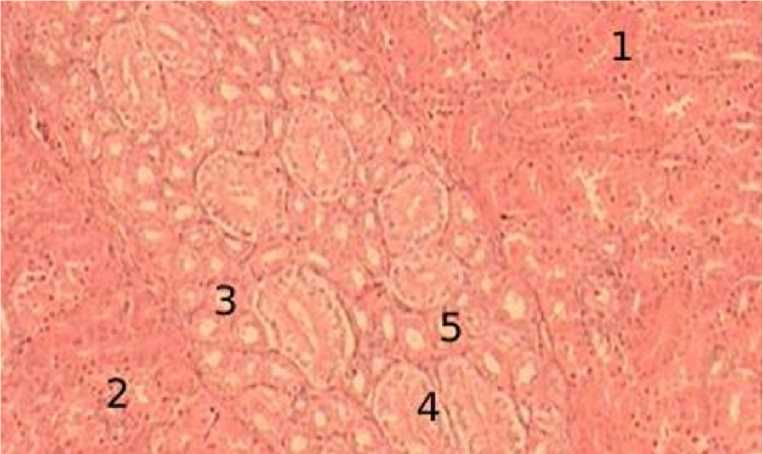
Рисунок 3 - Почечные канальцы, 15-суточный возраст, окраска гематоксилин-эозин, увеличение х400: 1 – проксимальные канальцы, 2 – дистальные канальцы, 3 – прямые участки петель Генле, 4 – собирательные трубочки, 5 – промежуточные канальцы
Количество проксимальных извитых почечных канальцев в возрасте 90 суток увеличивается за счет уменьшения их в объёме. Эпителий почечных канальцев плотный и сформированный. Ядра клеток имеют центральное расположение, границы клеток четко различимы. Базальная мембрана четкая. Плоский эпителий дистальных извитых почечных канальцев имеет равную высоту с кубическим эпителием. Отмечается больший внутренний просвет почечных канальцев. По сравнению с предыдущим возрастом происходит значительное увеличение количества дистальных извитых почечных канальцев и капилляров (рис. 4).
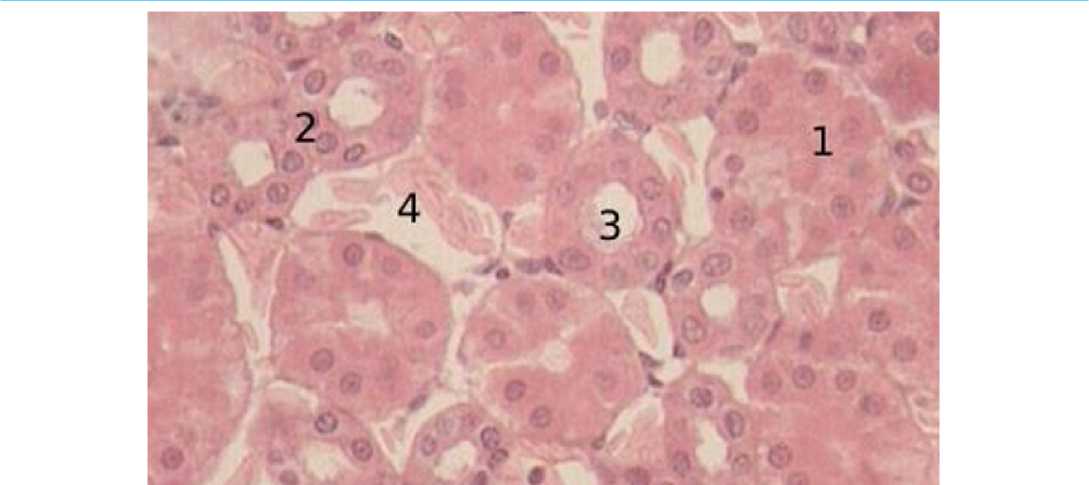
Рисунок 4 - Почечные канальцы, 90-суточный возраст, окраска гематоксилин-эозин увеличение х400: 1 – проксимальный извитой каналец; 2 – дистальный извитой каналец; 3 – просветы канальцев; 4 – капилляры
Собирательные трубочки, прямые участки петель Генле и промежуточные канальцы располагаются в средней части паренхимы почки. В противоположность корковым собирательным трубочкам мозговые собирательные трубочки выстланы однослойным цилиндрическим эпителием, состоящим только из основного типа клеток. Цилиндрический эпителий отличается светлой цитоплазмой, четкими межклеточными границами и округлыми ядрами. Корковые собирательные трубочки выстланы кубическим эпителием, состоящим из основных и вставочных клеток с характерной темной цитоплазмой.
В процессе онтогенеза происходит рост и развитие собирательных трубочек и прямых участков петель Генле. Клетки собирательных трубочек увеличивают свою высоту, апикальная часть прокрашивается лучше по сравнению с базальной частью клетки, а ядра клеток имеют не только центральное расположение, но и сдвинуты к базальной мембране, образуя вокруг себя пространство, не воспринимающее краситель (рис. 5).
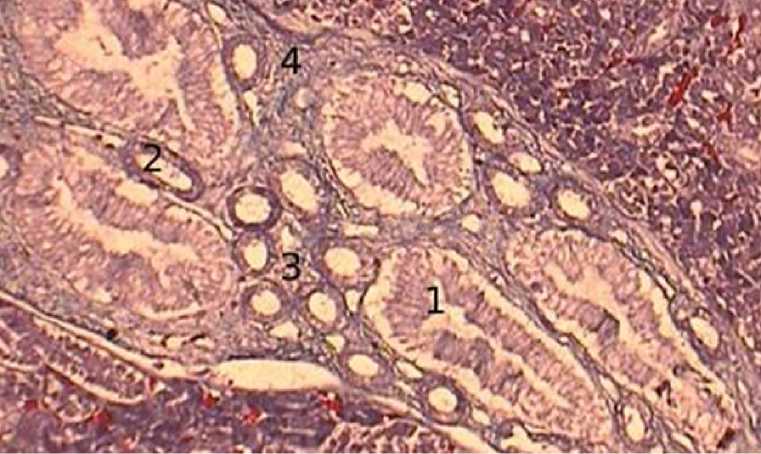
Рисунок 5 - Почечные канальцы, 60-суточный возраст, окраска Маллори, увеличение х400: 1 – собирательные трубочки, 2 – прямые участки петель Генле, 3 – промежуточные канальцы, 4 – волокнистая соединительная ткань
В суточном возрасте суперфициальные почечные тельца располагаются по одному или попарно, на значительном расстоянии, с одинаковым диаметром. Ядра извитых капилляров плотно прилегают (рис. 6).
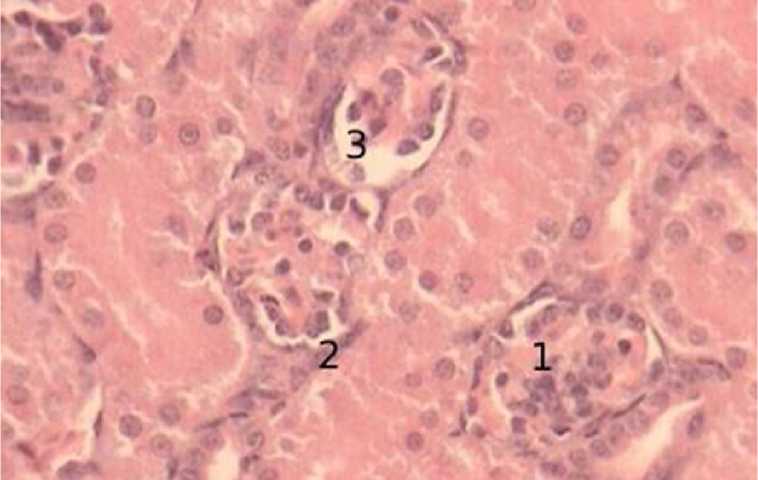
Рисунок 6 - Суперфицальные почечные тельца суточный возраст, окраска гематоксилин-эозин, увеличение х 400: 1 – клубочек, извитые капилляры, 2 – боуменова капсула, наружный листок, 3 – просвет капсулы
В возрасте 15 суток суперфициальные почечные тельца становятся крупнее, существенно изменяется просвет почечной капсулы Боумена. Сама капсула клубочка имеет разные размеры. Гломерула плотная с большим диаметром. В просвете наблюдаются ядра эритроцитов. Ядерные структуры увеличиваются, приобретая светлую окраску.
В возрасте 30 суток суперфициальные почечные тельца увеличиваются. Ядра извитых капилляров располагаются по периферии сосудистых клубочков, имея светлую красно-розовую окраску.
Ввозрасте 60суток, суперфицальные почечные тельца продолжают, увеличиваться в размерах. Ядра извитых капилляров располагаются по периферии сосудистого клубочка и не окрашиваются. Капсула не имеет просвета. Сосудистый клубочек плотно прилегает ко всему висциральному листку боуменовской капсулы.
Суперфициальные почечные тельца в возрасте 90 суток по сравнению с предшествующим возрастом не отличаются.
Интракортикальные почечные тельца располагаются в средней зоне мозгового вещества. В суточном возрасте они располагаются группами, в ряды, в количестве 3-4 почечных телец в одном поле зрения, также мы отмечаем их парное расположение. При такой локализации боуменовой капсулы и сосудистых клубочков они имеют одно место соприкосновения. Почечные тельца не имеют правильной округлой формы и их диаметр не одинаковый.
Толщина стенок боуменовой капсулы и ее просвет различный. Чем меньше извитые капилляры сосудистого клубочка, тем тоньше внутренний и наружный листок капсулы, а также больше ее просвет. Извитые капилляры соприкасаются с капсулой в нескольких местах. Сами почечные тельца имеют темную окраску ядер.
Ядра локализуются по периферии извитых капилляров сосудистых клубочков. Интракортикальные нефроны, в которых эритроциты не встречаются, имеют ядра с более светлой и прозрачной окраской, располагаются они плотной группой от базальной мембраны капсулы до центра сосудистого клубочка.
В 15-суточном возрасте количество и размер интракортикальных почечных телец остается практически неизменным. Капсула почки имеет больший просвет почечных телец в отличие от суточного возраста. Мы отмечаем плотное расположение сосудистых клубочков к внутреннему листку боуменовой капсулы. Широкий просвет капсулы обращен у всех клубочков в одну сторону. Сосудистые клубочки в данный возрастной период не содержат эритроцитов. Извитые капилляры представлены большим количеством тесно расположенных ядер, имеющих светлую окраску и мелко распыленный хроматин, распределенный по всей поверхности ядер. Ядра, локализующиеся на периферии сосудистого клубочка, обращенного к просвету капсулы идут по ходу ветвления капилляра (рис. 7).
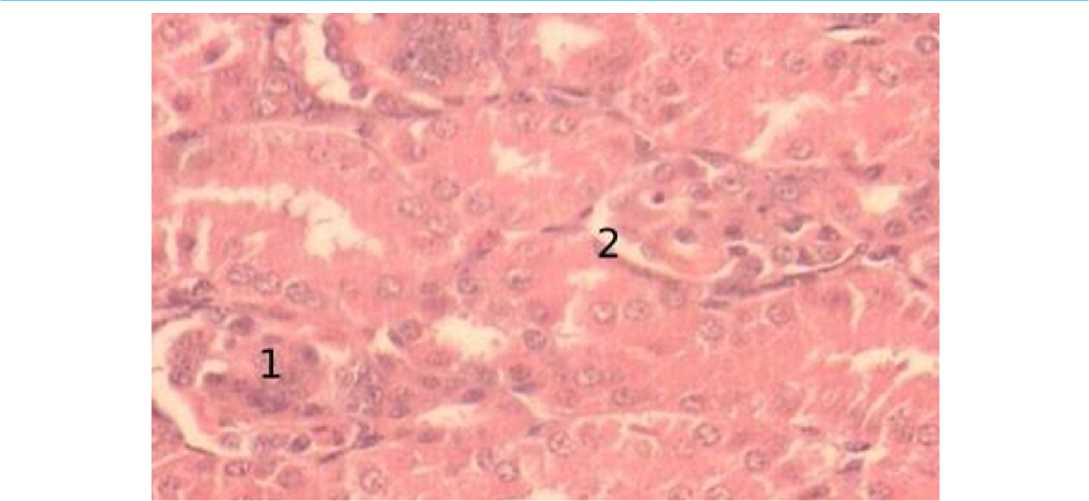
Рисунок 7 - Интракортикальные почечные тельца, 15 суточный возраст, окраска гематоксилин-эозин, увеличение х400: 1 – клубочек, извитые капилляры; 2 – боуменова капсула, наружный листок
В последующем периоде онтогенеза, а именно в 30-суточном возрасте мы отмечаем, резкое увеличение размера интракортикальных почечных телец. Боуменова капсула увеличивает свой диаметр, а толщина ее стенок становится больше. Сосудистые клубочки, как правило, не имеют тесной связи с капсулой, встречается только однократное место соприкосновения клубочка и подоцитов. Структура извитых капилляров становится более сложной, отмечаются не только плотно расположенные скопления ядер, но и сложное расположение ядер по ходу ветвления капилляров. Сами же интракортикальные почечные тельца локализуются в одну цепочку, друг за другом, примерно на равном расстоянии – радиально (рис. 8).
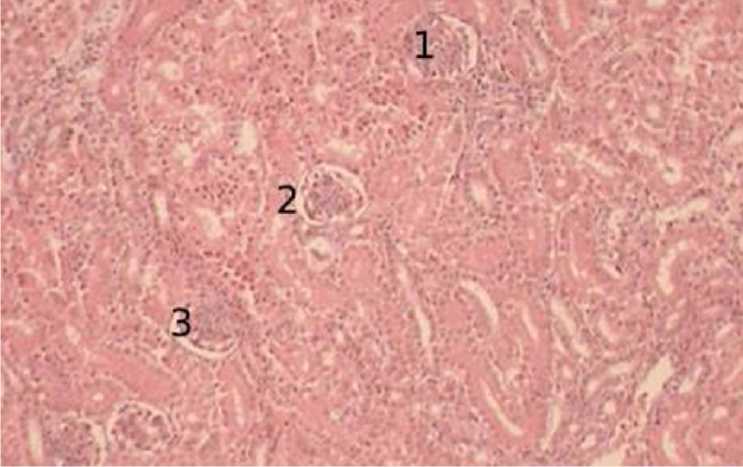
Рисунок 8 - Интракортикальные почечные тельца, возраст 30 суток, окраска гематоксилин-эозин, увеличение х100: 1 – почечные тельца, 2 – боуменова капсула, 3 – просвет капсулы
В 60-суточном возрасте интракортикальные почечные тельца располагаются попарно и имеют значительный диаметр в отличие от 30-суточного возраста. Извитые капилляры плотно прилегаю к капсуле почки, не имея просвета. Ядерные структуры капилляров сосудистых клубочков прозрачные, обладают светлой окраской, пылевидный хроматин окрашивается ярче фона самого ядра. Количество ядер меньше и характер их расположения менее плотный по сравнению со строением интракортикальных почечных телец в более ранних возрастах. И сами капилляры имеют слаборозовую окраску, сливаясь с дистальными и проксимальными извитыми канальцами.
В 90-суточном возрасте мы отмечаем схожее строение интракортикальных телец с 60-суточным возрастом. Тельца располагаются радиально на равном удалении друг от друга с хорошо развитой капсулой. Просвет капсулы меньше, чем в 60-суточном возрасте. Извитые капилляры представлены плотнорасположенными ядрами со светло-базофильной окраской. Коллагеновые волокна и рыхлая соединительная ткань отсутствует в интракортикальных почечных тельцах в течение всего онтогенеза.
Юкстамедуллярные почечные тельца локализуются в пограничной зоне между корковым и мозговым веществом. Почечные тельца у птиц в суточном возрасте в пограничной зоне располагаются, как правило, по одному. Размеры их меньше, чем у суперфициальных и интракортикальных почечных телец. Сосудистый клубочек плотно прилегает к внутреннему листку боуменовой капсулы, из-за чего просвет слабо выражен. Стенки капсулы имеют одинаковую толщину.
Ядра извитых капилляров представлены в виде скудных скоплений в центре клубочка и имеют темную окраску. Вблизи данных скоплений встречаются крупные, округлые ядра с прозрачной кариоплазмой. Сосудистые клубочки, содержащие эритроциты практически не встречаются.
В 15-суточном возрасте отмечается существенное увеличение количества юкстамедуллярных почечных телец, располагающихся как по одному, так и попарно. Также встречается и по три почечных тельца в непосредственной близости. Как и в суточном возрасте, слабо выражен просвет капсулы. Стенки капсулы имеют одинаковую толщину. Ядра извитых капилляров локализуются в центре клубочков, но их количество значительно увеличивается. Ядра крупные округлые со светлоокрашенной кариоплазмой. Ядра, имеющие плотную окраску с неразличимым хроматином оттеснены к периферии клубочков. Сосудистые клубочки, содержащие эритроциты, встречаются чаще (рис. 9).
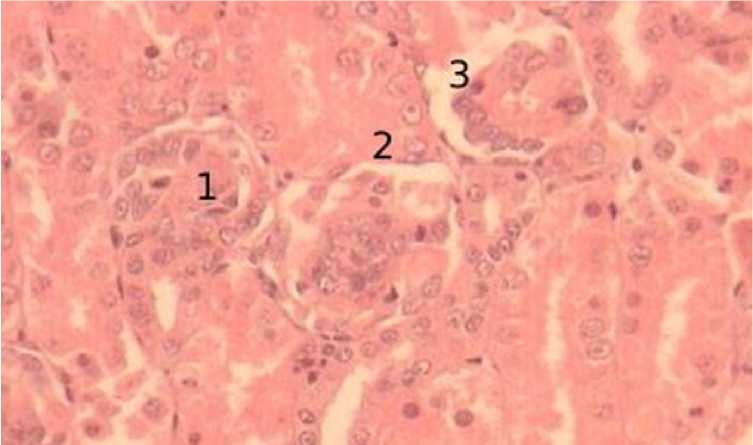
Рисунок 9 - Юкстамедуллярные почечные тельца, 15-суточный возраст, окраска гематоксилин-эозин, увеличение х100: 1 – почечные тельца, 2 – боуменова капсула, 3 – просвет капсулы
К 30-суточному возрасту количество юкстамедуллярных почечных телец продолжает увеличиваться, кроме того увеличивается диаметр непосредственно самих сосудистых клубочков и капсулы, покрывающей их. В отличие от суточного и 15-суточного возраста, когда почечные тельца имели примерно равный диаметр, в 30-суточном возрасте диаметр юкстамедуллярных почечных телец сильно варьируется. Расположение почечных телец хаотичное на различном расстоянии друг от друга. Форма округлая, извитые капилляры прилегают к подоцитам только с одной стороны почечного тельца. Стенки капсулы имеют разную толщину, и на данном этапе онтогенеза отмечается наличие ядер непосредственно в самой боуменовой капсуле. Эти ядра имеют овальную и s-образнную форму, слабо базофильную окраску и практически прозрачны.
Ядра извитых почечных капилляров собираются в группы на одинаковом расстоянии между собой с выраженной темной окраской. В почечных тельцах сосудистых клубочков, встречаются эритроциты. Почечные тельца имеющие меньший диаметр и представлены хорошо выраженным просветом капсулы, ядра капсулы в виде тонкой полоски, толщина стенок капсулы одинакова. Ядра извитых капилляров находятся в центре, на близком расстоянии и существенно в меньшем количестве.
В 60-суточном возрасте количество юкстамедуллярных почечных телец меньше 30-суточного возраста, они локализуются группами в количестве 3–5, иногда до 7 почечных телец. Полость боуменовой капсулы менее выражена из-за плотно прилегающих извитых канальцев на всей площади внутреннего листка капсулы. Отличительной особенностью в данный период онтогенеза является более выраженная локализация ядер извитых канальцев по периферии клубочков. В извитых канальцах эритроциты не отмечаются.
К 90-суточному возрасту юкстамедуллярные почечные тельца становятся крупнее, располагаются попарно. Листки боуменовой капсулы соприкасаются друг с другом, плотно прилегая к окружающим дистальным и проксимальным отделам нефрона. Сосудистый клубочек тесно связан с подоцитами, в результате чего полость капсулы слабо выражена. Мелкие ядра извитых капилляров в большом количестве распределены по всей поверхности сосудистого клубочка, более крупные и прозрачные ядра оттеснены к периферии так же в виде небольших групп (рис. 10).
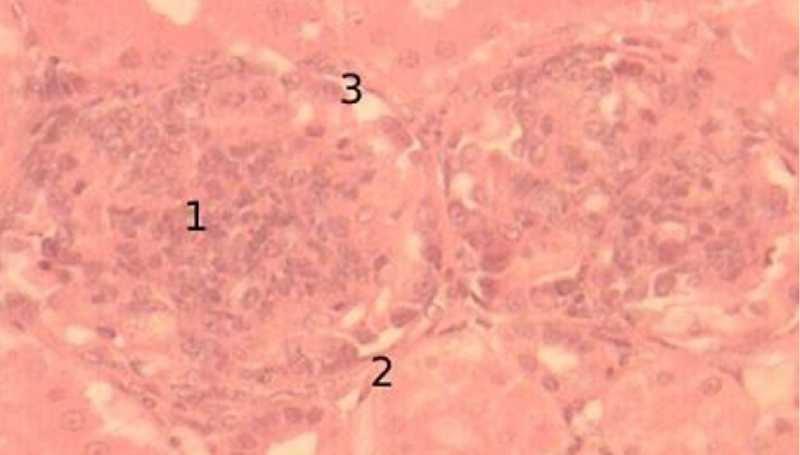
Рисунок 10 - Юкстамедуллярные почечные тельца, 90-суточный возраст, окраска гематоксилин-эозин, увеличение х100: 1 – почечные тельца, 2 – боуменова капсула, 3 – просвет капсулы
Заключение
Обобщая материалы собственных исследований и высказывания ряда авторов [4], мы соглашаемся, что птицеводство одно из перспективных разделов животноводства, и в закономерностях строения органов мочевыделения у птиц в постнатальном онтогенезе имеются возрастные особенности [1, 6].
Анализируя материалы собственных исследований и данные специальной литературы [7], мы можем утверждать, что соединительнотканные волокна и ядра располагаются в скудном количестве во всех структурах почки. Проксимальные и дистальные почечные канальцы в процессе развития не только увеличиваются в размере, но и возрастает их количество. В работах [5, 8] площадь извитых почечных канальцев у кур увеличивается за счет уменьшения площади просвета между капсулой.
С возрастом просвет почечных канальцев, количество эпителиальных клеток, расположение ядер в них и даже сам характер окрашивания изменяются. По данным Л.Н. Воронова [9], у перепела японского площадь просвета и высота эпителиоцитов с возрастом увеличиваются в разы, то же мы наблюдали и у курицы.
Фиброзная капсула на 90% состоит из плотных коллагеновых волокон,оставшиесяволокнапредставленыволокнистойсоединительной тканью.
Изменение в строении суперфициальных почечных клубочков в процессе онтогенеза связано с увеличением функции почки по сравнению с таковой в однодневном возрасте для поддержания гомеостаза.
Гистоструктура и архитектоника паренхимы почки отличаются на всех этапах постнатального онтогенеза. Капсула почки в суточном возрасте состоит из фиброцитов, послойно лежащих друг на друге в 4-5 рядов, создавая крупную стенку капсулы. Ядра фиброцитов со светлой кариоплазмой хорошо различимы и имеют вытянутую форму. С возрастом капсула истончается. Проксимальные и дистальные почечные канальцы в процессе онтогенеза увеличиваются в количестве, размере и просвете.
Полученные данные являются фундаментальной основой для дальнейших научных исследований в области нефрологии (биохимические показатели крови, общий анализ мочи – разработка диагностического профиля, выделительная цистография, рентгенография) в функциональном и прикладном аспекте у птиц в промышленном и домашнем содержании.
Список литературы Морфологические особенности почек кур в постнатальном онтогенезе
- Al-Azawy N.H. Comparative Anatomical and Histological Study of Kidney in Domestic Fowls and Geese. M.Sc. Diss. Iraq, 2005, pp. 45-59. (In English)
- Baumel J.J. et al. Handbook of Avian Anatomy. Cambrige, Massachusetts, Club Publ., 1993, pp. 330-347. (In English)
- Brenner B.M. Kidney development. The Kidney. New York, 1996, Vol.1, pp. 632-695. (In English)
- Fisinin V.I. Strategic trends of innovative development of poultry farming. Bulletin of the Russian Academy of Agricultural Sciences, 2015, no. 1, pp.11-14. (In English)
- Leshchinsky S.V. Morphology of Kidneys of Chickens of the Breed «Lohmann-Brown» in Postnatal Ontogenesis in Normal and when Using Mineral Top Dressing «BS»: Abstract of Dissertation of the Candidate of Veterinary Sciences: 16.00.02. Cand. Diss. Omsk, 2003. 20 p. (In English)
- Nicholson J. K. The microanatomy of the distal tubules, collecting tubules and collecting ducts of the starling kidney. J. Anat., 1982, no. 134(1), pp. 11-23. (In English)
- Techver Yu.T. Kidneys. Histology of Domestic Birds. Tartu, 1965. 76 p. (In English)
- Vodyanitskaya T.S. Features of Macro- and Microanatomy of the Kidneys of Haysex Brown Cross Chickens in Postnatal Ontogenesis: Abstract Diss.... Candidate of Veterinary Sciences: 16.00.02. Cand. Diss. Bryansk, 2006. 24 p. (In English)
- Voronov L.N. Kidney growth in chicks of rooks and jackdaws in the postnatal period. Ecology, 1983, No. 2, pp. 83-84. (In English)
- Ramzi Abdul-Gahaffor Abood AL-Ajeely, Fadhil S. Mohammed. Morpho-histological study on the development of kidney and ureter in hatching and adulthood racing pigeon. I.J.S.N., 2012, vol. 3(3), pp. 665-667. (In English)
- Лилли Р. Патогистологическая техника и практическая гистохимия: пер. с англ. / Р. Лилли; под ред. и с предисл. чл.-кор. АМН В.В. Португалова. – Москва: Мир, 1969. – 645 с.
- Маллори Ф.Б. Патологическая техника / Ф.Б. Маллори. – Нью- Йорк, 1961. – 470 с.: ил.
- Меркулов Г.А. Курс патологогистологической техники / Г.А. Меркулов. – 5-е изд., испр. и доп. – Ленинград: Медицина. Ленингр. отд-ние, 1969. – 423 с.: ил.
- Ромейс Б. Микроскопическая техника / Б. Ромейс; пер. с нем. проф. В.Я. Александрова, З.И. Крюкова; под ред. и с предисл. проф. И.И. Соколова. – Москва: Изд-во иностр. лит., 1954. – 719 с.: ил.
