Новые данные о миграции береговой линии Белого моря

Автор: Шелехова Татьяна Станиславовна, Лаврова Надежда Борисовна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Науки о земле
Статья в выпуске: 2 (115), 2011 года.
Бесплатный доступ
Диатомовый анализ, палинологический анализ, спорово-пыльцевой комплекс, белое море
Короткий адрес: https://sciup.org/14749859
IDR: 14749859
Текст статьи Новые данные о миграции береговой линии Белого моря
Исследованная территория расположена в низовье р. Выг, где находятся древние памятники наскальных искусств Залавруги (рис. 1). Изучению колебаний уровня Белого моря в поздне- и после-ледниковье, а также палеогеографическим реконструкциям голоцена Юго-Западного Беломорья, особенно в связи с древними поселениями человека, посвящены многочисленные публикации [1], [8], [9], [12], [15], [16], [19] и др. Среди них отметим работу Э. И. Девятовой, в которой на основании геолого-геоморфологических условий в низовье р. Выг, результатов спорово-пыльцевого и радиоуглеродного анализа представлены новые данные о трансгрессивно-регрессивных стадиях в развитии Белого моря, прослежена динамика в развитии древних поселений [3]. В этой работе Э. И. Девятова отмечала неправомерность определения хронологической последовательности стоянок по их высотному положению относительно уровня водоема без учета всех остальных особенностей формирования поверхности. По ее мнению, уточнению палеогеографической обстановки могут способствовать палеонтологические анализы торфяных отложений, расположенных на всех ступенях террас [3]. Нами были исследованы торфяные отложения болота, находящегося на абсолютной отметке 16,8 м, при этом использовался сопряженный комплекс методов (диатомовый, палинологический, радиоуглеродный), позволивший получить достаточно полную и достоверную информацию о динамике и хронологии климата и растительности, колебаниях уровня Белого моря, изменении его береговой линии, предопределивших заселение территории и существование поселений.
МАТЕРИАЛЫ И МЕТОДИКА
В 500 м к западу от Старой Залавруги (N 64°29'50", E 34°40'6,5") пробурена скважина мощностью 50 см, на забое которой тонкозернистые пески тонким слоем прикрывают кристаллические породы. В разрезе выделены 5 горизонтов, из которых послойно отобраны 7 образцов на спорово-пыльцевой и диатомовый анализы. С глубины 40–45 см взят образец на радиоуглеродный анализ. Разрез представлен следующими отложениями (сверху вниз): 0–20 см – торф, 20– 36 см – торфянистый сапропель, 36–39 см – слой органики (вероятно, с углистыми частицами), 39–45 см – сапропель, 45–50 см – песок тонкозернистый (рис. 2, 3).
Спорово-пыльцевой анализ выполнен по 7 образцам, отобранным через 5–10 см. При подготовке образцов применялась общепринятая методика [17]; при определении пыльцы и спор использовались справочники-определители [10], [11] и др., а также эталонная коллекция пыльцы и спор Института геологии КарНЦ РАН. Обработка материала, построение диаграммы процентного содержания пыльцы и спор проводились при помощи программ TILIA, TILIA-GRAPH [25] и TG-View 2.0.2. На основании изменений в составе спорово-пыльцевых спектров выделены палинологические зоны (ПЗ), для более объективного зонирования диаграммы использовалась программа CONISS. Периодизация диаграммы и установление относительного возраста отложений на основании палинологического анализа подтверждаются данными радиоуглеродного датирования.
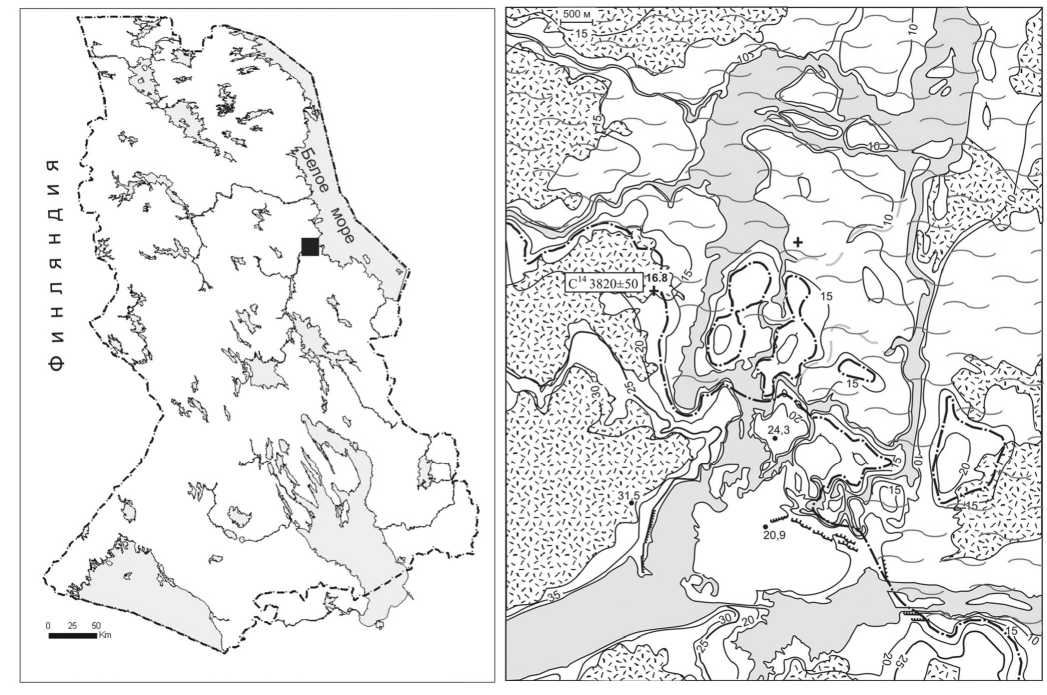

Рис. 1. Месторасположение изученных разрезов и береговая линия моря в суббореальное время: 1 - болота; 2 - береговая линия в суббореале; 3 - расположение разрезов; 4 - радиоуглеродная датировка; 5 - территория, занятая морем в суббореале
Техническая обработка проб и приготовление постоянных препаратов на диатомовый анализ выполнялись по общепринятой методике [2], [4]. При идентификации флоры использовалась справочная литература [26], [28] и др. Всего исследованы 7 образцов послойно без пропусков с разными интервалами. Результаты исследований представлены на диатомовой диаграмме (рис. 2).
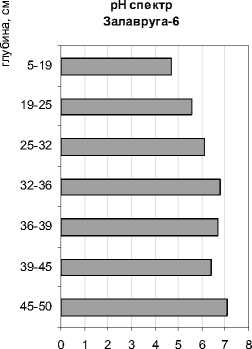
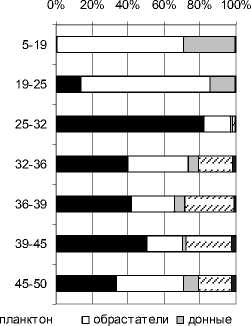
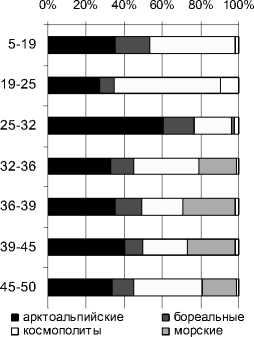
По составу ископаемой диатомовой флоры выполнены реконструкции теоретических значений рН-среды (по [27]). С использованием полученных результатов проводился подсчет видов (в %) по отношению к географическому распространению: арктоальпийских, бореальных и космополитов. Все эколого-географические характеристики приведены на рис. 3.
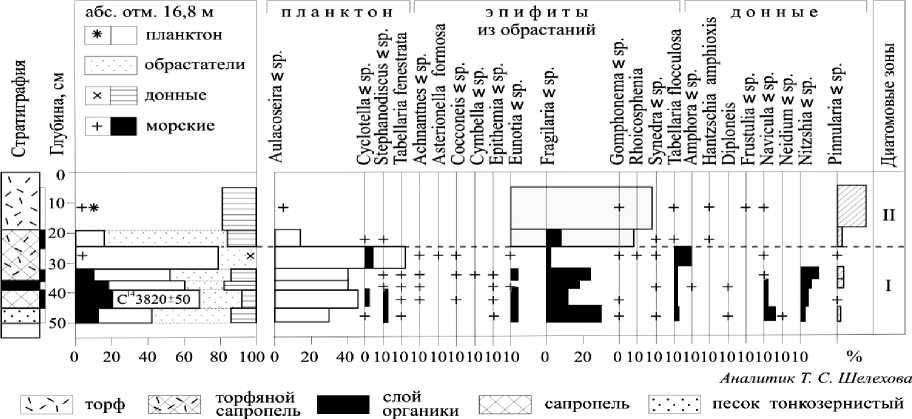
Рис. 2. Диатомовая диаграмма отложений разреза Залавруга-6
рН
Местообитание
Залавруга-6
морские неизв ест ные
Рис. 3. Эколого-географическая характеристика и рН-спектр разреза Залавруга-6
Биоге ография
Залавруга-6
неизвестные
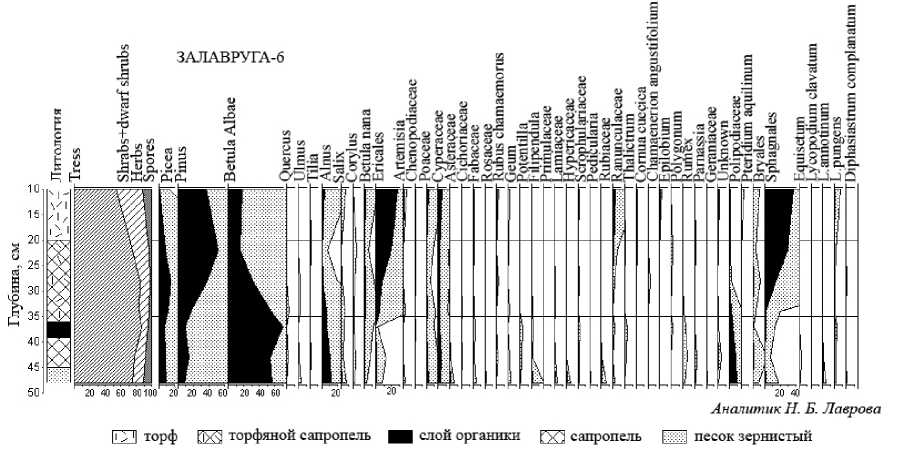
Рис. 4. Спорово-пыльцевая диаграмма разреза Залавруга-6
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Диатомовый анализ
В отложениях обнаружен довольно богатый комплекс пресноводной диатомовой флоры, состоящий из 85 видов и разновидностей пресноводных диатомей (табл. 1), а с глубины от 50 до 32 см встречаются многочисленные обломки морских и солоноватоводных видов, отмеченных ранее [24] в разрезе Залавруга-4. Их доля в общем составе флоры составляет 10–20 %, причем наибольшие значения характерны для глубины 35–45 см. Морские и солоноватоводные виды представлены 24 таксонами, список которых приводится в табл. 2.
По составу диатомовых комплексов можно выделить два этапа развития диатомовой флоры:
первый – глубина 50–32 см, второй – 32–0 см. На первом этапе формирование флоры происходило в условиях притока морских вод, на втором – диатомовый комплекс развивался исключительно в пресноводной среде.
Первый этап – глубина 50–32 см. В нем можно выделить время накопления песков, когда пресноводный комплекс пополнялся морскими формами, и период отложения сапропелей с торфом и органикой, во время которого отмечается максимальное участие морских диатомей. Пресноводная флора представлена планктонными формами родов Aulacoseira, Stephanodiscus, единичными Cyclotella, Tabellatia fenestrata, эпифитами Tabel-laria flocculosa, Fragilaria sp., Eunotia sp. и донными Navicula sp., Nitzschia sp., Pinnularia sp. Их содержание достигает 80 % (рис. 2). Среди них преобладают арктоальпийские виды (30–40 %) и космополиты (20–30 %). Среда близка к нейтральной (рН около 7) (рис. 3). Солоноватоводноморской комплекс состоит из 24 таксонов (табл. 2), в нем доминирует Paralia sulcata + P. sulcata var. crenulata – вид, обитающий в мелководной литоральной зоне моря. Выше по разрезу, когда пески сменяются сапропелями, в солоноватовод-но-морской флоре присутствуют типичные бентосные формы из рода Nitzschia, явно указывая на снижение уровня моря до глубины не более 0,5 м. Все остальные солоноватоводно-морские формы встречаются в данной части разреза единично. Вероятно, накопление песков происходило во время регрессии моря и периодического поступления морских вод в залив в результате приливно-отливной деятельности, а сапропелей с торфом и органикой – в период полного отделения данной территории от моря в виде мелкой замкнутой котловины с остаточной морской водой и постепенного опреснения. Последние результаты исследования переходных зон от морских условий к пресноводным в донных отложениях озер Прибеломорской низменности, расположенных на разных абсолютных отметках, указывают, что наличие около 20 % солоноватоводно-морских видов, как в нашем случае, свидетельствует о регрессивной фазе моря [21], [22], [23]. Отложение торфяного сапропеля (глубиной 32–35 см), по данным радиоуглеродной датировки (С14), происходило 3820 ± 50 лет назад (Ле8360). Максимальное содержание в данном интервале пресноводных из родов Fragilaria sp., Eunotia sp. может свидетельствовать об очень небольшой глубине бассейна и начальных стадиях заболачивания. Следовательно, во время регрессии моря около 3820 ± 50 лет назад (С14) скалы Залавруги с абсолютной отметкой 16,5 м не были покрыты водой и были созданы благоприятные условия для существования поселений.
Второй этап – глубина 32–0 см. Накапливается торфянистый сапропель, резко сменяющийся торфом. Состав флоры полностью пресноводный с единичными случайными обломками створок морских видов, что свидетельствует о прекращении притока морских вод, резком падении уровня воды и начале торфонакопления. Пресноводный диатомовый комплекс представлен типично болотными формами-обрастателя-ми и донными из родов Eunotia (70–80 % флоры) и Pinnularia (16 %), а также единичными Tabel-laria flocculosa , Hantzschia sp ., Frustulia sp ., Na-vicula sp. Нейтральная среда воды резко сменилась на кислую (рН снизилась с 6,9 до 5). Повысилось до 50 % содержание видов-космополитов, что наблюдается при некотором потеплении климата. Примечательно присутствие Hantzschia amphioxis . Обычно этот вид обнаруживается в золе кострищ, на мусорных местах, поэтому можно предполагать близость расположения стоянок человека и возможность пожаров.
Спорово-пыльцевой анализ
Спорово-пыльцевая диаграмма (рис. 4) характеризуется преобладанием пыльцы древесных пород на всем протяжении исследованного слоя, но при этом нельзя не отметить разнообразие травянистых. Помимо пыльцы Poaceae , Cy-peraceae , присутствует пыльца Asteraceae , Ci-choriaceae , Fabaceae , Primulaceae , Lamiaceae , Hypericaceae , Scrophulariaceae , Pedicularis , Ru-biaceae (Galium) , Ranunculaceae , Thalictrum , Cornus suecica , Chamaenerion angustifolium , Epi-lobium sp ., Polygonum sp ., Rumex , Parnassia , Ge-raniaceae , Rosaceae (в последнем встречается пыльца как определенная лишь до уровня семейства, так и принадлежащая следующим видам и родам: Rubus chamaemorus , Geum rivale , Filipendula ulmaria , Potentilla ). В количественном отношении доля этой пыльцы невелика, лишь Filipendula в нижней части диаграммы едва достигает 1 %, а Ranunculus в ее верхней части – 5 %. Отмечены единичные пыльцевые зерна древесных неморальных видов: Quercus , Ulmus , Tilia . Пыльца этих пород, по всей вероятности, синхронна времени накопления осадков, но, принимая во внимание ее ничтожную долю в отложениях, можно предположить, что граница их распространения располагалась на некотором расстоянии от изучаемой территории [14]. Наряду с пыльцой термофильных пород встречена пыльца гипоарктических видов ( Betula nana, Rubus chamaemorus, Lycopodium pungens ), которые, по всей вероятности, были приурочены к болотам и прибрежным местообитаниям. Анализ полученных данных позволяет выделить 3 пыльцевые зоны (ПЗ), отражающие определенные этапы в развитии растительности.
Первая пыльцевая зона (ПЗ 1, глубина 50–36 см) выделена в сапропеле, под которым залегает тонкий слой песка. Песок в незначительных количествах встречается во всем слое сапропеля, темно-коричневый цвет которого вверх по разрезу сменяется черным (36–39 см). Черный цвет, по всей вероятности, указывает на присутствие углистых частиц. Эти отложения, в соответствии с данными радиоуглеродного анализа (ЛЕ 8360, 3800 ± 50 лет назад), формировались во второй трети суббореального периода, климатические параметры этого времени несколько превышали современные [6]. ПЗ 1 характеризуется преобладанием пыльцы древесных пород, главным образом Betula sect. Albae , достигающей своего максимума (60 %) в отложениях черного сапропеля. В пределах 10–15 % варьируют кривые пыльцы Pinus и Picea , почти такие же значения имеет пыльца Alnus . Отмечены единичные пыльцевые зерна термофильных пород ( Quercus , Tilia , Ul-mus ). Постоянно встречается пыльца Corylus . Среди травянистых лидируют Cyperaceae и Poa-ceae , в разнотравье первые позиции занимают Filipendula ulmaria , Rubiaceae (Galium) , Hyper-icaceae , Geum rivale , Ranunculaceae , Thalictrum , среди споровых – Polypodiaceae . Обычны споры
Equisetum , Lycopodium annotinum , L. pungens , Diphasiastrum complanatum. Таким образом, вышеприведенные данные позволяют предположить, что на изучаемой территории преобладали березовые с примесью лещины и ольхи крупнотравные леса. Пыльца термофильных пород Tilia , Quercus , Ulmus , по всей вероятности, была заносной (см. выше). Максимального развития березняки достигли около 3800 лет назад. Принимая во внимание пыльцевую продуктивность ели и сосны, вполне вероятно предположение, что в благоприятных местообитаниях встречались небольшие участки ельников. Участие сосны в древостое, напротив, представляется весьма незначительным. Это обусловлено достаточно высоким уровнем моря, что следует из результатов диатомового анализа. Следовательно, уровень грунтовых вод был довольно высоким. Ольховые сообщества с влаголюбивым разнотравьем приурочены к наиболее переувлажненным, прибрежным участкам, которые постепенно зарастали болотной и прибрежноводной растительностью ( Rubus chamaemorus , Parnassia , Cyperaceae , Equisetum и др.). Такие виды, как Filipendula ulmaria , Geum rivale , тяготеют к сырым берегам, заболоченным лугам. В подобных местообитаниях могут встречаться виды Rumex , Ranunculaceae , Geraniaceae , Poten-tilla ; некоторые виды этих родов и семейств предпочитают морские побережья, расселяясь на лугах так называемого третьего уровня, испытывающих незначительное влияние морской воды [7], [18]. Отметим, что в этих отложениях определена пыльца семейства Asteraceae , принадлежащего типу Aster. Ранее, изучая морские и переходные от морских к озерным отложения, мы сделали предположение, что пыльца этого типа продуцируется астрой солончаковой - облигатным галофитом, расселяющимся на регулярно затапливаемых морской водой местообитаниях [13]. Учитывая данные диатомового анализа, указывающего на присутствие морских диатомей, то есть на формирование отложений в морских условиях, предположим, что и здесь могли произрастать облигатные и факультативные галофиты, а также гликофиты из семейств Ranun-culaceae , Fabaceae , родов Rumex, Potentilla . Обращает на себя внимание и отсутствие пыльцы макрофитов. Скорее всего, в спорово-пыльцевых спектрах нашла отражение локальная растительность зоны контакта между галофитами и растениями незасоленных почв, указывая на близость береговой линии моря. Разнообразие растительности, мягкие климатические условия, близость водоема делали рассматриваемую территорию благоприятной для создания поселений.
Состав спорово-пыльцевых спектров, слагающий вторую пыльцевую зону (ПЗ 2, глубина 34-20 см), претерпевает значительные изменения, а во вмещающих их отложениях - сапропеле - увеличивается примесь растительных остатков, и он постепенно приобретает вид торфя- нистого сапропеля. Отметим, что количество пыльцы Betula Albae уменьшается с 70 до 15 %, Alnus - до 1-5 %, Pinus, напротив, возрастает с 10 до 50 %. Во-вторых, значения Ericales и Sphagnales увеличиваются с 1-20 до 30 % соответственно. В чем причина столь резких изменений? На глубине 0,36-0,39 м залегает слой органики черного цвета, по всей вероятности, с углистыми частицами. Можем ли мы говорить о постпирогенных сукцессиях? Ведь при лесных пожарах обычно в большей степени страдают хвойные породы, береза же как порода-пионер первая начинает занимать гари. В нашем случае картина иная. Возможно, перераспределение процентного соотношения пыльцы в пользу сосны связано с регрессией моря, что подтверждается и данными диатомового анализа (см. выше). Регрессия повлекла за собой понижение уровня грунтовых вод и как следствие - зарастание водоема, на что указывает состав осадков и направленное нарастание кривой пыльцы Sphag-nales. Этот же фактор способствовал широкому распространению кустарничниковых сосняков. Но некоторые верескоцветные кустарнички произрастали также на заболоченных берегах водоема. Интересно отметить, что в составе растительного покрова постепенно возрастает участие ельников, которые тем не менее не занимают ведущего положения, каковое отмечалось в суб-бореале практически для всей территории Карелии [6]. Вероятно, это было обусловлено локальными почвенно-эдафическими и гидрологическими условиями изучаемой территории.
В то же время представляется весьма возможным, что осушение территории могло способствовать возникновению лесных пожаров, о чем свидетельствуют находки пыльцы Chamae-nerion angustifolium , Calluna vulgaris , углистых частиц и диатомовых водорослей Hantzschia am-phioxis , встречающихся в золе кострищ.
Непременно следует отметить также и то, что в спорово-пыльцевых спектрах ПЗ 1 и ПЗ 2 не встречена пыльца гидрофитов. Возможно, это обусловлено слабо повышенной соленостью воды. В подтверждение этому в отложениях, соответствующих ПЗ 1 и ПЗ 2, обнаружены единичные створки морских диатомей. Данные палинологического и радиоуглеродного анализа позволяют предположить, что спорово-пыльцевые спектры ПЗ 2 сформировались в последней трети суббореального - начале субатлантического периода.
Основные отличия спорово-пыльцевых спектров третьей палинозоны (ПЗ 3, глубина 2010 см), выделенной в слое торфа, заключаются в следующем: невысокое, но стабильное содержание пыльцы березы (15-17 %), значительная роль пыльцы верескоцветных кустарничков (до 30 %) и спор сфагновых мхов (40 %). По-прежнему представительно участие пыльцы сосны (30-40 %), появляется непрерывная кривая Pteri-dium aquilinum. В спорово-пыльцевых спектрах уже не встречается пыльца широколиственных пород. На основании вышеперечисленных данных можно сделать вывод, что в период накопления слоя торфа в районе исследования главенствующее положение занимали сосновые, сосново-березовые леса с участием орляка обыкновенного, предпочитающего, как правило, светлые смешанные и сосновые леса, иногда гари. Участие ели в составе древесной растительности незначительно, что, вероятно, обусловлено активной заболачиваемостью юго-западного Бе-ломорья [5]. На месте водоема образовалось болото, на котором доминировали сфагновые мхи и ерикоидные кустарнички. Переход водоема в тельматическую стадию развития делал территорию вокруг него неудобной для расселения человека, стоянки перемещались на более молодые террасы, ближе к урезу воды.
ЗАКЛЮЧЕНИЕ
Поверхность Прибеломорской равнины окончательно сформировалась в процессе неоднократных трансгрессивно-регрессивных стадий Белого моря в поздне- и послеледниковое время.
Колебания уровня моря наряду с изменением климата и растительности имели определяющее значение в размещении стоянок человека и создании памятников наскальных искусств.
Совокупность методов исследования с наибольшей долей достоверности позволяет сделать вывод о том, что около 3820 (С14) лет назад уровень воды в низовье р. Выг снизился и терраса на абсолютной отметке 16,5 м в это время не была покрыта водой. По мнению Э. И. Девятовой, начало регрессии и выход суббореальной террасы из зоны затопления произошли около 3600 (С14) лет назад, подтверждением чему служит возобновление старицы в районе пос. Золотец около 3500 (С14) лет назад [3]. Наши исследования показывают, что это произошло несколько ранее – около 3800 (С14) лет назад. Применение комплекса методов при изучении торфяной залежи позволило уточнить время этого события. Благоприятный климат и разнообразная растительность обеспечивали приток населения, и именно в это время могли появиться археологические памятники, расположенные на указанных абсолютных отметках.
Систематический список пресноводных диатомовых водорослей разреза Залавруга-6
Таблица 1
|
№ п/п |
Наименование вида |
s сЗ s ю о о |
W S сЗ &
О О S и |
н о ю § |
& О |
Глубина, см |
||||||
|
О\ 7 |
7 |
Г'! 7 |
ЧО 7 |
04 7 40 |
7 04 |
7 |
||||||
|
1 |
Achnanthes oestrupii Cl.) Hust. |
o |
a |
ind |
ind |
1 |
||||||
|
2 |
Achnanthes lanceolata var. rostrata (Östrup) Hust. |
o |
k |
ind |
ind |
1 |
||||||
|
3 |
Achnanthes calcar Cl. |
о |
a |
ind |
alkf |
1 |
||||||
|
4 |
Achnanthes kriophila Petersen |
о |
a |
hb |
ind |
1 |
||||||
|
5 |
Achnanthes minutissima Kütz. |
o |
a |
ind |
ind |
1 |
||||||
|
6 |
Amphora ovalis Kütz. |
b |
k |
ind |
alk |
|||||||
|
7 |
Asterionella formosa Hass. |
p |
k |
ind |
alkf |
2 |
||||||
|
8 |
Aulacoseira distans var. distans (Ehrenberg) Sim. |
p |
a |
ind |
acf |
1 |
65 |
24 |
14 |
24 |
12 |
|
|
9 |
Aulacoseira granulata (Ehrenberg) Ralfs |
p |
k |
ind |
alkf |
1 |
9 |
13 |
13 |
7 |
||
|
10 |
Aulacoseira granulata var. angustissima (O. Müll) Hust. |
p |
k |
ind |
alkf |
1 |
||||||
|
11 |
Aulacoseira islandica (O. Müll) |
p |
a |
ind |
ind |
9 |
48 |
13 |
10 |
|||
|
12 |
Aulacoseira islandica ssp. helvetica (O. Müll) |
p |
a |
ind |
ind |
30 |
49 |
62 |
52 |
32 |
||
|
13 |
Aulacoseira italica (Ehrenberg) Sim. |
p |
k |
ind |
alkf |
38 |
17 |
12 |
13 |
14 |
||
|
14 |
Aulacoseira sp. |
p |
1 |
6 |
||||||||
|
15 |
Cocconeis placentula (Ehrenberg) |
o |
b |
ind |
alkf |
1 |
1 |
1 |
||||
|
16 |
Cyclotella comta (Ehrenberg) Kütz. |
p |
k |
ind |
alkf |
1 |
5 |
5 |
2 |
|||
|
17 |
Cyclotella stelligera Cl. et Grün. |
p |
k |
ind |
ind |
1 |
||||||
|
18 |
Cyclotella kuetzingiana Thwait. |
p |
b |
hl |
ind |
4 |
||||||
|
19 |
Cyclotella kuetzingiana var. radiosa Fricke |
p |
b |
hl |
ind |
1 |
1 |
|||||
|
20 |
Cyclotella kuetzingiana var. schumanii Grün. |
p |
b |
hl |
ind |
1 |
||||||
|
21 |
Cymbella cistula (Hemp.) Grün. |
о |
b |
ind |
alkf |
1 |
||||||
|
22 |
Diploneis elliptica Kütz. Cl. |
b |
k |
ind |
alkf |
3 |
||||||
|
23 |
Epithemia zebra var. saxonica (Kütz.) (Ehrenberg) |
о |
k |
ind |
alkf |
4 |
2 |
2 |
3 |
|||
|
24 |
Eunotia clevei Grün. |
о |
a |
ind |
acf |
1 |
||||||
|
25 |
Eunotia diodon (Ehrenberg) |
o |
a |
hb |
acf |
1 |
||||||
|
№ п/п |
Наименование вида |
s s VO о о
|
s c3
& о О s to |
H о VO § |
& к 6 |
Глубина, см |
||||||
|
O\ 7 |
7 |
40 7 |
04 7 40 |
7 04 |
7 |
|||||||
|
26 |
Eunotia fallax Cl. |
о |
k |
hb |
acf |
107 |
53 |
3 |
2 |
|||
|
27 |
Eunotia lunaris (Kütz.) Grün. |
o |
k |
hb |
acf |
1 |
7 |
2 |
2 |
1 |
||
|
28 |
Eunotia microcephala Krasske |
о |
b |
hb |
acf |
2 |
||||||
|
29 |
Eunotia monodon var. maior (W. Sm.) |
о |
k |
hb |
acf |
1 |
||||||
|
30 |
Eunotia pectinalis (Dillw.) Grün. |
o |
k |
hb |
acf |
2 |
2 |
2 |
||||
|
31 |
Eunotia pectinalis var. bidens |
o |
k |
hb |
acf |
2 |
||||||
|
32 |
Eunotia pectinalis var. minor (Kütz.) Grün. |
o |
k |
hb |
acf |
2 |
1 |
|||||
|
33 |
Eunotia praerupta (Ehrenberg) |
o |
b |
hb |
acf |
1 |
1 |
|||||
|
34 |
Eunotia praerupta var. bidens (W. Sm.) Grün. |
o |
a |
hb |
acf |
1 |
||||||
|
35 |
Eunotia sp. |
o |
1 |
1 |
||||||||
|
36 |
Eunotia tenella (Grün.) Hust. |
o |
a |
hb |
acf |
85 |
15 |
2 |
1 |
1 |
||
|
37 |
Eunotia veneris (Kütz.) O. Müll |
o |
b |
ind |
acf |
1 |
3 |
3 |
||||
|
38 |
Fragilaria brevistriata Grün. |
o |
k |
ind |
alkf |
1 |
1 |
43 |
21 |
15 |
38 |
|
|
39 |
Fragilaria construens (Ehrenberg) Grün. |
o |
k |
ind |
alkf |
1 |
1 |
|||||
|
40 |
Fragilaria construens var. binodis (Ehrenberg) Grün. |
o |
k |
ind |
alkf |
1 |
||||||
|
41 |
Fragilaria construens var. venter (Ehrenberg) Grün. |
o |
k |
ind |
alkf |
4 |
3 |
3 |
2 |
7 |
||
|
42 |
Fragilaria lapponica Grün. |
o |
a |
ind |
ind |
1 |
2 |
|||||
|
43 |
Fragilaria leptostauron var. dubia Grün. |
o |
b |
hb |
alkf |
3 |
1 |
|||||
|
44 |
Fragilaria pinnata Ehrenberg |
o |
b |
hl |
alkf |
15 |
13 |
4 |
3 |
|||
|
45 |
Fragilaria pinnata var. lancetula (Schum.) Hust. |
o |
b |
hl |
alkf |
4 |
5 |
|||||
|
46 |
Fragilaria virescense Ralfs |
o |
a |
ind |
ind |
3 |
2 |
1 |
9 |
20 |
||
|
47 |
Fragilaria sp. |
о |
2 |
|||||||||
|
48 |
Frustulia sp. |
b |
acf |
1 |
||||||||
|
49 |
Gomphonema acuminatum var. coronatum (Ehrenberg) W. Sm. |
o |
b |
ind |
ind |
1 |
||||||
|
50 |
Gomphonema bohemicum Reich. |
o |
b |
ind |
ind |
1 |
1 |
1 |
||||
|
51 |
Gomphonema lanceolatum Ehrenberg |
o |
b |
ind |
alkf |
1 |
||||||
|
52 |
Gomphonema parvulum (Kütz.) Grün. |
o |
b |
ind |
ind |
2 |
||||||
|
53 |
Hantzschia amphioxis (Ehrenberg) Grün. |
b |
k |
ind |
alkf |
2 |
1 |
|||||
|
54 |
Navicula subtilissima Cl. |
b |
k |
hb |
acf |
1 |
||||||
|
55 |
Navicula lanceolata (Ag.) Kütz. |
b |
k |
ind |
alkb |
3 |
9 |
|||||
|
56 |
Navicula reinhardii (Grün.) Cl. |
b |
b |
ind |
alkb |
1 |
||||||
|
57 |
Navicula pupula var. rectangularis (Greg.) Grün. |
b |
k |
hl |
ind |
2 |
2 |
|||||
|
58 |
Navicula radiosa Kütz. |
b |
b |
hl |
alkf |
1 |
||||||
|
59 |
Neidium affine var. longiceps (Greg.) Cl. |
b |
a |
ind |
acf |
1 |
||||||
|
60 |
Nitzschia sp. |
b |
||||||||||
|
61 |
Opephora martyi Herib. |
o |
b |
ind |
alkf |
1 |
||||||
|
62 |
Pinnularia borealis Ehrenberg |
b |
a |
ind |
ind |
1 |
2 |
1 |
2 |
1 |
||
|
63 |
Pinnularia braunii var. amphicephala (A. Mayer) Kütz. |
b |
b |
hb |
ind |
1 |
||||||
|
64 |
Pinnularia distinquenda Cl. |
b |
k |
hl |
alkf |
2 |
||||||
|
65 |
Pinnularia globiceps var. krokei Grün. |
b |
k |
ind |
1 |
|||||||
|
66 |
Pinnularia intermedia Lagerst. |
b |
a |
alkf |
1 |
|||||||
|
67 |
Pinnularia interrupta W. Sm. |
b |
k |
ind |
acf |
|||||||
|
68 |
Pinnularia interrupta var. minutissima Hust. |
b |
k |
ind |
39 |
6 |
||||||
|
69 |
Pinnularia islandica Östrup |
b |
a |
2 |
4 |
1 |
7 |
8 |
||||
|
70 |
Pinnularia leptosomoides Cl.-Euler |
b |
k |
hb |
ind |
|||||||
|
71 |
Pinnularia microstauron (Ehrenberg) Cl. |
b |
k |
ind |
ind |
1 |
||||||
|
72 |
Pinnularia microstauron var. brebissonii (Kütz.) Hust. |
b |
b |
ind |
1 |
|||||||
|
73 |
Pinnularia nodosa (Ehrenberg) |
b |
a |
hb |
i |
1 |
||||||
|
74 |
Pinnularia sp. |
b |
3 |
1 |
2 |
1 |
||||||
|
75 |
Pinnularia subcapitata Greg. |
b |
b |
hb |
ind-acf |
1 |
||||||
|
76 |
Pinnularia viridis (Nitzsch.) Ehrenberg |
b |
b |
hb |
acf |
2 |
||||||
|
№ п/п |
Наименование вида |
s § s VO о о
|
s c3
& о О s to |
H о VO § |
к 6 |
Глубина, см |
||||||
|
04 7 |
7 |
40 7 |
04 7 40 |
7 04 |
7 |
|||||||
|
77 |
Pinnularia viridis var. intermedia Cl. |
b |
k |
ind |
ind |
1 |
||||||
|
78 |
Stephanodiscus astraea (Ehrenberg) Grün. |
p |
b |
ind |
ind |
2 |
2 |
2 |
5 |
5 |
||
|
79 |
Stephanodiscus astraea var.intermedia Fricke |
p |
b |
ind |
ind |
1 |
2 |
|||||
|
80 |
Stephanodiscus sp. |
p |
3 |
|||||||||
|
81 |
Synedra ulna (Nitzsch.) Ehrenberg |
o |
k |
ind |
alkf |
1 |
||||||
|
82 |
Synedra nana Meist. |
o |
a |
1 |
1 |
1 |
||||||
|
83 |
Synedra sp. |
o |
1 |
|||||||||
|
84 |
Tabellaria flocculosa (Roth.) Kütz. |
o |
b |
hb |
acf |
2 |
3 |
28 |
6 |
8 |
||
|
85 |
Tabellaria fenestrata (Lyngb.) Kütz. |
p |
b |
hb |
acf |
2 |
1 |
1 |
6 |
8 |
||
|
Epidathia muellery desmidiaceae |
1 |
|||||||||||
|
неопределенные |
1 |
1 |
||||||||||
|
Итого |
250 |
123 |
250 |
250 |
251 |
250 |
255 |
|||||
Примечание. Местообитание: p – планктон; b – бентосные; o – обрастатели; биогеография: a – арктобореальные, b – бореальные, k – космополиты; галобность: hl – галофилы, hb – галофобы, ind – индифференты; отношение к рН: alkf – алкалифилы, acf – ацидофилы, ind – индифференты.
Таблица 2
|
Список морских диатомовых во |
дор |
ослей разре |
за |
Залавруга-6 |
||
|
№ |
Название вида |
м |
б г рН |
Глубина, см |
||
|
п/п |
------ |
|||||
|
04 |
W*) (N 40 04 1/4 о еч m m m |
|||||
|
04 i/4 (N 40 04 i/4 |
||||||
|
1 |
Achnanthes brevipes Ag. |
pb |
k mz |
3 1 |
||
|
2 |
Achnanthes hauskiana Grun. |
b |
k mz |
1 |
||
|
3 |
Cocconeis scutellum Ehr. |
b |
k mz |
1 3 2 |
||
|
4 |
Diploneis interrupta (Kutz.) Cl. |
b |
mz alkf |
1 |
||
|
5 |
Diploneis smithi (Breb.) Cl. |
b |
k mz alkf |
1 1 |
||
|
6 |
Grammatophora oceanica (Ehr.) Grun. |
b |
k pg |
1 1 |
||
|
7 |
Grammatophora marina (Lyngb.) Kutz. |
b |
k pg |
2 |
||
|
8 |
Hyalodiscus scoticus (Kutz.) Grun. |
tp |
b mz |
1 1 |
||
|
9 |
Navicula humerosa Breb. |
b |
k mz |
4 1 |
||
|
10 |
Navicula hungarica Grun. |
b |
1 |
|||
|
11 |
Navicula mutica Kutz. |
b |
k hl alkf |
1 |
2 |
|
|
12 |
Navicula mutica var.cohii (Hilse) Grun. |
b |
k hl alkf |
2 |
||
|
13 |
Navicula pusilla W. Smith |
b |
bor hl |
3 3 |
||
|
14 |
Nitzschia obtusa (W. Smith.) |
b |
1 13 10 8 |
|||
|
15 |
Nitzschia punctata (W. Smith) Grun. |
b |
bor mz |
1 |
||
|
16 |
Nitzschia spp. |
b |
21 2 |
|||
|
17 |
Nitzschia intermedia Hantz. |
b |
1 |
|||
|
18 |
Odontella aurita (Lyngb.) Agardh. |
тп |
k pg |
2 |
||
|
19 |
Paralia sulcata. (Ehr.) Cl |
p |
mz |
1 46 |
||
|
20 |
Paralia sulcata var. crenulata Grun. |
p |
2 22 35 37 22 |
|||
|
21 |
Rhabdonema arcuatum (Lyngb.) Kutz. |
b |
k mz |
5 |
||
|
22 |
Rhabdonema minutum Kutz. |
b |
k mz |
11 |
||
|
23 |
Rhoicosphaenia curvata (Kutz.) Grun. |
1 |
||||
|
24 |
неопределенные |
1 |
||||
Примечание. М – местообитание: pb – полубентосные; b – бентосные (донные и эпифиты); tp – тихопелагические; б – биогеография: k – космополиты; bor – бореальные; г – галобность: pg – полигалобы; mz – мезогалобы; hl – галофилы; отношение к рН: alkf – алкалифилы.
Список литературы Новые данные о миграции береговой линии Белого моря
- Горецкий Г. И. Неолитические стоянки в районе г. Кеми//Краткие сообщения о докладах и полевых исследованиях Института истории материальной культуры. Вып. IX. М., 1941.
- Давыдова Н. Н. Диатомовые водоросли -индикаторы экологических условий водоемов в голоцене. Л.: Наука, 1985. 244 с.
- Девятова Э. И. Геология и палинология голоцена и хронология памятников первобытной эпохи в Юго-Западном Беломорье. Л.: Наука. Ленингр. отд-ние, 1976. 121 с.
- Диатомовый анализ. Определитель ископаемых и современных диатомовых водорослей. Л.: Госгеолиздат, 1949. Кн. 1, 2.
- Елина Г. А. Корреляция спорово-пыльцевых спектров голоцена Карельской АССР, Ленинградской области и Финляндии//Палинология голоцена. М.: Наука, 1971. С. 91-104.
- Елина Г. А., Лукашов А. Д., Юрковская Т. К. Позднеледниковье и голоцен восточной Фенноскандии. Петрозаводск: Карельский научный центр РАН, 2000. 242 с.
- Заславская Н. В. Флора и растительность засоленных приморских экотопов западного побережья Белого моря: Автореф. дисс. … канд. биол. наук. Петрозаводск, 2007. 24 с.
- Колька В. В., Евзеров В. Я., Мёллер Я.Й., Корнер Д.Д. Послеледниковые гляциоизоста-тические поднятия на северо-востоке Балтийского щита//Новые данные по геологии и полезным ископаемым Кольского полуострова/Ред. Ф. П. Митрофанов. Апатиты: Кольский научный центр РАН, 2005. С. 15-25.
- Колька В. В., Корсакова О. П. Относительное перемещение береговой линии Белого моря и возраст геологических объектов -каменных лабиринтов Кольского региона//Геология морей и океанов: Материалы XVIII Междунар. науч. конф. по морской геологии. Т. III. М.: ГЕОС, 2009. С. 179-184.
- Куприянова Л. А., Алешина Л. А. Пыльца и споры растений флоры Европейской части СССР. Т. 1. Л.: Наука, 1972. 171 с.
- Куприянова Л. А., Алешина Л.А. Пыльца двудольных растений флоры Европейской части СССР. Lamiaceae-Zygophyllaceae. Л.: Наука, 1978. 184 с.
- Лаврова М. А. История Белого моря//Последний европейский ледниковый покров. К VII Конгрессу INQUA (США, 1965). М.: Наука, 1965. С. 267-276.
- Лаврова Н.Б., Колька В.В., Корсакова О.П. Пыльца как индикатор миграции береговой линии Белого моря//Эволюция органического мира и биотические кризисы: Материалы LVI сессии Палеонтологического общества при РАН (5-9 апреля 2010 г.). СПб., 2010. С. 64-66.
- Лийвранд Э. Методические проблемы палиностратиграфии плейстоцена. Таллинн: Валгус, 1990. 176 с.
- Лобанова Н. В. Петроглифы старой Залавруги: новые данные -новый взгляд//Археология, этнография и антропология Евразии. 2007. № 1. С. 127-135.
- Невесский Е. Н., Медведев В. С., Калиненко В. В. Белое море. Седиментогенез и история развития в голоцене. М.: Наука, 1977. 236 с.
- Пыльцевой анализ/Под ред. И. М. Покровской. М.: Гостоптехиздат, 1950. 571 с.
- Раменская М. Л. Анализ флоры Мурманской области и Карелии. Л.: Наука, 1983. 216 с.
- Система Белого моря. Т. 1. Природная среда водосбора Белого моря/Отв. ред. А. П. Лисицын. М.: Научный мир, 2010. 480 с.
- Тарасов А. Ю., Мурашкин А. И. Новые материалы с поселения Залавруга I и проблема датировки петроглифов Новой Залавруги//Археологические вести. 2002. № 9. С. 41-44.
- Шелехова Т. С. История развития малых озер Прибеломорской низменности в голоцене по данным диатомово-го анализа//Материалы 4-й междунар. науч. конф. «Теоретические и прикладные аспекты современной лимнологии». Минск, 2009. С. 55-58.
- Шелехова Т. С., Колька В.В., Корсакова О. П. Диатомовые комплексы голоцена в донных осадках озер карельского побережья Белого моря//Материалы XI междунар. науч. конф. диатомологов стран СНГ «Диатомовые водоросли как биоиндикаторы современного состояния окружающей среды и их роль в палеоэкологии и био-стратиграфии». Минск, 2009. С. 142-144.
- Шелехова Т. С., Колька В. В., Корсакова О. П. Особенности диатомовых комплексов в донных осадках озер карельского побережья Белого моря//Эволюция органического мира и биотические кризисы. LVI сессия палеонтологического общества. СПб., 2010. С. 117-119.
- Шелехова Т. С., Лаврова Н. Б. Палеоэкологические условия развития юго-западного Беломорья в голоцене//Геология и полезные ископаемые Карелии. Вып. 11. Петрозаводск: Карельский научный центр РАН, 2008. С. 254-265.
- Grimm E. S. TILIA and TILIA GRAPH: Pollen spreadsheet and graphics programm//8th International Palynological Congress. Programm and Abstracts. Aix-en-Provence, France, 1992. 56 p.
- Krammer K., Lange-Bertalot H. Bacillariophiceae//Su?wasserflora von mitteleuropa/Ed. by H. Ettl, J. Gerloff, H. Heying, D. Mollenhauer. Band 2/1-4. Stuttgart: G. Fisher Vertag, 1986-1991.
- Renberg I., Hellberg T. The pH history of Lakes in Southwestern Sweden, as calculated from subfossil diatom flora of the sediments//Ambio. Vol. 11. № 1. 1982. P. 30-33.
- Tynni R. Uber Finnlands rezente und subfossile Diatomeen VIII-X1. Geol. Surv. Finland Bull. 274: 1-55 (1975); 284: 1-37 (1976); 296: 1-55 (1978); 312: 1-93 (1980).
